
Circulating exosomal lncRNA contributes to the pathogenesis of spinal cord injury in rats
2022-09-16 04:19JianAnLiMingPengShiLinCongMingYuGuYiHengChenSiYiWangZhenHuaLiChunFangZanWanFuWei
中国神经再生研究(英文版) 2023年4期
Jian-An Li , Ming-Peng Shi , Lin Cong, Ming-Yu Gu Yi-Heng Chen Si-Yi Wang Zhen-Hua Li, Chun-Fang Zan , Wan-Fu Wei
Abstract Exosome-derived long non-coding RNAs (lncRNAs) are extensively engaged in recovery and repair of the injured spinal cord, through different mechanisms.However, to date no study has systematically evaluated the differentially expressed lncRNAs involved in the development of spinal cord injury.Thus, the aim of this study was to identify key circulating exosome-derived lncRNAs in a rat model of spinal cord injury and investigate their potential actions.To this end, we established a rat model of spinal cord hemisection.Circulating exosomes were extracted from blood samples from spinal cord injury and control(sham) rats and further identified through Western blotting and electron microscopy.RNA was isolated from the exosomes and sequenced.The enrichment analysis demonstrated that there were distinctively different lncRNA and mRNA expression patterns between the two groups.Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis and Gene Ontology (GO) functional analysis were performed to determine the possible involvements of upregulated and downregulated lncRNAs in various pathways and different biological processes, as well as their cellular locations and molecular functions.Furthermore,quantitative reverse transcription-polymerase chain reaction showed that the expression of five lncRNAs--ENSRN0T00000067908, XR_590093, XR_591455,XR_360081, and XR_346933--was increased, whereas the expression of XR_351404, XR_591426, XR_353833, XR_590076, and XR_590719 was decreased.Of note, these 10 lncRNAs were at the center of the lncRNA-miRNA-mRNA coexpression network, which also included 198 mRNAs and 41 miRNAs.Taken together,our findings show that several circulating exosomal lncRNAs are differentially expressed after spinal cord injury, suggesting that they may be involved in spinal cord injury pathology and pathogenesis.These lncRNAs could potentially serve as targets for the clinical diagnosis and treatment of spinal cord injury.
Key Words: exosome; inflammation; lncRNA; microenvironment; miRNA; mRNA; spinal cord injury; spinal cord repair
Introduction
Spinal cord injury (SCI) is catastrophic, usually leading to permanent disabilities and imposing a heavy social burden (de Araújo et al., 2022;Fauss et al., 2022).Current epidemiological data show that the incidence of SCI worldwide is about 12.1-57.8 cases per million people every year (Van den Berg et al., 2010; Jazayeri et al., 2015).In terms of pathogenesis, the primary cause of SCI is trauma, such as sudden falls, sport injuries, and traffic accidents.The secondary causes are progressive edema, ongoing ischemia,and infiltrating inflammatory cells, as well as the release of proinflammatory cytokines, which further aggravate the initial injury (Rouanet et al., 2017;Fadeev et al., 2021).With regard to clinical treatment, current measures are still limited to early decompression, neuroprotection, neuroregeneration,airway and blood pressure management, and rehabilitative training, as there is a lack of knowledge regarding specific therapeutic targets (Rouanet et al., 2017).Even though many advancements have been made in the field of bioengineering, SCI treatment and prognosis remain unsatisfactory because of our limited understanding of SCI pathogenesis (O’Shea et al., 2017).
Exosomes are membranous-bilayer nanovesicles that participate in numerous biological activities (Kalluri and LeBleu, 2020; Kim et al., 2021).Recently, O’Brien et al.(2020) showed that exosomes may serve as cellular communicators, disease biomarkers, therapeutic targets, “rubbish bags,”and drug carriers.In addition, recent studies have suggested that exosomes are pivotal in spinal cord repair in the context of SCI (Li et al., 2019; Yu et al.,2019; Zhong et al., 2020), and that their actions are mediated, at least in part, through the Wnt/β-catenin signaling pathway.Yu et al.(2019) showed that exosomes derived from miRNA-29b-modified bone marrow-derived stem/stromal cells (BMSCs) could enhance rat limb function after SCI.Noncoding RNAs (ncRNAs) have a critical effect on the development of SCI, as they are extensively involved in several processes, such as neuronal apoptosis and demyelination, as well as inflammatory reactions (Wang et al., 2019a;Ling et al., 2021).Of special note, exosomal RNA has substantially different properties than cellular RNA and can reflect the actual physiological and/or pathological status of their source cells (O’Brien et al., 2020).In addition,SCI affects non-cell-autonomous mechanisms as well as interactions among neurons, microglia, astrocytes, and other immune cells, a process that may or may not be accompanied by the secretion of growth-modulating molecules and that varies at different lesion sites (O’Shea et al., 2017).Long non-coding RNAs (lncRNAs) are also associated with several pathophysiological processes following SCI, such as neural apoptosis and inflammation, as well as oxidative stress (Wang et al., 2019a).In addition, Ji and coworkers demonstrated that the lack of miR-21, which is an exosome-shuttled miRNA, may decrease the beneficial impact that exosomes have on the injured spinal cord (Ji et al.,2019).
Given the fact that they are highly involved in human diseases, such as sepsis, cancer, and atrial fibrillation (Yao et al., 2020; Li et al., 2021; Kang et al., 2022), circulating exosomal lncRNAs can be considered diagnostic markers and/or therapeutic targets.Yao et al.(2020) found that circulating exosomal lnc-EPC1-4 and lnc-GPR89B-15 levels are correlated with serum α-fetoprotein expression, suggesting that both of these lncRNAs contribute to hepatocarcinogenesis.Thus, based on our current knowledge of exosomes and lncRNAs in SCI, it is reasonable to speculate that exosomal lncRNA-elicited changes in the microenvironment may contribute to SCI development.Therefore, the aims of this study were to obtain a comprehensive overview of all exosomal lncRNAs involved in SCI rats, and especially to identify significantly upregulated or downregulated lncRNAs that could be promising therapeutic targets for SCI in the future.
Methods
Ethics statement
This study was approved by and conducted in strict accordance with the guidelines of the Animal Care and Use Committee of Tijian Hospital of China.Ethical approval was granted on December 19, 2018.All experiments were designed and reported according to the Animal Research: Reporting ofIn Vivo
Experiments (ARRIVE) guidelines (Percie du Sert et al., 2020).Experimental rats
Six adult female Sprague-Dawley (SD) rats (7-8 weeks old, 230-250 g)from the Orthopaedic Research Institute of Tianijn Hospital were randomly assigned to one of two groups (n
= 3 for each group) using the random number table method.In this field, it is very normal to use only female or only male rats (Onifer et al., 2007).All rats were kept in the same environment with the same food and at a temperature of 24°C with a standard 12/12-hour light/dark cycle.The rats were housed two per cage under these conditions for at least 1 week prior to surgery.Then, SCI surgery was performed on the rats in the experimental group, while only the skin was injured on rats in the control group.After surgery, the rats were housed singly for 24 hours while they recovered.The experimental design is shown in Figure 1.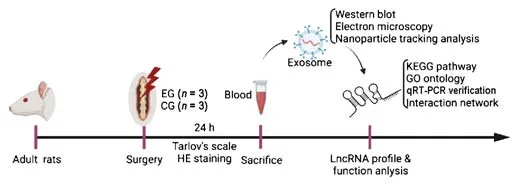
Figure 1|Experimental design.
Spinal cord injury surgical procedure
The experimental rat SCI model was established as described previously (Ding et al., 2016).Briefly, the rats were deeply anesthetized using 3.5% isoflurane(Vetpharma, Barcelona, Spain) for induction and 1.75% isoflurane for maintenance at a rate of 300-600 mL/min before surgery, then immobilized.Next, the skin on the back was cut open from T8 to T12.After the spinal cord was fully exposed, the scalpel was carefully inserted into the dorsal midline of the spinal cord at a vertical angle, and the left half of the spinal cord was cut at the level of T10.In contrast, only the skin incision from T8 to T12 was made on rats in the control (sham) group.After surgery, the incisions were sutured for both groups of rats.In addition, all rats were given the same postoperative care.A heating pad was prepared to keep the temperature of rats around 37°C, and the surgical incisions were sterilized with 2% iodophor disinfectant.The day after SCI surgery, all rats were injected with penicillin solution at a dose of 5 mL/kg to prevent postoperative infections.At 24 hours post-surgery, blood was drawn, and the rats were subjected to transcardial perfusion with 0.9% saline and then sacrificed (Beyotime, Shanghai, China).
Evaluation of the rat model of spinal cord injury
Tarlov’s scale and hematoxylin and eosin (HE) staining were used to determine whether the rat model of SCI was successfully established.Firstly, rat motor function scores were evaluated using a modified Tarlov’s scale, the specific details of which were described by Yu and Qian (2020).Two investigators who did not perform the surgery evaluated the motor function of rat hind limbs using the modified Tarlov’s scale.At 24 hours after surgery, hind limb motor function was evaluated using the following scale: 0: no hind limb movement;1: hind limb motion without gravity, barely perceptible; 2: hind limb motion against gravity but unable to support weight; 3: able to stand, crawl with assistance; 4: able to walk with assistance; 5: normal function.For HE staining,the whole spinal cord was harvested from both the control and experimental groups after transcardial perfusion.A 0.8-cm spinal segment of from T8 to T12 was collected and fixed in 4% paraformaldehyde (Morphisto GmbH,Frankfurt, Germany) for 24 hours, dehydrated with different concentrations of ethanol (75%, 90%, 100%), immersed and embedded in paraffin, and then sliced into serial 4-μm sections.For staining, the sections were deparaffinized with xylene twice for 5 minutes, then dehydrated in an ethanol gradient(75%, 90%, 100%).Following this, the spinal cord sections were stained with hematoxylin (MilliporeSigma, Burlington, MA, USA) for 10 minutes and eosin(MilliporeSigma) for 30 seconds.Finally, representative images were acquired by light microscope (Olympus BH-2 Microscope, Olympus, Tokyo, Japan).
Exosome extraction
At 24 hours after SCI surgery, 1- to 2-mL blood samples containing exosomes were carefully drawn from the abdominal aorta of the six rats included in this study.The samples from the CG were numbered CG1, CG2, and CG3, and the samples from the EG were numbered EG1, EG2, and EG3.To isolate the plasma, the blood specimens were centrifuged at 300 ×g
for 10 minutes.Afterward, exosomes were separated and enriched based on density gradient centrifugation.Briefly, an exoEasy column (Thermo Fisher Scientific, Waltham,MA, USA) was filled with the mixture of plasma and XBP buffer, then centrifuged at 500 ×g
for 1 minute.Next, 10 mL of XWP buffer was added to wash the column by centrifugation at 5000 ×g
for 5 minutes.Then, the membrane was incubated in 0.4-1 mL of XE buffer for 60 seconds, and the eluent was centrifuged for 5 minutes at 5000 ×g
.The final eluent contained the purified exosomes.Exosomes were extracted from all six samples following the same procedure.Identification of exosomes by western blotting
To further confirm the features of exosomes collected as described above,the markers CD9 and HSP70 were detected by Western blot, as previously described (Park and Yang, 2018).Briefly, the vesicles were denatured in radioimmunoprecipitation assay (RIPA) buffer (MilliporeSigma).A bicinchoninic acid (BSA) protein assay kit (MilliporeSigma) was used to determine protein concentrations of these samples, which were further diluted in 2× lithium dodecyl sulfate (LDS) lysis buffer (Thermo Fisher Scientific, Waltham, MA, USA).After mixing, all prepared samples were boiled at 95°C for 10 minutes.For electrophoresis, 30 μL of sample was loaded into each well, and the 11% Bis-Tris gel (Thermo Fisher Scientific) was run at 120 V for 15 minutes and 140 V for 80 minutes.Then, the separated proteins were transferred to a methanol-activated polyvinylidene fluoride (PVDF) membrane(Merk Millipore, Darmstadt, Germany) at 30 V for 90 minutes.Following this, the PVDF membrane was blocked with 5% bovine serum albumin (BSA)(MilliporeSigma) blocking solution at room temperature for 2 hours, then incubated with mouse CD9 (Cat# sc-13118, RRID: AB_627213), mouse HSP70(Cat# sc-66048, RRID: AB_832518), and mouse β-actin (Cat# sc-47778, RRID:AB_626632) antibodies (Santa Cruz Biotechnology, Dallas, TX, USA) in 5% BSA blocking buffer (1:1000) at 4°C overnight.On the second day, all used primary antibody solution was collected and stored at -20°C for reuse, and the membranes were rinsed with homemade 0.1% Tween-20-Tris-buffered saline(TBS-T).After three washes, the membrane was incubated with horseradish peroxidase (HRP)-labeled secondary antibody diluted 1:10,000 (Jackson Laboratory, California, USA) at room temperature for 1 hour.After three washes with TBS-T, the membrane was incubated with chemiluminescence detection reagents (Thermo Fisher Scientific) and developed using a LI-COR imaging systems (LI-COR Biosciences, NE, USA) to visualize the protein bands.The relative expression of the target protein was normalized to β-actin.
Verification of exosomes by electron microscopy
Electron microscopy (EM) was used to obtain high-resolution images of exosomes.A TEM H-7000 electron microscope (Hitachi Ltd., Hitachi, Japan)was used as described previously (Zhao et al., 2018).Briefly, the isolated exosomes were rinsed with phosphate-buffered saline (PBS), fixed in 2%osmic acid (Carl Roth, Karlsruhe, Germany) at 4°C for 2 hours, rinsed with PBS again, and then dehydrated with different concentrations (75%, 90%,and 100%) of ethanol.Following this, the exosomes were impregnated with acetone (Carl Roth) several times, embedded, polymerized, and then stained with uranyl acetate (Carl Roth) for 10 minutes and lead acetate (Carl Roth) for 10 minutes.After washing, representative images of exosome structure were acquired using a transmission electron microscope.
Nanoparticle tracking analysis
To further determine the size distribution of the exosomes, we performed nanoparticle tracking analysis (NTA) using a Malvern Nanoparticle Size Analyzer (Malvern Panalytical, Malvern, UK).Exosomes were prepared, and particle concentrations within the specified size range (10-1000 nm) were determined using ZetaView software (Particle Metrix, Meerbusch, Germany).Particle sizes within the range of 10-1000 nm were analyzed, and exosome diameter was determined by peak analysis (Oosthuyzen et al., 2013).
Exosomal RNA isolation
To obtain total exosomal RNA, Trizol reagent (Thermo Fisher Scientific) was used following the standard protocol.RNA concentrations were determined using a Nanodrop spectrophotometer (Thermo Fisher Scientific).Before use,the purity and integrity of the extracted RNA were assessed.The OD260/OD280 values ranged from 1.8 to 2.1, indicating sufficient purity, and 1%agarose gel (Thermo Fisher Scientific) analysis demonstrated sufficient integrity.
Construction of RNA library and RNA sequencing
To remove the ribosomal RNAs (rRNAs) from the total RNA, an NEBNext rRNADepletion Kit (New England Biolabs, Ipswich, MA, USA) was used according to the manufacturer’s instructions.Next, an NEBNext® UltraII Directional RNA Library Prep Kit (New England Biolabs) was used to construct RNA libraries according to the standard procedures, and the quality and quantity of the RNA libraries was assessed using a BioAnalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA).Finally, RNA sequencing was conducted on an illumina HiSeq sequencer (Cloud-Seq Biotech, Shanghai, China).
RNA sequencing data analysis
The raw data for the paired-end reads were obtained using an Illumina HiSeq 4000 system (Illumina, San Diego, CA, USA).To identify high-quality reads,the 3′ adaptor was trimmed, and low-quality reads were eliminated based on Cutadapt software (https://cutadapt.readthedocs.io/; Martin, 2011).Then,the pre-trimmed high-quality reads were analyzed to determine the relative expression of lncRNAs as well as mRNAs (Trapnell et al., 2010).To this end,these pre-selected reads were aligned with the human reference genome using HISAT2 software (http://daehwankimlab.github.io/hisat2/; Kim et al.,2015).Next, using gene transfer format (GTF) files from the Ensembl database(http://www.ensembl.org/; Hubbard et al., 2002), the fragments per kilobase of exon model per million mapped fragments (FPKM) values indicating the lncRNA and mRNA expression patterns were calculated using Cuffdiff software.Additionally, specific fold changes and correspondingP
-values were calculated based on the FPKM values to identify differentially expressed lncRNAs and mRNAs.To find functional genes near the lncRNAs, the upstream and downstream regions of the genome were analyzed for predicted genes,as described in more detail later.Moreover, GO analysis (http://www.geneontology.org; Balakrishnan et al., 2013) and KEGG pathway analysis(http://www.genome.jp/kegg; Kanehisa and Sato, 2020) were carried out for upregulated and downregulated lncRNAs, to identify functional processes and pathways that they are potentially involved in.The value -log10(P
-value) was calculated as the enrichment score.For both GO and KEGG pathway analysis,fold change > 2 andP
-value < 0.05 were taken as the threshold for differential expression of lncRNAs and mRNAs.Quantitative reverse transcription-polymerase chain reaction
To confirm these circulating differentially expressed lncRNAs in blood from rats, we used quantitative reverse transcription-polymerase chain reaction(qRT-PCR) to determine the relative expression of the 10 lncRNAs extracted from blood samples from both groups.The relevant predicted pathway information for each lncRNA was checked using the NCBI Nucleotide database(https://www.ncbi.nlm.nih.gov/nucleotide/) (Muñoz-Baena and Poon, 2022).RNA was isolated using Trizol reagent (Thermo Fisher Scientific), as mentioned above.cDNAs were synthesized using a SuperScriptIV First-Strand synthesis kit (Thermo Fisher Scientific).SYBR Green master mix (CloudSeq, Shanghai,China) was used for the RT-PCR reaction.Primers targeting the lncRNAs and a housekeeping gene, i.e.glyceraldehyde-3-phosphate dehydrogenase(GAPDH), were designed using Primer Premier 5.0 software (PREMIER Biosoft,Palo Alto, CA, USA).All primers were purchased from Shanghai CloudSeq Biotechnology Co., Ltd.The primer sequences are shown in Table 1.All raw data were acquired using a QuantStudio 5 Real-Time PCR System (Thermo Fisher Scientific), and the 2method was applied to determine the relative expression of these lncRNAs (Livak and Schmittgen, 2001).
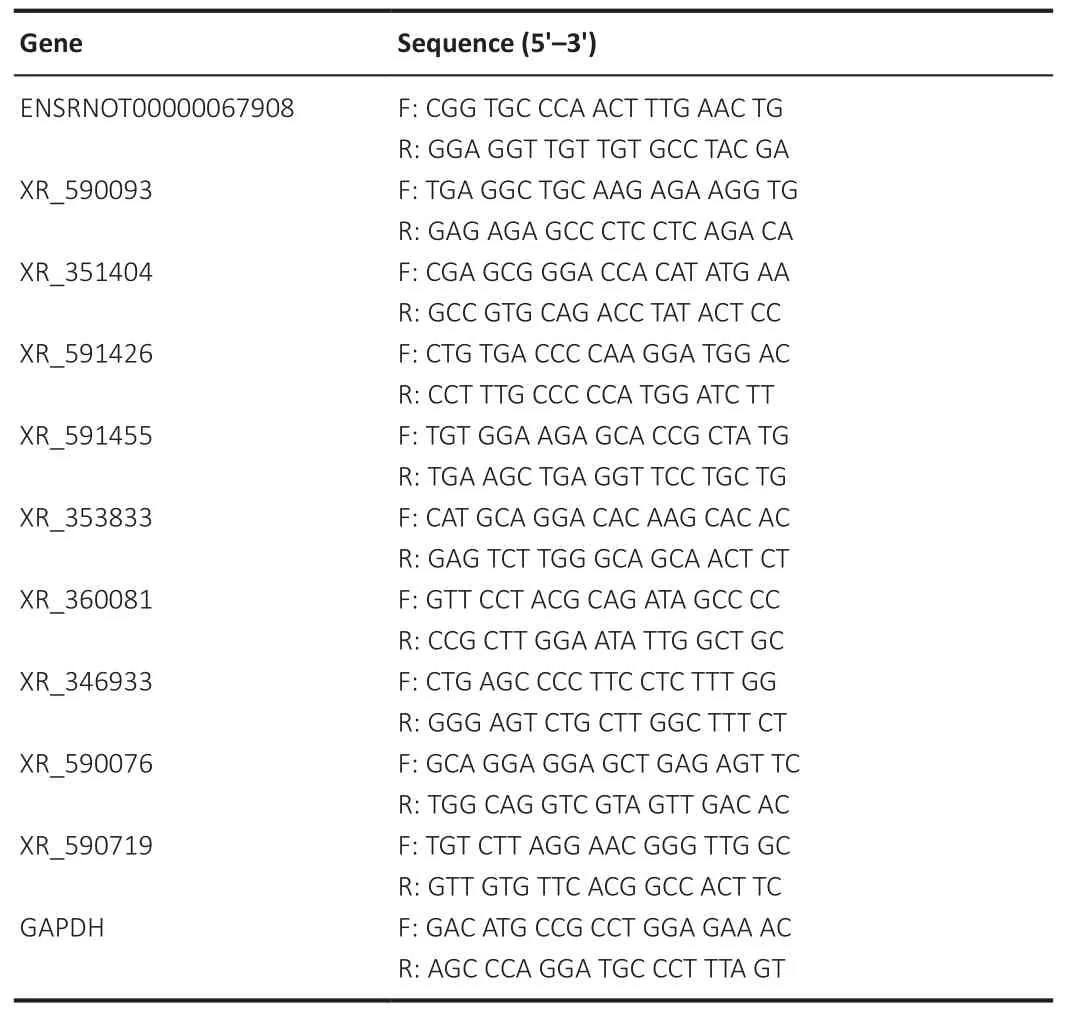
Table 1 |The primer sequences for quantitative reverse transcription-polymerase chain reaction
LncRNA-miRNA-mRNA network construction
Given that functional interactions among lncRNAs, mRNAs, and miRNAs are important in gene regulation, miRNAs corresponding to the differentially expressed lncRNAs were predicted using miRcode (http://www.mircode.org/), and potential mRNAs were predicted using Targetscan (https://www.targetscan.org/vert_80/), miRTarBase (https://mirtarbase.cuhk.edu.cn/), and miRDB (http://www.mirdb.org/; Shan et al., 2021).Finally, the lncRNA-miRNA-mRNA coexpression network was constructed using Cytoscape software(https://cytoscape.org/what_is_ cytoscape.html; Lei et al., 2022), with the differentially expressed lncRNAs placed at the center of the network.
Statistical analysis
No statistical methods were used to predetermine sample sizes; however, our sample sizes are similar to those reported in previous publications (Marcol et al., 2015; Zhou et al., 2020).Of note, functional and histological analyses were conducted by researchers blinded to the group, whereas the sequencing analyses were not performed blinded.GraphPad Prism 8 software (GraphPad Software, San Diego, CA, USA, www.graphpad.com) was employed to analyze the experimental results.Data are shown as means ± standard deviation(SD) using data from more than three statistically independent experiments.To compare the statistical significances among different groups, Student’st
-test for independent samples was used.When theP
-value was < 0.05, the difference was regarded as statistically significant.Results
Successful establishment and evaluation of spinal cord hemisection model
The experimental group rats had significantly lower limb function scores (i.e.Tarlov’s score 1-2) after spinal cord hemisection (P
< 0.001) compared with the control group, as shown in Figure 2A.This observation suggested that we successfully established the SCI rat model.In addition, HE-stained images showed that the spinal cord was injured in rats from the experimental group(Figure 2B), which was consistent with the functional finding described above.
Figure 2|Establishment of a rat model of spinal cord injury and evaluation by functional score and hematoxylin and eosin staining.
Identification of exosomes
To determine the features of the exosomes that were extracted from the plasma of the six experimental rats, Western blot and electron microscopy were used to detect the protein markers and the structure of exosomes,respectively.CD9 and HSP70, as two of the most classical protein markers for exosomes (Kowal et al., 2016), were chosen for detection.The representative Western blot shown in Figure 3A demonstrated that the samples from both groups were positive for CD9 and HSP70 (Figure 3A), indicating that exosomes were extracted successfully.In addition to western blotting, electron microscopy also showed that these exosomes were 62-91 nm in diameter and were circular vesicles with clear and complete membrane structures, and indicated by the arrow in Figure 3B.Measurement by NTA showed that the average exosome diameter was about 145.1 nm.Collectively, these results confirm that the extracted exosomes were suitable for use in the following analyses.
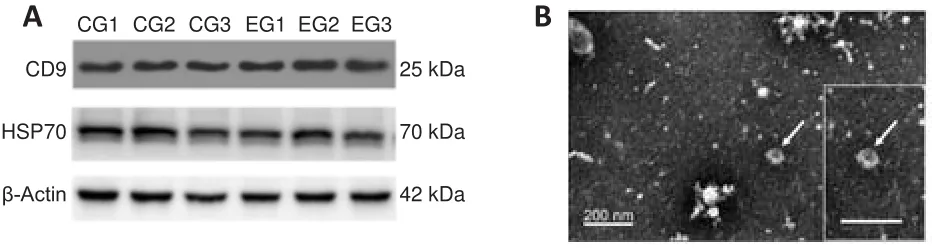
Figure 3 | Extraction and identification of circulating exosomes.
RNA expression profile analysis results
Quality control analysis of the RNA sequencing library showed that the volume of the RNA was greater than 1 μL, while the OD260/OD280 ratios ranged between 1.8 and 2.1, indicating that the samples were suitable for use in the following experiments.Enrichment analysis identified 76 differentially expressed lncRNAs, of which 30 were upregulated and 46were downregulated.Additionally, we detected 372 upregulated and 635 downregulated differentially expressed mRNAs.FPKM values were used to create heat maps of the differentially expressed lncRNAs (Figure 4A) and mRNAs (Figure 4C), which showed that there was a significantly different expression pattern of mRNAs and lncRNAs in rats after SCI in comparison with those in the control rats (P
< 0.05).Similarly, the volcano plots showed consistent changes in differentially expressed lncRNAs (Figure 4B) and mRNAs(Figure 4D) in SCI rats.
Figure 4 | lncRNA and mRNA expression is significantly different after spinal cord injury.
KEGG pathway and GO functional analysis of differentially expressed lncRNAs
Given the fact that lncRNAs are closely associated with their adjacent coding genes (Paraskevopoulou et al., 2013), we performed further enrichment analysis of the exosomal lncRNAs that were differentially expressed in SCI rats to predict their possible functions.To this end, both GO analysis and KEGG analysis were carried out to identify functional activities and signaling pathways involved in SCI.Figures 5 and 6 show the top ten enriched GO terms and KEGG pathways.Analysis of the upregulated lncRNAs is shown in Figure 4,and analysis of the downregulated lncRNAs is shown in Figure 6.The results may provide some meaningful indications for future research.
The top ten enriched biochemical pathways for the upregulated lncRNAs were glycosylphosphatidylinositol (GPI)-anchor biosynthesis, synaptic vesicle cycle, nicotine addiction, type II diabetes mellitus, taste transduction, longterm depression, GABAergic synapse, longevity regulating pathway, morphine addiction, and glycerophospholipid metabolism (Figure 5A).The top ten enriched terms for upregulated lncRNAs in three categories (biological process, cellular component, and molecular function) are shown in Figure 5B-D.
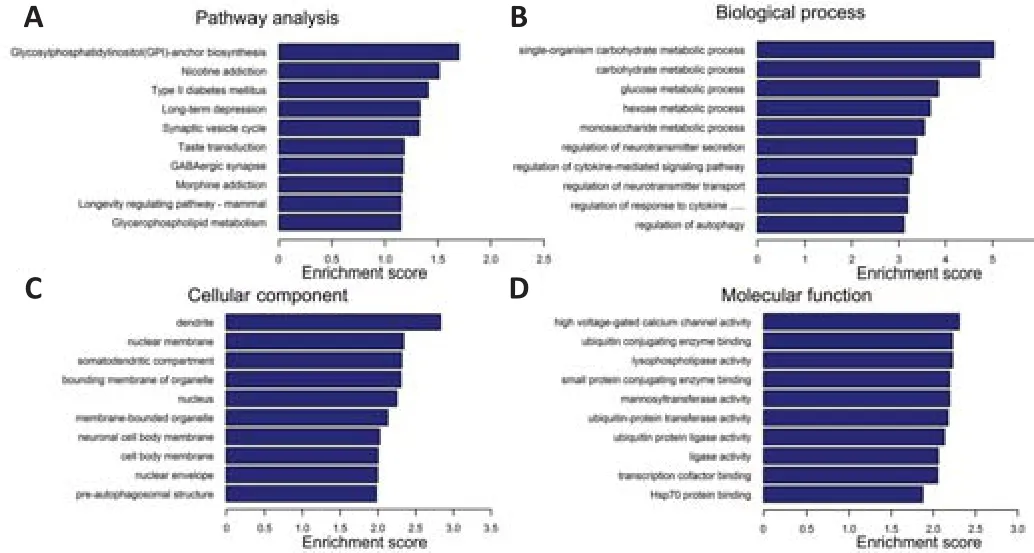
Figure 5| KEGG pathway analysis and GO functional analysis of upregulated circulating lncRNAs after spinal cord injury.

Figure 6| KEGG pathway analysis and GO functional analysis of downregulated circulating lncRNAs after spinal cord injury.
As for downregulated lncRNAs, the top ten enriched biochemical pathways were fatty acid biosynthesis, biotin metabolism, ether lipid metabolism,other types of O-glycan biosynthesis, proteasome, mTOR signaling pathway,adherens junction, fatty acid metabolism, and glycerophospholipid metabolism (Figure 6A).The top ten enriched terms for downregulated lncRNAs in three categories (biological process, cellular component, and molecular function) are shown in Figure 6B-D.
qRT-PCR quantification results
Next, qRT-PCR was performed to confirm the relative expression of the 10 differentially expressed lncRNAs identified as described above.The relative levels of ENSRNOT00000067908, XR_590093, XR_591455, XR_360081, and XR_246933 were significantly increased in SCI rats in comparison with control rats (P
< 0.05; Figure 7A).In contrast, the levels of XR_351404, XR_591426,XR_353833, XR_590076, and XR_590719 were significantly reduced in SCI rats compared with control rats (P
< 0.05; Figure 7B).These results confirm the expression profiles of the lncRNAs obtained by RNA sequencing and offer several promising targets for follow-up studies.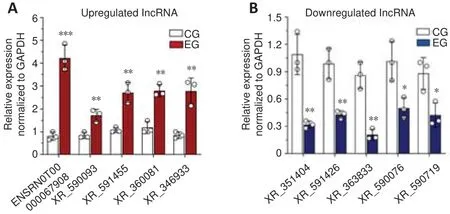
Figure 7 | Verification of differentially expressed lncRNAs by quantitative reverse transcription-polymerase chain reaction.
Coexpressed lncRNA-miRNA-mRNA interaction network
To address the importance of lncRNAs and potential correlations among lncRNAs, miRNAs, and mRNAs, we constructed a lncRNA-miRNA-mRNA coexpression network.As shown in Figure 8, this network included 10 differentially expressed lncRNAs, validated through qRT-PCR as described above.These lncRNAs were coexpressed and interacted with 41 miRNAs and 198 mRNAs (Figure 8).Similarly, individual miRNAs or mRNAs interacted with several lncRNAs or miRNAs in this network.Based on the mechanism of competing endogenous RNA (ceRNA), lncRNAs could serve as a key regulator of miRNA target genes by binding miRNAs (Tay et al., 2014).Of interest,miR-210-3p was found to be coexpressed with XR_591455 and XR_590719.Interestingly, XR_590719 expression was elevated, whereas XR_591455 expression was reduced, as shown in the lncRNA expression profile.Additionally, XR_351404 was found to be coexpressed with rno-miR-219a-5p and rno-miR-34b-5p; XR_353833 was coexpressed with rno-rno-miR-219a-5p;and XR_590076 was coexpressed with rno-miR-347 (Figure 8).Taken together,these bioinformatics data indicate the potentially important functions of the differentially expressed lncRNAs in regulating other genes.
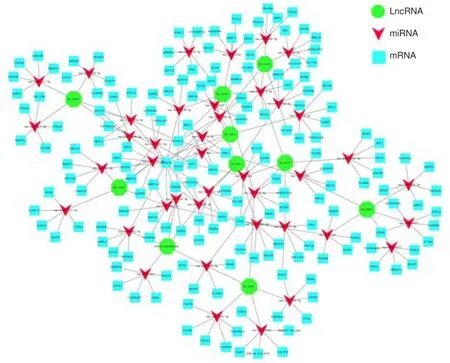
Figure 8| lncRNA-miRNA-mRNA coexpression network.
Discussion
The inflammation that occurs after SCI both protects and damages nerves(Wang et al., 2019b).Similarly, astrocytes not only promote the recovery of injured neurons (Gu et al., 2019), but also hinder neuronal growth and repair of neurons (Okada et al., 2018; Tran et al., 2018).However, exosomes may contribute to the SCI repair by acting as carriers of ncRNAs (Huang et al., 2020; Mu et al., 2021; Zhou et al., 2022).In particular, exosome-derived lncRNAs have been implicated in SCI through different mechanisms (Shao et al., 2020; Wang et al., 2020; Liu et al., 2021).Nevertheless, no study to date has systematically evaluated the differentially expressed lncRNAs that are implicated in SCI development.For this reason, in this study we evaluated differentially expressed exosomal lncRNAs and predicted their possible roles in SCI.Firstly, we successfully established a rat model of SCI,collected exosomes, and isolated high-quality RNA.Next, we performed RNA sequencing to analyze the overall RNA profile.Interestingly, heat maps and volcano maps showed that the SCI rats had significantly differential expression of both lncRNAs and mRNAs compared with the control rats.The enrichment analysis results suggested that these upregulated and downregulated lncRNAs may have important effects on SCI development and even repair.Of note, the top enriched biochemical pathway for upregulated lncRNAs was glycosylphosphatidylinositol (GPI)-anchor biosynthesis, and the top pathway for downregulated lncRNAs was biotin metabolism.Furthermore, the change in expression of 10 circulating differentially expressed lncRNAs was confirmed by qRT-PCR.Finally, a lncRNA-miRNA-mRNA network was constructed using these 10 lncRNAs, five of which appear to be associated with SCI recovery and repair: XR_591455, XR_351404, XR_353833, XR_590719, and XR_590076.Collectively, these findings provide the first comprehensive overview of exosomal lncRNAs involved in SCI in rats and indicate several significantly upregulated or downregulated lncRNAs that could be promising diagnostic and/or therapeutic targets for SCI in the future.
Significant experimental evidence has confirmed that lncRNAs competitively inhibit miRNA expression and thereby play a pivotal role in regulating their target genes, at least partially through the ceRNA mechanism (Wang et al.,2019b).This could also explain why lncRNAs are involved in many diseases.Several lncRNAs, such as lncRNA Xist, lncRNA-scir1, and others, display obviously differential expression after SCI and may be mechanistically associated with neuroinflammation, neuroprotection, scar formation,astrocyte proliferation, and migration (Wang et al., 2015; Huang et al., 2016).Even so, experimental studies of lncRNAs that participate in SCI repair are still limited.Moreover, considering the potential diagnostic and therapeutic value of circulating lncRNAs in the context of SCI (Zhang and Yang, 2017), the objectives of this study were to investigate promising exosomal lncRNAs in an animal model of SCI and predict their potential roles, thereby providing valuable lncRNA targets for clinical diagnosis and treatment.
Construction of a lncRNA-miRNA-mRNA coexpression network showed that XR_351404 and miR-34b-5p were coexpressed in SCI.Intriguingly, Xie and colleagues have shown that inhibition of miR-34b-5p expression may constrain the pulmonary production of proinflammatory cytokines (Xie et al.,2018), suggesting that miR-34b-5p plays a functional role in the inflammatory response.Another study by Liu et al.(2016) demonstrated that miR-34b-5p is pivotal to modulating astrocyte apoptosis.In our study, XR_351404 was found to be both upregulated and coexpressed with miR-34b-5p after SCI.This could shed light on how lncRNAs participate in SCI pathologic progression,considering that lncRNAs serve as ceRNAs for miRNAs.In addition to miR-34b-5p, miR-219a-5p may be a secondary target of XR_351404 and/or XR_353833,as indicated by the lncRNA-miRNA-mRNA interaction network.Given the fact that miR-219a-5p plays a regulatory role in neuronal apoptosis (Yan et al., 2019), we speculate that XR_351404 downregulation may attenuate inflammatory reactions and accelerate astrocyte apoptosis.In combination with the literature, our findings suggest that XR_351404 and XR_353833 may act asbe promising targets for therapeutic modulation of neuronal apoptosis after SCI.
In addition to XR_351404 and XR_353833, which were discussed in detail above, XR_591455 and XR_590719 also attracted our attention, because both of these lncRNAs competitively bind to miR-210-3p, at least partially through the ceRNA mechanism.Interestingly, XR_591455 was downregulated after SCI, whereas XR_590719 was upregulated after SCI, as confirmed via qRT-PCR.Therefore, it is reasonable to speculate that both XR_591455 and XR_590719 contribute to SCI development and recovery by affecting miR-210-3p binding to other target genes.Recent studies found that mesenchymal stem cellderived extracellular vesicles (MSC-EVs) can ameliorate NSCs apoptosis induced by hypoxic injury by upregulating miR-210-3p expression (Li et al.,2020).Thus, the XR_590719/miR-210-3p pair might also be associated with modulation of NSC hypoxic injury of NSCs; although the specific mechanisms of how they contribute to SCI require further investigation.Taking the substantial potential of NSCs in SCI repair into account (Kadoya et al., 2016;Kumamaru et al., 2018), how lncRNA/miRNA pairs affect the fate of NSCs also would be a very intriguing future research direction.Moreover, XR_590076 was found to be coexpressed with miR-347, which functions as a vital factor that aggravates neuronal apoptosis after focal ischemia (Gubern et al.,2013).Therefore, it is possible that XR_590076 inhibits miR-347 and thereby attenuates neuronal loss.
This study had some limitations.In terms of methodology, we only used female rats in this study; to substantiate the observed phenotypes, rats of both sexes should be used.In addition, even though the number of rats that we used was in line with many published studies (Marcol et al., 2015; Zhou et al., 2020), the sample size should be increased.Moreover, this study did not address the clinical significance of these differentially expressed exosomal lncRNAs in blood from patients with SCI.Finally, this study did not encompass anyin vitro
andin vivo
functional evaluation of these lncRNAs, which is of importance indeed.In conclusion, this is the first study to screen for differentially expressed exosomal lncRNAs in a rat model of SCI.RNA sequencing was performed,the expression changes of ten important lncRNAs, including several lncRNAs related to spinal cord repair, were verified by qRT-PCR, and an interaction network of lncRNAs, miRNAs, and mRNAs was constructed to predict their potential roles in SCI recovery.Our findings suggest that circulating exosomal lncRNA-mediated changes to the microenvironment may contribute to the development of rat SCI.
Author contributions:
Study design and conception: JAL, MPS, CFZ, WFW;experiment implementation, data collection and analysis: JAL, MPS, LC, MYG,YHC, SYW, ZHL; data interpretion: JAL, MPS, CFZ, WFW; manuscript draft: JAL,MPS, CFZ, WFW.All authors read and approved the final manuscript.
Conflicts of interest:
None of the authors has any potential conflict of interest.
Open access statement:
This is an open access journal, and articles are distributed under the terms of the Creative Commons AttributionNonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.

- 中国神经再生研究(英文版)的其它文章
- Neural and Müller glial adaptation of the retina to photoreceptor degeneration
- Agomelatine: a potential novel approach for the treatment of memory disorder in neurodegenerative disease
- MicroRNAs: protective regulators for neuron growth and development
- In vivo astrocyte-to-neuron reprogramming for central nervous system regeneration: a narrative review
- Intranasal nerve growth factor for prevention and recovery of the outcomes of traumatic brain injury
- Altered O-GlcNAcylation and mitochondrial dysfunction,a molecular link between brain glucose dysregulation and sporadic Alzheimer’s disease