
In vivo astrocyte-to-neuron reprogramming for central nervous system regeneration: a narrative review
2022-09-16 04:19ZuliyaerTalifuJiaYiLiuYunZhuPanHanKeChunJiaZhangXinXuFengGaoYanYuLiangJieDuJianJunLi
中国神经再生研究(英文版) 2023年4期
Zuliyaer Talifu , Jia-Yi Liu Yun-Zhu Pan , Han Ke Chun-Jia Zhang Xin Xu Feng Gao ,Yan Yu Liang-Jie Du Jian-Jun Li ,
Abstract The inability of damaged neurons to regenerate within the mature central nervous system (CNS)is a significant neuroscientific challenge.Astrocytes are an essential component of the CNS and participate in many physiological processes including blood-brain barrier formation, axon growth regulation, neuronal support, and higher cognitive functions such as memory.Recent reprogramming studies have confirmed that astrocytes in the mature CNS can be transformed into functional neurons.Building on in vitro work, many studies have demonstrated that astrocytes can be transformed into neurons in different disease models to replace damaged or lost cells.However, many findings in this field are controversial, as the source of new neurons has been questioned.This review summarizes progress in reprogramming astrocytes into neurons in vivo in animal models of spinal cord injury, brain injury, Huntington’s disease, Parkinson’s disease, Alzheimer’s disease, and other neurodegenerative conditions.
Key Words: astrocyte; astrocyte-to-neuron; central nervous system; in vivo; nerve regeneration;neurological disorders; reprogramming; review
Introduction
Damaged neurons in the adult mammalian central nervous system (CNS)show an inability to regenerate and form new functional connections.As a result, most CNS injuries and diseases cause permanent disability (Curcio and Bradke, 2018).The ability of the CNS to undergo the same robust regeneration and axonal growth observed during embryonic development would allow for novel, more effective treatments of neurodegenerative conditions and CNS injuries.Stem cell transplantation was once thought to be the future of neural repair and was expected to promote production of neurons that could restore function.However, stem cell use is subject to ethical controversy, and immune rejection and the inability of differentiated cells to functionally integrate into the CNS have hindered use of this technology (Li and Chen, 2016).Therefore,focus has shifted to repairing neural function through endogenous cells.
Historically, glial cells in the CNS were believed to primarily support and protect neurons.Advances in molecular biology and genetics, and advances in characterization of morphology and physiology have provided a more indepth understanding of glial cells, and significant progress has also been made in the field of neuronal transdifferentiation (Wang et al., 2015; Gascón et al., 2017; Yu et al., 2020).Astrocytes are the most widely distributed and largest glial cells in the mammalian nervous system.In addition to providing physical and metabolic support to neurons and isolating the CNS from the systemic circulation by forming the blood-brain barrier (BBB), astrocytes also participate in synaptic development, maturation, and pruning.The role of astrocytes in nervous system repair has received increasing attention(Clarke and Barres, 2013; Tai et al., 2020; Gӧtz and Bocchi, 2021).This review is focused on research progress in repair of central nervous system injury through astrocyte transformation into neuronsin vivo
.Search Strategy
We performed electronic searches using PubMed, Medline, and Google Scholar databases for literature published in English before March 2022.The keywords used for the literature search were: Astrocyte ANDin vivo
AND reprogramming OR transdifferentiation.The results were further screened by heading and abstract.Astrocyte Functions and Reprogramming
The physiological functions of astrocytes
Glial cells are the second-most abundant component of the mammalian nervous system after neurons.Although they do not produce electrical impulses, they participate in CNS activities in a variety of ways.The four primary types of glial cells in the human CNS are astrocytes, oligodendrocytes,microglia, and ependymal cells.Each of these four cell types affect neurons in different ways.Glial cells regulate synapse formation (Farhy-Tselnicker and Allen, 2018), participate in physiological activities such as breathing (Beltrán-Castillo et al., 2017), remove dead neurons and pathogens (Clarke and Barres,2013), and are involved in learning and other advanced cognitive functions(Fields et al., 2014).Astrocytes are the most widely-distributed type of glial cells in mammals, and are also the largest (Freeman and Rowitch, 2013).In addition to providing physical and metabolic support, and isolating the CNS through BBB formation, astrocytes are involved in synapse development,maturation, and pruning.As their list of known functions has grown, their potential for repairing the nervous system has received increased attention(Clarke and Barres, 2013).
When the CNS is damaged, glial cell activity increases, and glial scars form in a process called “reactive gliosis” to prevent further damage to adjacenttissues.Although glial cells can secrete substances that promote neural repair,glial scars create physical obstacles to axon regeneration (Pekny and Nilsson,2005; Hara et al., 2017; Boghdadi et al., 2020).Therefore, proliferation of reactive glial cells proliferation is a double-edged sword for the CNS.Simply inhibiting glial proliferation slows recovery (Burda and Sofroniew, 2014).Identification of mechanisms to promote nerve repair while eliminating scars would significantly improve CNS structural and functional recovery.
Discovery of astrocyte reprogramming potential
In 1999, Doetsch et al.reported the existence of neural stem cells with differentiation potential in the subventricular area of adult mammals.Astrocytes in this area can self-renew and act as stem cells to generate immature precursors and neuroblasts (Doetsch et al., 1999).These intriguing findings raised many questions.Why do glial cells in specific areas function as neural stem cells? Can this feature be leveraged to promote nerve injuryrepair? Researchers have attempted to induce migration of astrocytes in these regions to injured sites (Alvarez-Buylla and GarcA-Verdugo, 2002; Cayre et al., 2006; Mamber et al., 2010).However, due to the limited number of astrocytes in the subventricular area, and the difficulty in precisely regulating their migration, the efficacy of this method for treating serious diseases is unclear.
In the 21century, researchers have moved beyond the subventricular area and hippocampus and begun to ask whether astrocytes elsewhere in the CNS could transform into neurons.In vivo
andin vitro
studies have shown that glial cells can differentiate into neurons through activation of transcription factors and through drug treatments (Li and Chen, 2016; Qin et al., 2017; Tai et al.,2020).In 2012, the Nobel Prize in Physiology or Medicine was awarded to Sir John B.Gurdon and Shinya Yamanaka for their discovery that mature cells can be reprogrammed to become pluripotent.In addition, development of viral vector technologies such as adenoassociated viruses (AAVs) and retroviruses, and new theoretical and experimental methods, have enabled rapid advances in the field of cell reprogramming (Verdera et al., 2020; Muhuri et al., 2021).Breakthroughs have been made in treating neurological diseases by reprogramming cells and transforming them into neuronsin vitro
andin vivo
.Astrocyte Reprogramming and Transdifferentiation into Neurons
The main difference between reprogramming and transdifferentiation is whether protocells become pluripotent stem cells.During reprogramming,protocells dedifferentiate into pluripotent stem cells, then differentiate into specific cell types via retroviruses (Chen et al., 2017), AAV viruses (Chen et al., 2020), and small molecule drugs (Qin et al., 2017).In the process of transdifferentiation, protocells are directly induced into target cells without becoming pluripotent stem cells (Xie et al., 2017).
Animal studies of traumatic central nervous system injury and degenerative central nervous system disease have shown that regulating astrocyte fate through viral vector-mediated overexpression of crucial transcription factors can convert a subset of astrocytes into neurons in situ following CNS damage.These transformed neurons can connect with the existing neural network and produce spontaneous electrical activity (Figure 1 and Table 1).
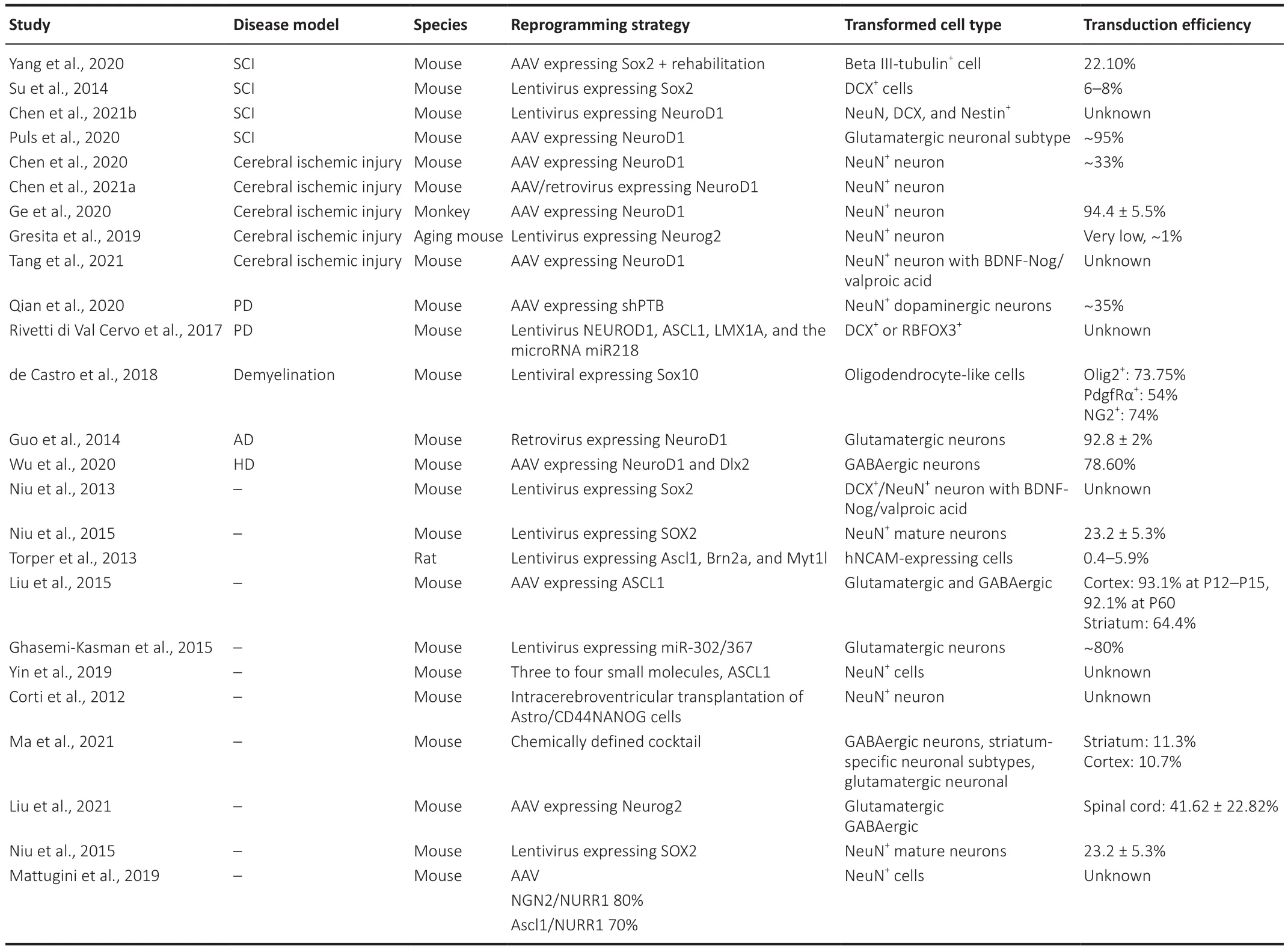
Table 1 |Summary of included studies of in vivo astrocyte reprogramming
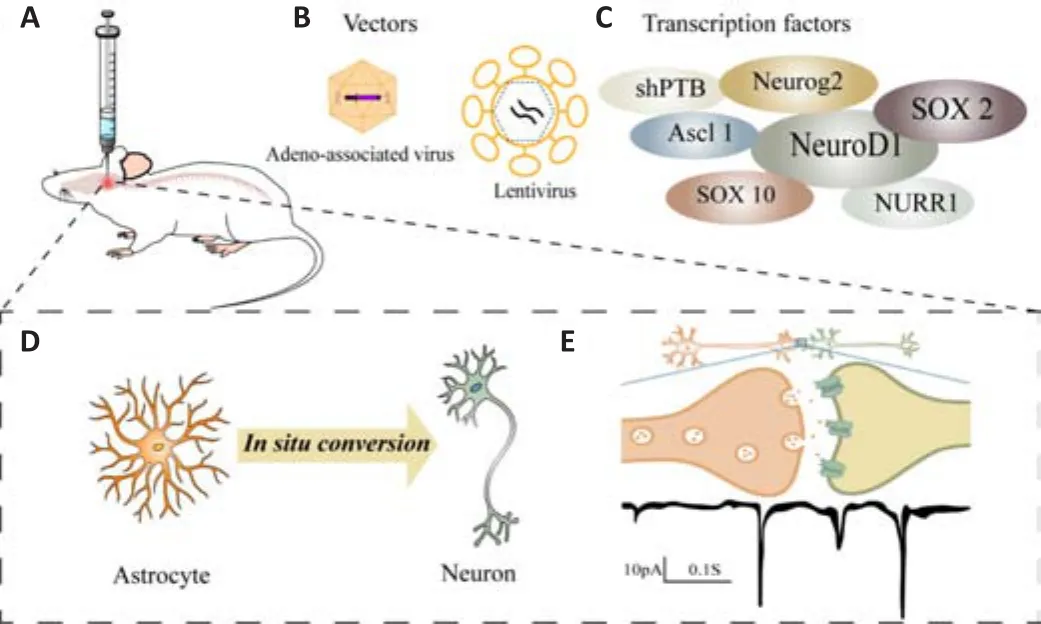
Figure 1 | Schematic depicting the in vivo astrocyte conversion strategy.
Astrocyte reprogramming after brain injury
Non-specific neuronal death caused by external forces or ischemia and hemorrhage is the leading cause traumatic brain injury and stroke-induced dysfunction.The prevalence of TBI was 55.50 million in 2016, and caused 8.1 million years of life lived with disability (YLDs).In addition, stroke caused 6.1673 million all-aged death (GBD 2017 Causes of Death Collaborators, 2018;GBD 2016 Traumatic Brain Injury and Spinal Cord Injury Collaborators, 2019).Replacement of dead neurons with reprogrammed astrocytes may be a novel therapeutic avenue.
The Sox (sex-determining region Y) family of transcription factors plays an essential role in mammalian embryo development, and Sox2 is necessary for maintaining embryonic stem cell pluripotency (Sarkar and Hochedlinger,2013).Many studies have examined how the Sox family of transcription factors affects astrocyte reprogramming.Injection of lentivirus overexpressing Sox2 into mouse brains resulted in reprogramming of astrocytes in the striatum into neural progenitors.Further induction by brain-derived neurotrophic factor (BDNF)/BDNF noggin or histone deacetylase inhibitor resulted in transformation of DCX/ASCL1neural progenitors into NeuNneurons that persisted for as long as 32 weeks after injection (Niu et al., 2013,2015).
NeuroD1 (neurogenic differentiation 1) is a neural transcription factor that participates in initiation of neural differentiation.It plays a vital role in embryonic brain development and maintenance of normal peripheral nervous system (PNS) and CNS functions (Cho and Tsai, 2004; Jahan et al., 2010; Lai et al., 2020).Several studies reported that NeuroD1 was closely related to astrocyte transdifferentiation (Chen et al., 2017, 2020; Puls et al., 2020; Tang et al., 2021).When NeuroD1 is overexpressed in the brains of mice with ischemic brain injury, the number of damaged neurons is reduced by onethird, and another one-third of the damaged neurons are regenerated by glial cells.After 2 months, the motor and cognitive functions of the treatment group were significantly improved compared with those of the control group (Chen et al., 2020).Similarly, Xiang et al.(2021) showed that NeuroD1 overexpression with a retroviral or AAV vector transformed astrocytes into neurons in the cortices of mice with ischemic stroke.In this study, the investigators suggested avoiding high concentrations of injected virus to avoid “artifacts” (Xiang et al., 2021).A study using a rhesus monkey cerebral ischemia model showed that astrocytes were efficiently transformed into neurons by an AAV carrying NeuroD1 with a > 90% conversion efficiency (Ge et al., 2020).These positive results in primates are an important step toward clinical application.
Astrocyte reprogramming in Huntington’s disease
Huntington’s disease (HD) is caused by a gene mutation that results in loss of gamma-aminobutyric acid (GABAergic)-releasing neurons in the striatum,leading to motor, cognitive, and mental dysfunction.The population incidence of HD varies from 0.02 to 1 per 10,000 (Rikani et al., 2014; Rawlins et al.,2016).Transformation of astrocytes into GABAergic neurons to supplement lost neurons is a promising example of reprogramming to treat HD.When NeuroD1 and Dlx2 are overexpressed in the striatum of HD model mice,astrocytes can be reprogrammed into GABAergic neurons.These transformed neurons show similar electrophysiological characteristics as the original neurons and reduced dysfunction and prolonged the lives of model mice (Wu et al., 2020).
Astrocyte reprogramming in Parkinson’s disease
Parkinson’s disease (PD) is a neurodegenerative disease characterized by progressive loss of dopaminergic neurons in the striatum (Kalia and Lang,2015).The goal of astrocyte reprogramming to treat PD is to transform astrocytesin situ
into dopaminergic neurons in the striatum to replenish the lost neurons and prevent disease progression.Regulation of transcription factors and microRNAs are typically used to replenish dopaminergic neurons(Wei and Shetty, 2021).Astrocytes can be reprogrammed into dopaminergic neurons using lentiviral vectors that overexpress NeuroD1, Ascl1, and Lmx1a, and microRNA 218.The results from 2 weeks to 15 weeks after injection showed that these transformed neurons gradually matured and generated action potentials.Furthermore, these PD model mice showed a degree of improvement in motor function (Rivetti di Val Cervo et al., 2017).Another study showed that injection of an AAV vector carrying RNA targeted CRISPR/CasRX knockdown polypyrimidine trace binding protein 1 (PTBP1)into the striatum of PD model mice induced transformation of astrocytes into functional neurons with specific dopaminergic neuron markers.These neurons produced electrical activity and improved the motor function of the PD mice in this study (Zhou et al., 2020).Recently, Giehrl-Schwab et al.(2022) reported that an AAV-based intein split dcas9 activator system (AAV DCAS) carrying different combinations of Ascl1, Lmx1a, Nr4a2 (ALN)/Ascl1,Lmx1a, NeuroD1, and miRNA218 (ALNe-218) could reprogram astrocytes into GABAergic neurons, resulting in improved motor function in mice.In addition to astrocytes, fibroblast reprogramming is also a major research hotspot in PD research (Yavarpour-Bali et al., 2020).
Astrocyte reprogramming in Alzheimer’s disease
Alzheimer’s disease (AD) is a common neurodegenerative disease characterized by neuronal and synaptic loss in the cerebral cortex and subcortical areas (Wenk, 2003).Overexpression of NeuroD1 in the cortices of AD model mice using a retrovirus resulted in reorganization of reactive glial cells into functional glutamatergic neurons.These neurons produced spontaneous synaptic responses and induced postsynaptic reactions, which indicated that the resulting neurons were successfully integrated into the local neural network (Guo et al., 2014).That study also showed that NG2 cells were reprogrammed to glutamatergic and GABAergic neurons (Guo et al.,2014).
Astrocyte reprogramming after spinal cord injury
Spinal cord injury (SCI) interrupts communication between the CNS and the PNS.Axonal rupture and neuronal death caused by various physical and chemical factors are the fundamental causes of sensory, motor, and autonomic nervous system dysfunction (Zhang et al., 2021).The goal of astrocyte reprogramming in SCI is to replace dead neurons with transformedneurons and restore function by forming functional connections with the original system (McDonald and Sadowsky, 2002).Like other CNS injuries, SCI can induce glial scar formation through reactive gliosis mediated by astrocytes(Hara et al., 2017).Furthermore, astrocyte proliferation after SCI is more extensive than following brain injury, so glial cells may play a more critical role in the pathological process of SCI (Boghdadi et al., 2020).For these reasons,glial cell reprogramming after SCI is a research hotspot (Schnell et al., 1999;Bradbury and Burnside, 2019).
When a lentiviral vector overexpressing NeuroD1 was delivered to the spinal cords of SCI rats, the numbers of neural stem cell marker-labeled cells, immature neurons, and mature neurons in the spinal cord all increased significantly (Chen et al., 2017).Similarly, when an AAV was used to overexpress NeuroD1 in the spinal cords of SCI model mice, astrocytes were transformed into neurons (Puls et al., 2020).Interestingly, when NeuroD1 alone was used for induction, most of the transformed neurons were glutamatergic.When Dlx2 was included with NeuroD1, most of the transformed neurons were GABAergic.Electrophysiological techniques were used to confirm that the transformed neurons participated in physiological functions (Puls et al., 2020).These results suggested that multiple transcription factors are essential to induce different types of target cells that release different neurotransmitters.
The role of the transcription factor Sox2 was also studied in astrocyte reprogramming.Sox2 alone was sufficient to reprogram endogenous astrocytes to neuroblasts and mature neurons in the adult mammalian spinal cord (Su et al., 2014).In addition, new neurons were able to form synapses with pre-existing motor neurons, which indicated that the newborn neurons integrated into the local neural network of the injured spinal cord and exerted physiological functions (Su et al., 2014).A recent study found that endogenous Sox2 could transform NG2 glial cells (not astrocytes) into neuroblasts following SCI, but the low levels present were not sufficient to induce development of these neuroblasts into mature neurons (Tai et al., 2021).However, addition of Sox2 via exogenous means resulted in development of neuroblasts into mature neurons.
Glial reprogramming for vision restoration
Vision, an advanced brain function, can input information from the objective world into the brain.The death of retinal ganglion cells and photoreceptors is the leading cause of irreversible visual impairment.Restoration of visual function via reprogramming of glial cells would represent a significant medical advancement (Blackshaw and Sanes, 2021).Overexpression of the NeuroD1 gene in the brains of mice with cerebral ischemia resulted in reprogramming of astrocytes into functional neurons that responded to visual stimuli.Within 3-6 weeks after reprogramming, gradual maturation and functional integration of synapses of the transformed neurons was observed, and this process was similar to cortical circuit development (Tang et al., 2021).
Müller glial cells are the primary type of glial cell in the retina, and provide structural support for neurons.Reprogramming retinal Müller glial cells and oligodendrocytes through overexpression is a potential strategy for restoration of visual function (Sekiryu and Matsuda, 2021).A study showed that knockdown of polypyrimidine trace binding protein 1 (ptbp1) using an AAV vector carrying RNA targeted CRISPR/casrx resulted in efficient transformation from Müller glial cells to retinal ganglion cells, which then underwent functional integration.This resulted in reduced symptoms associated with RGC loss in a retinal injury mouse model (Zhou et al., 2020).The regulatory transcription factors Math5 and Brn3b were able to reprogram Müller glia in mice to retinal ganglion cells that had typical neuronal electrophysiological characteristics and improved the visual function of the mice (Xiao et al., 2021).Interestingly, the AAV-specific promoter used in this study was GFAP (typically considered an astrocyte-specific promoter), but evaluation of astrocyte-specific markers and lineage tracing confirmed that the induced cells were indeed Müller glia and not astrocytes (Xiao et al.,2021).
In addition, regeneration and axon formation of retinal ganglion cells can also be induced by pharmacological reprogramming (Mahato et al.,2020) and regulation of DNA methylation mediated by transcription factor overexpression (Lu et al., 2020).These findings may provide an essential theoretical basis for restoration of visual function through reprogramming glial cells.
Challenges and Perspectives
Identifying the source of newborn neurons
Although many previous studies have shown that astrocyte reprogramming can be accomplished using many different strategies (Lentini et al., 2021; Ma et al., 2021; Zhou et al., 2021), recent studies have questioned the source of regenerated neurons.Tai et al.reported that functional neurons could be restored through exogenous overexpression of theSox2
gene.However, the source was not astrocytes, but NG2 glial cells (Tai et al., 2021).Another study reported extremely low transfection efficiency after viral vector treatment,especially in an aged animal model (Gresita et al., 2019).NG2 glial cells are oligodendrocyte precursor cells that proliferate throughout their lifetime and participate in neuronal information transmission (Dimou and Gӧtz, 2014).The ability of NG2 cells to proliferate throughout their lifetime is a trait of stem and progenitor cells, and indicates that these cells may be attractive targets for reprogramming.A significant controversy in the field of glial reprogramming is whether the source of reprogrammed neurons is NG2 glial cells or astrocytes.According to Heinrich et al.(2014),DCX positive neuroblasts were obtained by lentiviral overexpression of Sox2 in cortical stab model mice, and the source was traced using fat genetic mapping.More than 60% of the cells were derived from NG2 cells, and only a small proportion of the reprogrammed cells were derived from reactive astrocytes.
The gold standard for direct reprogramming in vivo: Lineage tracing
In the past decade, direct induction of neurons to treat diseases has shown promise in various animal injury models.However, the primary problem remains identification of the source of induced neurons and confirmation that the induced neurons exert physiological functions.Multi-dimensional evaluation of neuronal induction and morphology requires information transmission ability to be strictly defined as the “spectrum of neurons” (Yang et al., 2011).Therefore, regardless of intervention, the possible influence of the body’s internal regeneration mechanisms must be ruled out.The most serious challenge facing this field may be to clarify that “newborn neurons”generated by different regulatory means actually come from astrocytes, and eliminating other possible sources as the progenitors of these neurons.This problem is a primary focus in the field of astrocyte reprogramming.
Wang et al.(2021a) reported that neither AAV-induced NeuroD1 overexpression nor Ptbp1 gene knockdown induced transformation of astrocytes into neurons.Instead, the regenerated neurons were neurons that were already present, and leakage of the AAV likely caused the reprogramming of neurons reported in previous studies, but the mechanism is unclear (Wang et al., 2021a).In that study, Wang et al.(2021a) found that the commonly used viral vector AAV, even with the commonly used astrocytespecific promoter GFAP, had significant differences in the specificity of vector transfection for different serotypes.After 4 days ofin vivo
transfection,different serotypes mainly transfected astrocytes, but after 14 days, the commonly used serotypes 8, 9, and PHP.eB transfected significantly less astrocytes, and the expression of fluorescent protein began to appear in many pre-existing neurons (Wang et al., 2021a).Similarly, Chen et al.(2022)recently reported that pedigree tracking of expression of ptbp1 showed that reactive astrocytes did not transform into neurons in the brains of PD model mice.Furthermore, the previously reported new neurons may have been neurons originally.In addition, some researchers have expressed concern that direct transformation of astrocytes may negatively impact the nervous system.At the same time, due to the problems of promoter leakage and specificity in the AAV-based gene regulation system, it should not be applied to the proof of lineage (Svendsen and Sofroniew, 2022).Recently, Giehrl Schwab et al.(2022) reported that multiple transcription factors were overexpressed simultaneously in PD model mice following treatment with an AAV-based intein split dcas9 activator system (AAV DCAS), and claimed that this improved AAV vector could eliminate ambiguity with regard to the source of new neurons.Strict lineage tracking has received increased attention in the field of direct astrocyte reprogrammingin vivo
(Wang and Zhang, 2022).Therefore, a more specific and safer delivery system, and well-designed experiments, are needed to clarify the source of new neurons.Optimizing reprogramming strategies
Single agonists or blockers may not provide adequate therapeutic benefit due to differences in CNS disease pathogenesis, damaged cell types, and physiological environments.Specific reprogramming strategies need to be formulated for various conditions, and complex therapeutic schemes for multiple targets need to be explored to achieve the best curative effects (Liddelow and Barres, 2017).Analysis of specific transcriptional and epigenetic characteristics of different regions and neurons may improve the efficacy of reprogramming (Herrero-Navarro et al., 2021).Transcriptome analysis and pathway verification will aid in identification of critical factors that regulate glial cell reprogramming, targeted small molecule drug research and development will aid in implementation of clinical interventions (Ma et al., 2019).Although the conversion process from living glial cells to neurons has been characterized, there are significant differences in reprogramming efficiencies between white matter and gray matter (Liu et al., 2020).In addition, damage to the CNS results in induction of two types of reactive astrocytes: A1 and A2.A1 astrocytes may promote neuron death in neurodegenerative diseases, and A2 astrocytes exert neuroprotective effects(Liddelow et al., 2017).Different lineages and local microenvironments of glial cells regulatedin vivo
transformation of glial cells into neurons (Mattugini et al., 2019).Therefore, it is necessary to determine the causes and consequences of these differences to maximize the neuroprotective effects of astrocyte and to inhibit potential negative effects of reprogramming.Use of an appropriate control vector is critical for developing a reprogramming strategy.At present, lentivirus or AAV virus vectors are used in the field of live weight programming for regulation, but the reprogramming efficiency has differed across studies, as summarized in Table 1.Adenoassociated virus vectors are mostly used in animal experiments because they are easy to prepare, have a high transfection rate, and induce little host immune response (Liddelow et al., 2017).Adeno-associated virus-mediated gene therapy has shown great potential for treatment of systemic diseases,such as those that affect the cardiovascular system (Shi et al., 2020; Cao et al., 2021), hereditary diseases (Kodippili et al., 2018; Nguyen et al., 2021), and immune system diseases (Priddy et al., 2019).The biggest obstacle to use of AAV vectors in central nervous system diseases is inefficient penetration of the blood-brain-spinal cord barrier and difficulty inducing local overexpression of target genes.Development of clinical applications requires design of a reliable astrocyte-targeted regulatory system that is non-integrated and can be temporally and spatially expressed (Torper et al., 2013).
The proportions of various cells in the nervous system are directly related to the nervous system’s evolution, development, aging, and pathological processes (Herculano-Houzel, 2011).The glia-neuron ratio (GNR) refers to the ratio of the number of glial cells to the number of neurons in the same volume of brain parenchyma.Since the 1960s, about it was widely believed that there were 1 trillion glial cells and 100 billion neurons in the human brain, which corresponded to a GNR of ~10:1.More recent data obtained using new cell counting methods showed that the total number of glial cells and neurons in the each human brain is less than 10 billion, and the GNR is~1:1 (von Bartheld et al., 2016).Although the actual GNR is not known, GNR stability is essential for normal physiological function of the CNS.Prior to artificial manipulation of the GNR, high-quality research must be performed to determine whether this manipulation will affect normal CNS function,including the integrity of the blood-brain barrier and accuracy of electrical activity.
In traumatic injuries of the CNS, the ideal time window for using cell reprogramming to repair damage may be after primary scar formation and injury inhibition by reactive glial cells.The purpose of reprogramming reactive glial cells into functional neurons is to revert glial scars back to neural tissue and eventually recover lost neuronal function.In the early stage of damage and disease, the microenvironment will be in a relatively unstable state due to acute stress reactions such as inflammation (Ahuja et al., 2017).In the chronic stages, the entire CNS may have undergone adaptive changes.Therefore,it is necessary to determine the optimal treatment windows for different pathological states.In some degenerative diseases, harmful factors such as ROS and inflammatory cytokines may be present throughout the nervous system, and even neurons produced by reprogramming may degenerate(Wang et al., 2021b).Therefore, more high-quality research is needed on how to get the best therapeutic effect of reprogramming in different pathological environments.
Limitations
This review was subject to some limitations.First, we mainly searched PubMed, MEDLINE, and Google Scholar databases.Although these databases are widely used, some relevant literature may have been omitted.Second,we included manuscripts published up to March 2022.Therefore, ongoing research and new results were not included in this review.Finally, the manuscripts reviewed in this paper were only published in English language journals.
Conclusion
Astrocytes have been successfully reprogrammed into different types of neurons through pharmacological approaches and regulation of various transcription factors.Basic research has led to significant progress in improving animal models of disease.Future studies of the mechanisms and pathways of astrocyte reprogramming are needed, as identification of critical pathways will facilitate development of methods for safer,controllable human cellular reprogramming.Pathological CNS conditions are characterized by damage to and loss of diverse neuron subtypes in different microenvironments, so it is necessary to identify the best regulatory methods in these varied scenarios.In vivo
experiments to control the proportion of glial cells converted into neurons are also needed.The GNR remains within a specific range under physiological conditions.It is unclear if artificially changing the balance will lead to CNS hyperactivity and induce epilepsy or other symptoms, so long-term experiments should be performed (Goldman,2016).We must carefully evaluate the long-term effects of transformed neurons before applying this technology to treat human diseases.Developing the potential of transforming glial cells into neurons changes our understanding of the CNS and has the potential to open a new research field.However, the road to clinically treat CNS diseases using this approach will be long.We need to advance the development of this field in terms of clarifying the source of reprogramming cells, obtaining safer and more efficient gene vectors, and optimizing reprogramming schemes for different diseases.
Author contributions:
Design and concepts: JJL, FG, ZT; definition of intellectual content: YY, LJD, ZT; literature search: ZT, JYL, YZP, HK; data acquisition: ZT, CJZ, XX; manuscript preparation: ZT, JYL; manuscript editing:JYL, YZP, FG; manuscript review: JJL, YY.FG.All authors approved the final version of the manuscript.
Conflicts of interest:
None declared.
Open access statement:
This is an open access journal, and articles are distributed under the terms of the Creative Commons AttributionNonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.

- 中国神经再生研究(英文版)的其它文章
- Neural and Müller glial adaptation of the retina to photoreceptor degeneration
- Agomelatine: a potential novel approach for the treatment of memory disorder in neurodegenerative disease
- MicroRNAs: protective regulators for neuron growth and development
- Intranasal nerve growth factor for prevention and recovery of the outcomes of traumatic brain injury
- Altered O-GlcNAcylation and mitochondrial dysfunction,a molecular link between brain glucose dysregulation and sporadic Alzheimer’s disease
- Signaling interactions among neurons impact cell fitness and death in Alzheimer’s disease