
厌氧铁氨氧化的电子传递机制、环境意义和影响因素
2022-09-07 02:54田莹莹师东阳
昆明理工大学学报(自然科学版) 2022年4期
田莹莹,师东阳,王 强
(1.河南工学院 材料科学与工程学院,河南 新乡 453003;2.河南大学 作物逆境适应与改良国家重点实验室,河南 开封 475004)
0 引 言

(1)
(2)
(3)

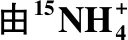
表1 厌氧条件下生成30N2和29N2的潜在反应过程Tab.1 The potential reactions anaerobically to 30N2 and 29N2
1 铁氨氧化过程中的电子传递
异化铁还原菌(FeRB)是Feammox反应中的一类关键微生物,它们具有氧化有机或无机物同时还原Fe(III)成Fe(II)的能力.广泛报道的有地杆菌(Geobacter)、希瓦氏菌(Shewanella)、厌氧粘细菌(Anaeromyxobacter)和酸微菌(Acidimicrobiaceae A6)等[16-17].此外,富集培养实验发现脱硫孢子菌(Desulfosporosinus)、脱氯单胞菌(Dechloromonas)及纤发菌(Geothrix)也对Feammox反应极为重要[5].已报道的微生物Feammox反应胞外电子传递机制主要有以下4种:
1) 微生物与Fe(III)直接接触. 电子传递至微生物外膜表面之后,最直接的是,外膜细胞色素c(c-Cyts)与Fe(III)矿物直接接触.以Shewanella属为例,处于细胞膜最外端的MtrC和OmcA蛋白直接负责了电子向膜外电子受体的传递.缺少MtrC和OmcA,电子传递过程则被阻断[18-19].在Geobacter属中,影响c-Cyts与固态电子受体直接接触传递电子的蛋白是OmcZ,若缺失后,也将阻断电子传递[20](图1(a)).
2) 微生物自身纳米导线. 2005年,Reguera等[21]在Geobacter属中发现一种携带OmcS蛋白的导电纤毛状物质,称其为纳米导线(Nanowire),在S.oneidensisMR-1、蓝细菌等其他菌属中也发现了类似的纳米导线[22].学者们认为通过纳米导线进行胞外呼吸的方式很可能是元素生物地球化学循环中的一种普遍现象,这种方式在微生物与矿物之间起着不可忽视的作用.Geobacter和Shewanella是两类重要的胞外呼吸模式菌,虽都有纳米导线但结构上存在差异.Geobacter产生的纳米导线截面具有很强的导电性,传递微生物胞外电子并延长了呼吸电子传递的距离以代谢远端的矿物[21].Shewanella的纳米导线本质上是细胞外膜与周质向外延伸的一种形式,虽不具电导性,但电子可以在纳米导线表面附着的密集c-Cyts之间跃迁传递,是一种多级电子跃迁的导电机制[23-24](图1(b)).此外,胞外呼吸菌集聚形成生物膜,其具有的纳米导线交织在一起形成纳米导线网络,也能够促进胞外电子的传递(图1(c)).

图1 微生物参与铁氨氧化的电子传递机制Fig.1 Mechanisms of electron transfer in microbes
3) 螯合促溶机制. 一些分子或离子可作为溶铁螯合剂,与Fe(III)氧化物形成可溶性螯合铁,扩散至微生物表面,使Fe(III)被胞外电子还原(图1(d)).螯合剂的存在一方面可增加反应体系中生物可利用Fe(III)的浓度;另一方面还可以提高Fe(III)与铁还原菌直接接触的几率,从而提高铁还原的速率.自然界中除存在多种溶铁螯合物外,有些微生物也能分泌螯合物,如高铁载体(Siderphores),螯合增溶铁.Siderphores含有多种含氧配体,通常对铁离子有很高的亲和力.研究表明,植物根际微生物可通过Siderphores途径促进植物吸收Fe元素[25]。Geothrixfermentans和ShewanellaalgaBrY分泌的Fe(III)螯合物,较大程度上加速Fe(III)氧化物的溶解,进而促进Fe(III)还原[26].某些FeRB分泌胞外聚合物(EPSs),促进还原含镉聚合硫酸铁絮体中的Fe(III),使得絮体解构,释放Fe3+、Fe2+及Cd2+,Fe原子与EPSs中磷酸化蛋白质和核酸中的磷酸基形成P-O-Fe键,固定和聚集溶液中的Fe(III)、Fe(II),促进形成纤铁矿,磁铁矿等铁矿物,依靠铁矿物丰富的羟基官能团,增强了对溶液中Cd2+的吸附[27].螯合铁种类影响Feammox脱氮效能及微生物群落组成,腐殖酸-铁较柠檬酸-铁、乙二胺四乙酸-铁、氨三乙酸-铁总氮去除率最高,达83.2%,优势Comamonadaceae丰度约为17.57%[28].螯合物的存在改变了环境中Fe的存在状态,促进了Fe及其他元素的循环[28],对整个生物圈有重要影响,但螯合物在环境中可能受到其他物质或微生物的影响,使其作用力减弱,对不溶性Fe(III)的增溶减少.
4) 微生物利用电子穿梭体实现电子传递. 环境中大量存在着氧化还原活性的有机物(如可溶态或固态腐殖酸、醌类、抗生素等)都可以作为“电子穿梭体”.微生物氧化呼吸产生的胞外电子首先传递给“电子穿梭体”,然后“电子穿梭体”扩散到含铁矿物表面,并将电子传递给Fe(III),同时还原态的“电子穿梭体”得到氧化(图1(e)).研究发现,某些微生物还能在自身代谢过程中分泌多样的氧化还原活性物质作为电子穿梭体参与胞外电子传递,如Von Canstein 等[29]首次确认Shewanella菌株能够向胞外分泌黄素单核苷酸(FMN)和核黄素促进 MR-1 的厌氧生长和对弱结晶铁氧化物的还原.Wang等[30]认为,微生物产生的FMN可能在其还原Fe(III)氧化物的过程中发挥多重作用:(1)作为MtrC的辅因子促进向铁氧化物(纤铁矿、赤铁矿和针铁矿)传递电子;(2)在还原纤铁矿时作为Mtr路径的电子穿梭体;(3)在还原赤铁矿和针铁矿时作为一种“非反应活性”的电子汇.Okamoto等[31-32]指出 MR-1分泌的黄素分子并非以游离状态发挥作用,而是作为c-Cyts的氧化还原辅因子促进电子传递.黄素与c-Cyts的相互作用促进了通过半醌进行的一电子氧化还原反应,形成结合于MtrC的氧化还原电势为-145 mV的半醌(FMNH),反应速率较游离态黄素的两电子氧化还原反应快103~105倍.除此之外,一电子传递机制对于核黄素同样适用,只不过核黄素分子结合的是外膜上的另一种细胞色素OmcA而非MtrC.Geobactersulfurreducens也能够分泌FMN和核黄素等黄素类分子,且黄素也作为辅因子通过结合于G.sulfurreducens的c-Cyts而参与胞外电子传递过程[33].
上述四种胞外电子传递机制是Feammox反应中重要的微生物金属呼吸方式。目前,有关螯合剂的特性及其与电子穿梭的差异还有待进一步研究.
2 厌氧铁氨氧化的环境学意义
2.1 参与农田土壤氮转化
2.2 影响水生态系统脱氮

2.3 处理废水和污泥

3 铁氨氧化的影响因素
3.1 生物因素
(4)
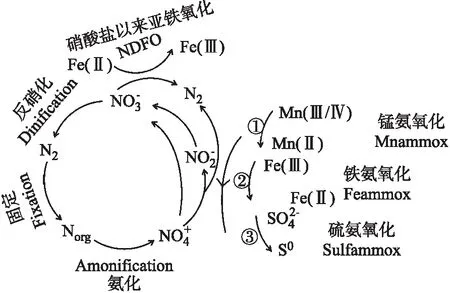
图2 铁氨氧化与其它生物过程的耦合反应[40,47,52] Fig.2 The coupled reactions of Feammox with other biological process
自然环境中元素耦合循环的机制十分复杂.目前,已报道的可能影响Feammox的微生物过程概括如图2.今后基于高通量测序、功能预测和纯培养技术等对环境中微生物进行定性分析、定量分析、功能预测和生理研究等的工作还有待进一步完善.
3.2 非生物因素
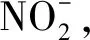
(5)

(6)
(7)
(8)
(9)
4 总结与展望
Feammox是近年来新发现的一种微生物脱氮过程.虽然学者们对Feammox在多种生态系统和污废水处理体系中的重要作用等方面已积累了一定的研究基础,但已有研究主要关注的是Feammox菌群在特定环境下的分布情况,对Feammox菌群与环境中其他功能微生物物种间的潜在相关关系了解甚少。今后,要基于宏基因组测序技术和生物信息学软件深度挖掘特定生态系统或体系中的关键微生物,并将关键微生物与其执行的生态功能(如氨氧化、碳固定、反硝化等)同分析。对关键微生物的生理特性研究也不容忽视。采用传统的微生物分离和纯化技术,获得Feammox纯菌株,通过室内模拟实验,完善纯菌生理和生化特性,深入探究纯培养和混合培养条件下Feammox菌的脱N能力、发生机制和各成员间的生态关系等对生态环境学科发展具有推动作用。此外,自然环境中Fe的氧化还原往往可与生境内的本身存在多种其它元素(如Mn、P、S、C等)存在耦合,可能促进或抑制Feammox活性,因此,查明复杂环境中Feammox的反应效能和作用机制也将是一项重要课题.最后,Feammox作为新近发现的一种微生物过程其机理和应用研究方兴未艾.
猜你喜欢
肥料与健康(2022年3期)2022-12-04
科技创新与应用(2022年12期)2022-05-08
山西医科大学学报(2021年10期)2021-11-18
天津大学学报(自然科学与工程技术版)(2021年9期)2021-06-01
汽车电器(2021年4期)2021-05-07
纺织科学研究(2021年1期)2021-03-19
农业与技术(2021年1期)2021-01-18
汽车电器(2020年2期)2020-03-13
海峡姐妹(2019年8期)2019-09-03
视野(2016年6期)2016-05-14
