
响应面法优化产木聚糖酶耐热菌株的发酵条件
2022-09-13 14:18:42王瑞君
宜春学院学报 2022年3期
王瑞君,袁 欣
(宜春学院 化学与生物工程学院,江西 宜春 336000)
半纤维素是自然界中含量仅次于纤维素的可再生资源,半纤维素的主要成分为木聚糖,是构成植物细胞壁的主要成分之一,占植物细胞干重的30-40%。[1]自然界中木聚糖含量丰富,但因其难以降解而无法被人类充分利用。在高等植物中木聚糖是主链以木糖残基通过β-1,4糖苷键连接而成的线性分子,木聚糖的降解离不开木聚糖酶。
木聚糖酶是一类能够降解木聚糖为木寡糖、木糖的复合酶系,木聚糖酶可应用于能源、饲料、医药、食品、造纸、生物脱胶等领域。[2-3]木聚糖酶加入面粉中可以延缓了面包的水分迁移速率和失水速率、增强面包抗老化能力。[4]王茜等[5]在研究复合菌群脱胶效果时发现,脱胶效果较好的复合菌系的组合中木聚糖酶的活力也较高。木聚糖酶可用于纸浆漂白,芬兰森林公司在1988年首次进行木聚糖酶工厂化制浆实验。[6]木聚糖酶来源广泛,可来源于动物、植物、微生物。[7-8]因微生物产生的木聚糖酶因具有多样性和微生物生长周期短等优点,应用于工业生产的木聚糖酶大部分来源于微生物。[9-10]
产木聚糖酶菌株的环境来源广泛,可来源于土壤[11]、也可来源于水体[12],或者腐烂的植物[13]以及某些动物的肠道[14]。本文从江西宜春温汤温泉水淤泥中分离到一株产耐热木聚糖酶菌株,对产酶条件进行优化,有利于为木聚糖酶的进一步工业化利用奠定基础。
1 材料与方法
1.1 材料
1.1.1 试剂
木聚糖(sigma)、玉米芯木聚糖(成都)、燕麦木聚糖(sigma)、麸皮(购于农贸市场),其他主要试剂购于国药集团。
1.1.2 培养基
普通营养琼脂培养基:牛肉膏0.3%,蛋白胨1%,NaCl 0.5%,琼脂2%。
木聚糖酶菌株筛选培养基:普通营养琼脂培养基的基础上添加1%的木聚糖。
基础产酶培养基:麸皮15 g/L,蛋白胨5 g/L,1000 mL蒸馏水。
1.1.3 DNS溶液[15]
10g 3,5-二硝基水杨酸、16g NaOH、5.0g苯酚、5.0g亚硫酸钠、300g酒石酸钾钠、超纯水定容1L。
1.2 方法
1.2.1 菌株富集及筛选
从江西宜春温汤的温泉中取带有淤泥的温泉水,混匀,50 mL加入无菌锥形瓶中,同时加入1 g木聚糖,放入摇床中,55℃,180 r/min,培养12 h进行产木聚糖酶菌株的富集。
富集液10倍梯度进行稀释,取合适浓度的稀释液100 μL涂布于木聚糖酶菌株筛选培养基,放于生化培养箱,在55℃下培养24h。测量并记录各菌落直径d1、透明圈直径d2,计算出透明圈与菌落直径之比(d2/d1)。挑取透明圈与菌落直径之比较大的菌株在普通营养琼脂培养基平板上划线,55℃下培养24h。重复上述操作3次,最后挑取单菌落于营养肉汤培养基中,液体培养纯化菌株,以用于后续的试验。
1.2.2 木糖标准曲线的制备
按表1加入不同体积的1 mg/mL木糖溶液和蒸馏水后,在各试管中加入1 mL的DNS试剂,煮沸5 min,流水迅速冷却,加入8 mL的蒸馏水,混匀,测定540 nm波长处的光吸收值。以吸光度值为纵坐标,木糖浓度(质量)为横坐标,绘制木糖标准曲线。得出标准曲线为Y=1.982X-0.03,(R2=0.999)。

表1 木糖标准曲线溶液的配制
1.2.3 木聚糖酶酶活力的测定
将发酵液在10000 r/min、4℃条件下离心5 min,所得上清液为粗酶液。木聚糖酶粗酶液200 μL加入到800 μL的用柠檬酸-柠檬酸钠缓冲液配制的1%木聚糖底物溶液中,60℃水浴反应5 min,采用DNS法测定所释放的还原糖。
酶活力单位(U)定义为:在合适的条件下,单位时间(以每分钟表示)内水解木聚糖释放1μg还原糖(以木糖表示)所需的酶量为1个酶活力单位。[16]
即:酶活力(U)=1000M × N/(T×L)
公式中M为还原糖(木糖)的质量(mg),N代表酶液稀释倍数,T为反应时间(min),L为反应酶液的体积(mL)。
1.2.4 酶学性质
最适温度:木聚糖溶于pH5.5的0.1M的柠檬酸-柠檬酸钠缓冲液中,配制1%的木聚糖底物溶液。800μL的底物溶液分别放置于45℃、50℃、55℃、60℃、65℃、70℃温度条件下保温10min后添加200μL粗酶液在相应的温度条件下反应5min,测定酶活。
最适pH:配制pH5、pH5.5、pH6、pH6.5的0.1M的柠檬酸-柠檬酸钠缓冲液与pH值为7.0的磷酸盐缓冲液。木聚糖溶于不同酸碱度的缓冲液中配制不同pH的1%的木聚糖底物溶液。60℃测定不同pH值条件下的木聚糖酶活。
1.2.5 单因素试验
最适碳源及最适添加量:在产酶培养基的氮源为蛋白胨、pH7.0±0.2的基础上,改变碳源,分别添加10g/L的麸皮、燕麦木聚糖、玉米芯木聚糖、苎麻半纤维素,接种2%发酵液,在60 ℃,180r/min的条件下发酵24h,然后测定发酵液的酶活,确定最佳碳源。确定最佳碳源后,在氮源不变的基础上,改变最适碳源的浓度,分别设定添加量为5g/L、10g/L、15g/L、20g/L、25g/L、30g/L,60 ℃,180r/min的条件下发酵24h,然后测定发酵液的酶活,确定最佳碳源的最适浓度。
最适氮源及最适添加量:在产酶培养基的碳源为麸皮、pH7.0±0.2的基础上,改变氮源,分别添加1%的酵母浸粉、蛋白胨、尿素、NH4Cl,接种2%发酵液,在60 ℃,180r/min的条件下发酵24h,然后测定发酵液的酶活,确定最佳氮源。确定最佳氮源后,在碳源不变的基础上,改变最适氮源的浓度,分别设定添加量为1g/L、2g/L、3g/L、4g/L、5g/L、6g/L、7g/L,60 ℃,180r/min的条件下发酵24h,然后测定发酵液的酶活,确定最佳氮源的最适浓度。
发酵时间的检测:利用最佳发酵培养基发酵,培养12h后,每间隔4h取10mL发酵液,离心后测定上清液的酶活,确定最适发酵时间。
1.2.6 响应面实验设计
分析单因素试验的数据,以木聚糖酶活(Y)为响应值对麸皮浓度(A)、氯化铵浓度(B)、发酵时间(C)进行响应面优化,根据Box-Behnken设计的原理,将数据进行二次多项回归拟合并分析其变化情况,以确定该菌株产酶的最适组合。
2 结果分析
2.1 耐热菌株的筛选
在选择培养基上挑选到4株透明圈与菌落直径比较大的菌株(结果见表2),产酶量与比值呈正相关,可见菌株XY1菌株产酶量较大,对XY1分离纯化并进行后续试验。

表2 透明圈与菌落大小
2.2 木聚糖酶的酶学特性
2.2.1 木聚糖酶的最适温度和最适pH值
图1(a)为酶活力随温度变化趋势图。从图1可以看出,在一定的温度范围内,木聚糖酶的酶活力随着温度的升高而升高,60℃时酶活力达到最大,温度高于60℃后,随着温度的升高,酶活力急剧下降,结果显示,该酶的最适温度为60℃。在55~65℃的温度范围内,相对酶活力(以酶活最高者为100%)达到最适温度酶活的70%以上。不同pH值下的木聚糖酶变化图1(b)所示,数据表明,在pH5.5时,木聚糖酶的酶活最高,该酶最适反应pH为5.5,属于弱酸性水解酶。在pH5-7.0的范围,相对酶活达到最适酶活的65%以上。

图1 温度和pH值对木聚糖酶活力的影响
2.3 产木聚糖酶培养基优化
2.3.1 碳源及用量对酶活的影响
在初始产酶培养基的基础上,替换不同碳源(添加量为1%)(苎麻半纤维参考文献[17]制备)进行试验。由图2(a)可知当碳源为麸皮时酶活力最高,燕麦木聚糖次之,苎麻半纤维素为底物时,木聚糖酶的活力最低。麸皮为制备面粉时的副产物,为重要的农业废弃物,作为发酵培养基时可充分利用农业废弃物生产高附加值的木聚糖酶,因此在后续的发酵研究中选择麸皮为发酵培养基的碳源。以麸皮为碳源改变其添加量进行发酵,图2(b)数据显示,当麸皮添加量为20 g/L时菌株产酶能力最强,增大麸皮的添加量,酶活反而降低。这可能是由于麸皮添加量较高时体系中溶氧过低导致的。
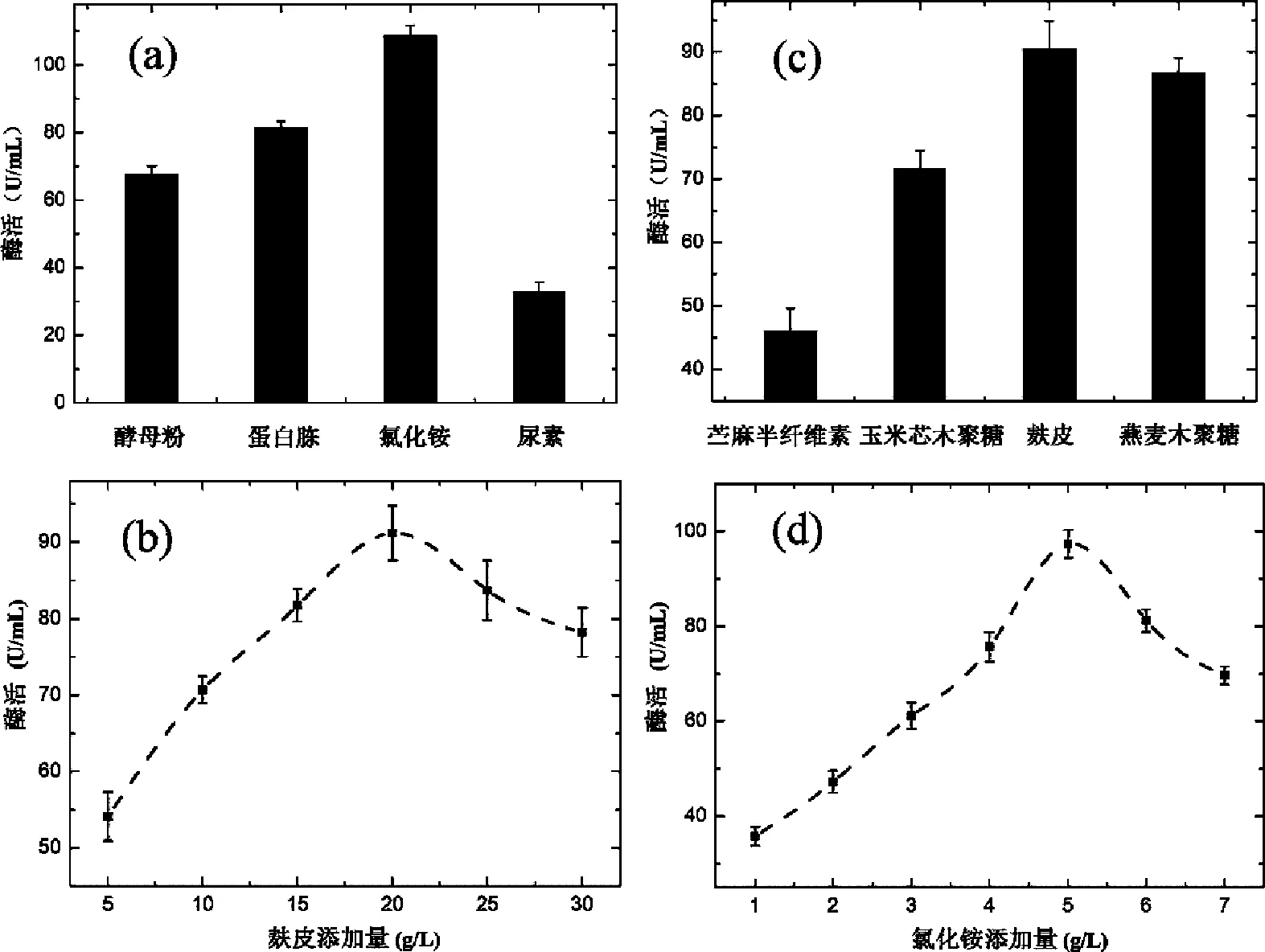
图2 不同碳源和氮源对产酶的影响
2.3.2 氮源及用量对酶活的影响
固定其他因素,替换不同氮源(添加量均为1%),由图2(c)可知氯化铵为氮源时,菌株发酵过程中产酶能力较好,蛋白胨次之。尿素为氮源时,产酶能力最低。可能是由于氯化铵为速效氮源,易于吸收。改变氯化铵添加量为1 g/L、2 g/L、3 g/L、4 g/L、5 g/L、6 g/L、7 g/L分别进行发酵实验,图2(d)结果显示,木聚糖酶的酶活在氯化铵浓度为5 g/L时酶活达到最高值。
2.3.3 发酵时间对酶活的影响
图3结果显示XY1菌株在发酵28h时其酶活达到最高值之后逐渐降低,经spss26.0软件对两组数据分析发现,在24h和28h的酶活数据没有明显差异(p>0.05),故选择24h作为该酶的最适发酵时间。

图3 相对酶活随培养时间的变化
2.4 响应面设计方案与结果
根据单因素试验的结果,令麸皮浓度(A)、氯化铵浓度(B)、发酵时间(C)为自变量,木聚糖酶酶活力(Y)为响应值,以Box-Behnken设计的原理设计响应面,其因素与水平见表3,Box-Behnken设计试验及结果见表4,方差分析结果见表5。

表3 Box-Behnken设计实验因素与水平
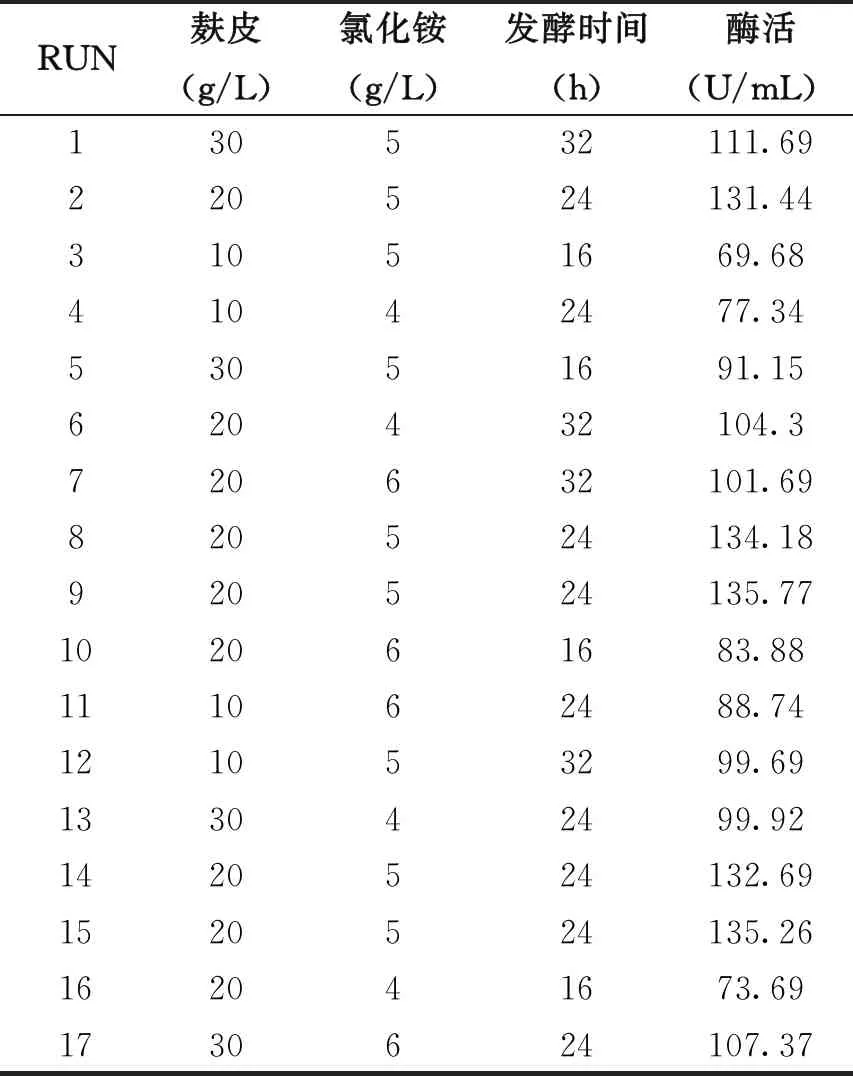
表4 Box-Behnken试验设计与结果

表5 回归模型方差分析
利用软件Design-Expert对实验结果进行回归分析,得出Y木聚糖酶酶活(U/mL)关于A麸皮(g/L)、B氯化铵(g/L)、C发酵时间(h)的回归方程为:
Y=+133.87+9.34A+3.30B+12.37C-0.99AB-2.37AC-3.20BC-19.18A2-21.34B2-21.63C2
根据表5可知p<0.0001,表明该模型极显著;失拟项p = 0.2017>0.05,不显著,说明该回归方程较准确,可以用作发酵产木聚糖酶的优化模型。R2=0.9508,调整R2=0.9897,说明该方程拟合度较高,且该模型可以解释98.97%数据的变化。由于表中BC的p<0.05,而AB、AC的p值大于0.05,说明氯化铵与发酵时间的交互影响较为显著,而麸皮与氯化铵、麸皮与发酵时间的交互影响相对之下不显著。同时表中A2、B2、C2均对响应值产生较为显著的影响(p<0.0001)。
图4是木聚糖酶酶活(Y)对应各因素麸皮(A)、氯化铵(B)、发酵时间(C)形成的三维空间曲面图。令某一因素不变,图8(A)结果显示,在麸皮浓度不变时,酶活随着氯化铵浓度的增大呈现出先增大后减小的趋势,氯化铵浓度不变时,麸皮浓度增大时,酶活也出现先增大后减小的情形。图4(B)与图4(C)表明麸皮与发酵时间、氯化铵与发酵时间的交互影响同样使得酶活出现先增大后减小的趋势。经过Design-Expert优化,分析得到最佳的发酵培养基为:麸皮22.25 g/L,氯化铵5.05 g/L,发酵时间为26.16 h。在此优化条件下所产木聚糖酶酶活理论值为136.68 U/mL。

A麸皮与氯化铵交互作用的等高线图与三维图
2.6 模型验证
为了确定该模型的准确性,在软件给出的优化条件下实验3次,比对分析。测得木聚糖酶酶活的平均值为139.37 U/mL,与理论值误差仅有1.96%,由结果可知此发酵条件能够适用于该菌株发酵产木聚糖酶。
3 讨论与结论

常用于微生物发酵条件优化的方法有单因素法、正交实验、响应面法等。单因素法可筛选出影响发酵的关键因素,但无法反映最佳因素组合,正交实验法可以找出最佳因素水平之间的组合,但难以寻找到因素与组合之间的回归方程,无法考查各因素之间的交互作用。响应面法可以得到高精度的回归方程,在微生物的发酵优化中应用越来越多。Shakir等[23]用响应面法优化提高一株芽孢杆菌产蛋白酶的产量。Vijayaraghavan[24]利用响应面法优化培养基提高了稻黄单胞菌IND3产纤溶酶的活性。
在本研究中,首先采用单因素法确定碳源、氮源、发酵时间等因素,然后利用响应面法确定各因素的最佳值,得到了最佳发酵条件:麸皮22.25 g/L、氯化铵5.05 g/L、发酵时间26.16 h,在此优化的发酵条件下产酶的最大值为139.37 U/mL,比本文中单因素试验最大值提高了29.05%,证明根据响应面法所得的回归方程以及据此优化得到的发酵工艺条件是有效的。
通常情况下,筛选的野生菌株发酵时产酶水平不高,本文通过响应面法优化产酶条件,XY1菌株所产酶活达到139.37 U/mL。粗酶液的酶学性质结果表明,该酶的最适反应温度为60℃,最适反应pH为5.5。该系列结果表明,该菌株产木聚糖酶具有进一步研究的价值和一定工业应用潜力。
猜你喜欢
河南畜牧兽医(2021年9期)2021-12-10 10:43:50
今日农业(2017年11期)2017-01-31 14:53:50
山东工业技术(2016年15期)2016-12-01 05:31:50
中国农资(2016年1期)2016-12-01 05:21:14
中国化肥信息(2016年49期)2016-05-17 04:26:16
中国酿造(2016年12期)2016-03-01 03:08:19
应用海洋学学报(2015年3期)2015-11-22 07:39:30
食品工业科技(2014年15期)2014-03-11 18:17:49
食品工业科技(2014年15期)2014-03-11 18:17:20
中国化肥信息(2014年37期)2014-02-27 13:41:26
