
幼龄胡杨气孔行为对土壤质地和地下水埋深变化的响应
2022-08-31 06:08何汐然丁晓雪许毓哲
生态学报 2022年15期
何汐然, 丁晓雪, 许毓哲, 李 君
1 中国科学院大学生态与地理研究所,荒漠与绿洲国家重点实验室,乌鲁木齐 830011 2 中国科学院大学, 北京 100049 3 中国科学院阿克苏绿洲农田生态系统国家野外科学观测研究站,阿克苏 843017
气孔控制植物叶片水分损失和二氧化碳吸收,在植物适应环境变化中起关键作用[1—2]。植物叶片气孔周围保卫细胞与叶表皮之间膨压差控制气孔开闭,而保卫细胞膨压变化非常快,因此气孔开度能够快速改变以适应生境条件变化[3]。这种快速调节能力对于植物在多变生境条件下优化水碳关系至关重要[3—4]。最优气孔行为理论认为气孔最优化行为是在控制一定水分消耗的同时,使碳吸收最大化[5]。水分利用效率反映了水分消耗与碳吸收之间的关系,表征了单位耗水的固碳量。气孔导度斜率是气孔模型中的重要参数,能有效反映气孔对CO2浓度、水汽压亏缺等环境要素变化的敏感性。在气孔机理模型中,气孔导度斜率通常与水分利用效率呈一定比例关系,可用来描述植物水分利用策略[6]。研究表明,干旱时植物的气孔导度斜率降低,水分利用效率升高[7],且干旱时,湿润区植物气孔导度斜率降低,而半湿润区植物气孔导度斜率保持稳定[8]。因此,对比不同生境下植物气孔行为可反映植物对生境条件变化的适应策略。
胡杨(Pouluseuphratica)作为塔里木河流域荒漠河岸林建群种[9—10],在维护区域生态安全方面起着重要作用[11—12]。洪水漫溢后形成的湿润、裸露河漫滩是胡杨种群的自然更新生境[13—14]。由于水文波动和河流沉积作用影响,河漫滩生境地下水波动频繁,土壤质地时空异质性高[15],这些因素将制约胡杨幼苗在河漫滩的生长发育。幼龄期胡杨抗干扰能力低、死亡率高,生活史早期存活率是限制种群天然更新的关键过程[16—17]。研究表明,地下水埋深变化会影响植物生物量分配格局[18—19]、响应机制及适应策略[20—21]。刘深思等研究发现,沙柳气孔导度随地下水埋深加深而降低[22]。王鹏等研究发现不同矿化度处理下,地下水埋深越浅多枝柽柳幼苗碳同化能力越强[23]。这些研究多集中于探讨地下水埋深变化对于植物生理行为及适应方式的影响[24],而对于土壤质地及其与地下水埋深的交互作用对植物生理行为的影响报道较少。因此,本研究以幼龄胡杨为研究对象,通过简易蒸渗仪控制土壤质地和地下水埋深,研究不同土壤质地、地下水埋深及两者的交互作用对幼龄胡杨气孔行为的影响,分析相关气孔参数之间的关系,以阐明胡杨在适应河漫滩多变生境的气孔行为的调控作用,为胡杨林更新和保护提供重要的科学依据。
1 研究区概况
该研究区位于中国科学院阿克苏水平衡实验站,地理坐标为40°27′N、80°45′E。研究点所在区域属暖温带干旱气候,年平均气温为10.6—11.5 ℃,年平均降水量为45.7 mm,年平均潜在蒸发量为2000—3000 mm,全年无霜期达207 d,日照时数达3000 h左右。研究区域原生植被为荒漠河岸植被,主要为杨柳科、柽柳科、豆科等,优势种为胡杨、柽柳(Tamarixspp.),其他常见种主要为花花柴(Kareliniacaspica)、疏叶骆驼刺(Alhagisparsifolia)、铃铛刺(Halimodendronhalodendron)等。
2 研究方法
2.1 试验材料与设计
试验开始于2020年5月20日,历时92 d。供试材料是两年生幼龄胡杨,其长势均一,移植时株高(40±5) cm。栽培装置为简易蒸渗仪,蒸渗仪口径20 cm、高120 cm,每个蒸渗仪移栽2株。试验设计为地下水埋深和土壤质地交互试验,各因子3个处理,地下水埋深为30 cm(W1),60 cm(W2)、90 cm(W3),土壤质地为砂土(S1)、砂壤土(S2)、黏壤土(S3),供试土质理化性质见表1。供试土壤经过筛后按容重装填至蒸渗仪。不同地下水埋深、土壤质地组合共计9个处理,每个处理4个重复,共计36个蒸渗仪。幼龄胡杨移栽一个月后待其正常生长后,开始对其进行水分控制处理。开始试验后,每天20:00向控制水位柱加水以保证试验期间地下水埋深稳定在控制水平。
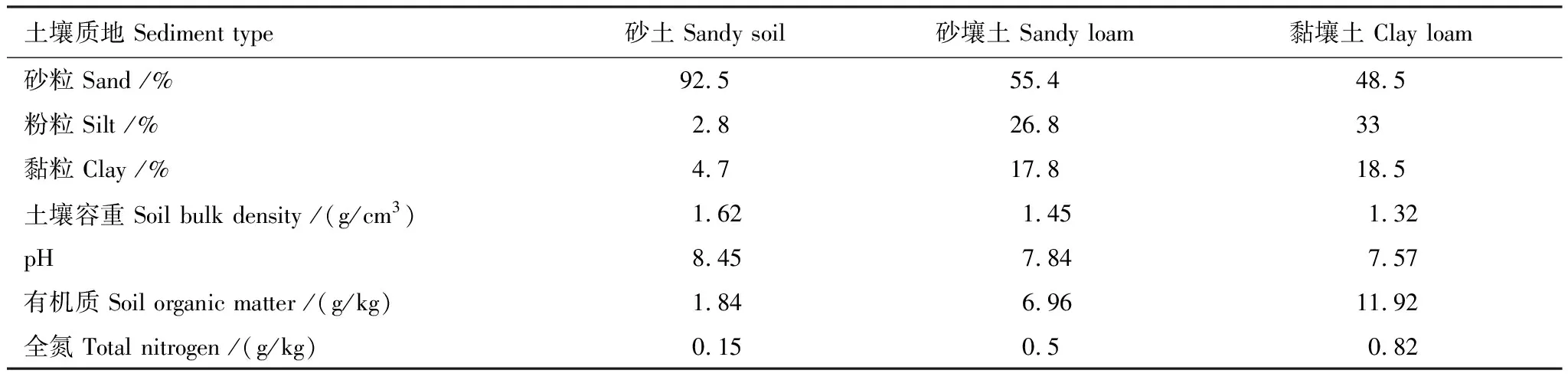
表1 供试土壤的理化性质
2.2 参数测定
利用植物压力室PMS(PMS Instrument Co.,Albany, OR, USA)对不同处理条件的胡杨枝条的凌晨和正午水势进行测定。测量时每一处理条件3个重复,取其平均值记为水势值。凌晨、正午水势的测定时间为05:30—07:30和13:30—15:30。
2020年8月,采用LI- 6400便携式光合测量系统(LI-COR Corp., Lincoln, NE, USA)测量不同处理下胡杨光合日变化过程。光合日变化过程观测选择晴朗无云天气下进行。每一处理随机选取3株,每株选取四片向阳健康的叶片,在8:00—20:00每隔2 h测定一次光合参数。测定的参数包括Gs、净光合速率(Pn)、蒸腾速率(Tr)、饱和水汽压亏缺值(D)、环境二氧化碳浓度(Ca)、胞间二氧化碳浓度(Ci)等光合参数。测定完扫描计算叶室内叶片面积,对光合参数进行重计算。
2.3 数据处理
Medlyn等人提出的气孔导度机理模型[7],结合了collatz等人的BBL模型[25]、Santrackova等人的光合模型[26]和Cowan等人的最优气孔控制模型[5],能综合反映环境二氧化碳浓度(Ca)、饱和水汽压亏缺(D)及净光合速率(Pn)与气孔导度的关系。为明确不同处理条件下胡杨气孔导度斜率变化,采用气孔导度机理模型对气孔导度进行拟合[7]:
(1)
式中,gs为气孔导度(mol m-2s-1),g0为零光合时的气孔导度;A为光合净CO2吸收速率(μmol m-2s-1),即所测Pn;Ca为环境二氧化碳浓度,D为饱和水汽压亏缺;g1为气孔导度斜率,能有效反映气孔导度对CO2浓度、饱和水汽压亏缺和光合作用的敏感性[4],且与临界水分利用效率(λ= ∂A/∂E,mmol CO2/mol H2O)成一定比例关系[3](方程(2)),可描述植物水分利用策略。
(2)
式中, Г为CO2补偿点(单位:μL/L),λ为临界水分利用效率(单位:mmol CO2/mol H2O)。
为明确不同处理条件下气孔行为对叶片光合限制的差异,计算了气孔限制值(Ls),Ls的计算采用Berry和Downtow提出的公式(方程(3))[27]:
Ls=(Ca-Ci)/(Ca-Γ)
(3)
式中,Г为CO2补偿点,常忽略不计,公式可改写为:
(4)
光合的非气孔限制值由Ci/Gs确定[28]。
水分利用效率(WUE)按照如下公式计算:
(5)
式中,Pn为净光合速率(μmol m-2s-1),Tr为蒸腾速率(mmol m-2s-1)。
采用双因素方差分析(Two-Way ANOVA)检验土壤质地、地下水埋深及二者的交互效应对胡杨气孔导度、气孔限制值、水分利用效率等指标的影响。数据统计分析软件采用SPSS 20.0(SPSS for Windows, Version 20.0, chicago, IL, USA)。
3 结果
3.1 不同处理条件下胡杨生理参数的差异
如表2所示,土壤质地对所测定的胡杨指标参数均有显著影响(P<0.05);地下水埋深对胡杨气孔限制值与正午水势均有显著影响(P<0.05);土壤质地和地下水埋深的交互作用仅对非气孔限值具显著影响(P<0.05)。总之,土壤质地是引起胡杨气孔行为差异的主效应因素,而地下水埋深及两者交互作用对胡杨气孔行为无明显影响。
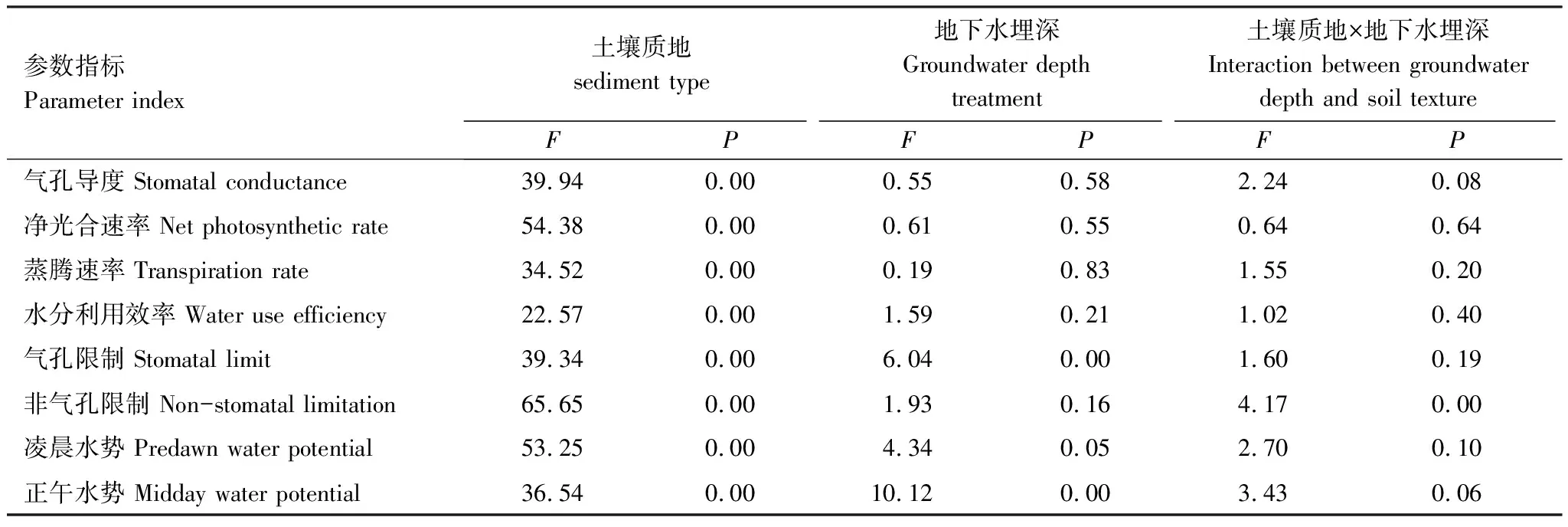
表2 土壤质地及地下水埋深对胡杨各参数指标的方差分析结果
3.2 不同处理条件下的胡杨枝条水势变化
由图1可以看出,在三种地下水埋深条件下,S1中胡杨凌晨水势最低,S1W1为-2.54 Mpa,S1W2为-2.26 Mpa,S1W3为-2.65 Mpa,且显著低于S2与S3;而S1中胡杨正午水势显著低于S3;以上结果表明S1水分条件差,胡杨水分亏缺严重。其次,植物枝条水势对于地下水埋深的响应因土壤质地的不同有所差异,在S2与S3中胡杨凌晨与正午水势随着地下水埋深的增加而升高;这说明随着地下水埋深加深,胡杨水分亏缺程度有所缓解;而S1中胡杨水势变化不具该规律,在S1W3时具最小凌晨水势与正午水势 (-3.64 Mpa),S1W2具最小凌晨水势与正午水势(-3.22)。

图1 不同处理条件胡杨凌晨水势与正午水势变化Fig.1 The changes of predawn and midday water potential in xylem of Populus euphratica under different treatment conditionsW1: 地下水埋深30 cm Groundwater depth 30 cm;W2: 地下水埋深60 cm Groundwater depth 60 cm;W3: 地下水埋深90 cm Groundwater depth 90 cm;S1:砂土 sandy soil;S2:砂壤土sandy loam;S3:黏壤土 clay loam;不同小写字母代表同一地下水埋深条件下不同土质间存在显著差异(P<0.05)
3.3 不同处理条件下的气孔行为变化
3.3.1不同处理条件下的气孔导度变化
由图2可知,Gs对地下水埋深的响应因土质不同具有一定差异。W1条件下,S1中胡杨Gs显著低于S2与S3;W2条件下,S3中胡杨Gs显著高于S1与S2;W3条件下,S3中胡杨Gs显著高于S2,S2显著高于S1,这表明胡杨气孔开度在不同地下水埋深下的表现与土壤质地状况相关。同一土质中胡杨Gs对地下水埋深变化的响应为:S1中胡杨在W2时具最大Gs(0.15 mol m-2s-1),W3时最小Gs(0.09 mol m-2s-1),两者相比,Gs降低40%;S2中胡杨在W1时具最大Gs(0.23 mol m-2s-1),W2时最小Gs(0.17 mol m-2s-1),两者相比,Gs降低26.1%;S3中胡杨Gs随着地下水埋深加深而增大。以上结果表明,胡杨气孔开度对地下水埋深变化的响应因土壤质地具有一定差异。
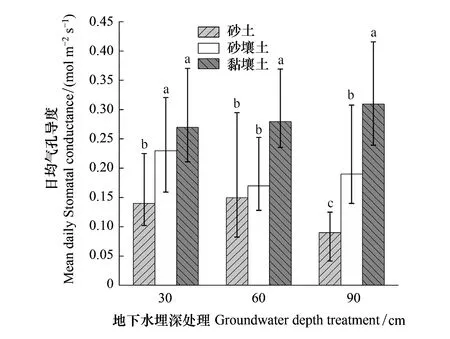
图2 不同处理条件对胡杨气孔导度的变化 Fig.2 The changes of stomatal conductance of Populus euphratica under different treatment conditions
3.3.2不同处理条件下的气孔导度斜率变化
在不同处理条件下,胡杨幼苗具有不同的气孔导度模型参数。由气孔优化模型拟合线性回归分析结果可知,除S1W3条件下胡杨幼苗气孔导度变化不符合气孔导度模型外(P>0.05),其他处理条件下的拟合结果较好(P<0.05)。在W1与W2条件下,S1中胡杨g1最大,S3次之,S2最小;W3条件下,胡杨g1大小依次为S3最大,S2次之,S1最小。而相同土壤质地中的胡杨g1对地下水埋深变化的响应规律表现为:随着地下水埋深加深,S1中胡杨g1无明显变化;S2中g1呈先降低后升高趋势,在W1时出现最大值,为2.001,W2时出现最小值,为1.49;S3中g1呈下降趋势。上述结果说明,胡杨g1对地下水位的响应因土壤质地的差异而表现不同的规律(图3)。
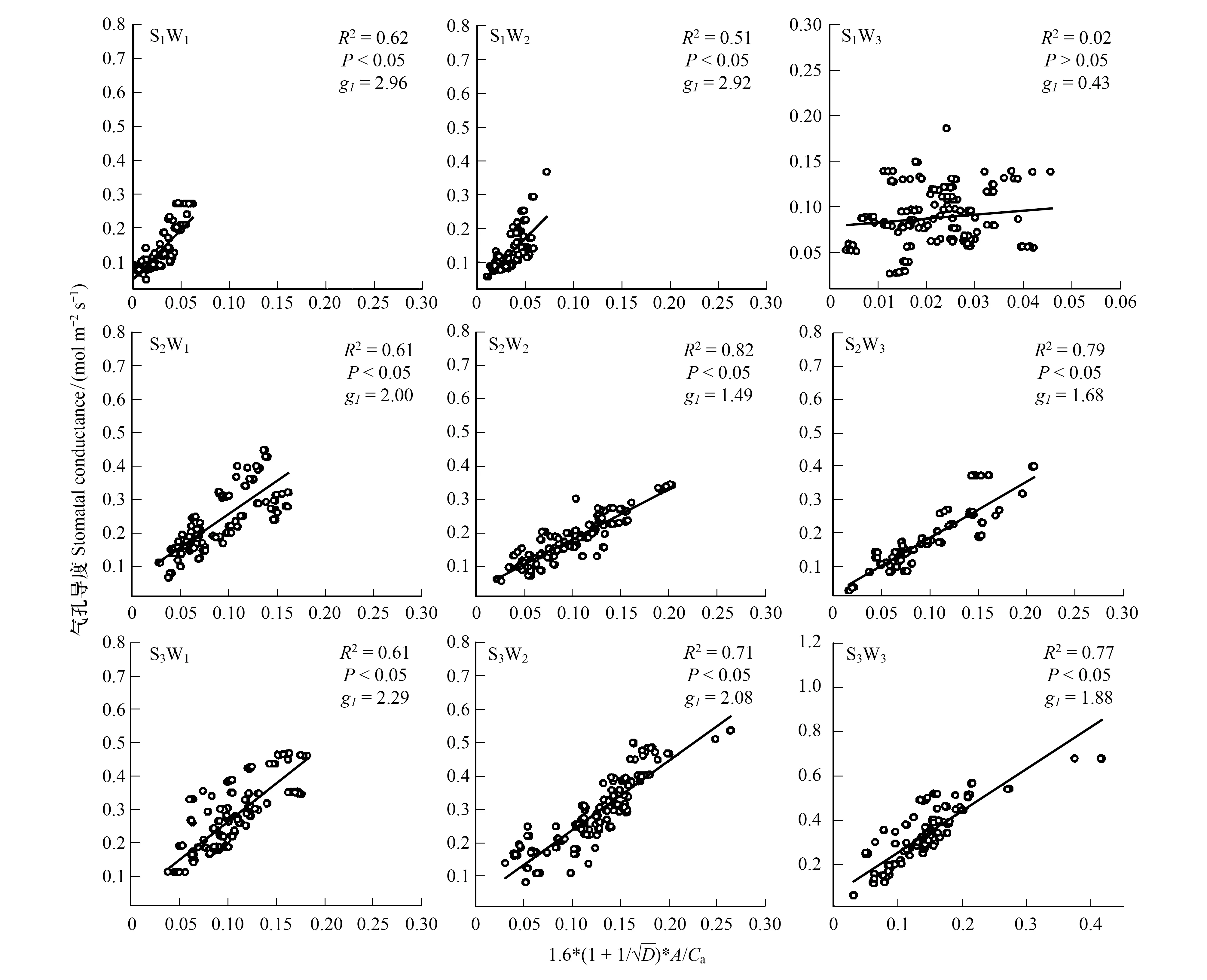
图3 不同处理条件下胡杨气孔导度与环境因子及光合参数的关系Fig.3 Relationships between stomatal conductance and the environmental and photosynthetic factors for the Populus euphratica under different treatment conditions光合、环境综合因子 Photosynthetic and environmental factors;D:水汽压亏缺 Vapor pressure deficit;A: 净光合速率 Net photosynthetic rate;Ca:环境二氧化碳浓度 Ambient carbon dioxide concentration;g1:气孔导度斜率 stomatal conductance slope
由图4可知,W1条件下,S2与S3中胡杨WUE显著高于S1,这说明这两种土质中的胡杨维持较高水平的WUE;W2与W3条件下,WUE则表现为S2显著高于S3,S3显著高于S1,即随着地下水埋深加深,S2中的胡杨提高了单位耗水碳生产量。且由分析可知,气孔导度斜率与水分利用效率呈显著负相关,当气孔导度斜率减小时,水分利用效率呈增大趋势。

图4 不同处理条件胡杨气孔导度斜率与水分利用效率的变化Fig.4 The changes of stomatal conductance slope and water use efficiency of Populus euphratica under different treatment conditions
3.3.3不同处理条件下气孔对光合作用的限制
不同处理条件下,胡杨气孔、非气孔因素对于光合的限制作用差异很大。由图5可知,相同地下水埋深处理条件下,S2与S3中胡杨Pn显著高于S1,且S1中胡杨非气孔因素对光合作用的限制显著高于S2与S3。而胡杨气孔因素对光合的限制作用变化与非气孔限制有所差异,W1条件下,S2与S3中胡杨气孔对于光合作用的限制显著大于S1;W2与W3条件下,S2中胡杨气孔对光合作用的限制显著高于S3,S3显著高于S1。随着地下水埋深变化,三种土壤质地中胡杨气孔对于光合的限制中呈相同的趋势,即在W2与W3时,胡杨气孔对光合的限制处于同一水平,但高于W1时的限制作用;而非气孔因素对光合的限制在三种土质中表现具有差异,主要体现在S1中非气孔因素对光合的限制作用在W3时达到最大(3.61),S2与S3中胡杨非气孔因素对光合的限制作用随着地下水埋深的加深而减弱。
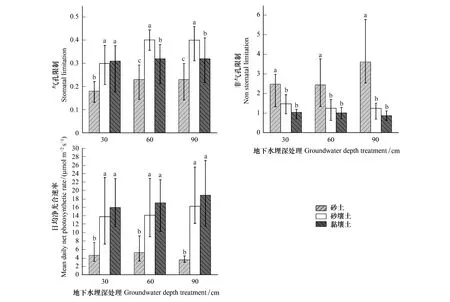
图5 不同处理条件胡杨气孔、非气孔对光合作用的影响Fig.5 The effects of stomatal and non-stomatal limitations of Populus euphratica under different treatment conditions
4 讨论
凌晨和午间水势反映了植物水分的亏缺程度。本试验中,相同地下水埋深条件下,S1中胡杨枝条水势显著低于S2与S3,表明S1条件下胡杨受到较高程度的水分亏缺。何建新[29]等人的研究表明,0.05—0.005 mm颗粒含量是影响毛细水上升高度的重要因素,且该粒径范围的颗粒含量越高,毛细水上升高度越高。而本研究砂壤土与黏壤土中粉粒(0.002—0.05 mm)含量显著高于砂土,相应的毛细水上升高度升高,更利于植物水分吸收,这可能是不同土质引起胡杨水势差异的原因。已有研究表明,不同地下水埋深将会影响植物水势[30—31],而本研究中S2和S3的胡杨凌晨与正午水势随着地下水位降低而增大。朱文东的研究发现,当地下水埋深越浅,表层土壤盐分积累越明显[32]。而本试验区域位于塔里木河上游干流地区,土壤盐渍化较为严重[33],地下水位浅导致上层土壤盐分积累,降低土壤水分的有效性,这可能是地下水埋深变化引起胡杨水势差异的原因。
气孔开度影响植物水分利用与碳吸收,对植物生产力与抗旱性具重要作用[34]。裴艳武等的研究表明,同一水分处理下,长柄扁桃不同时期气孔导度在不同质地土壤间均表现出显著差异,但这种差异在不同水分条件下表现不同[35]。而本研究发现,在同一地下水埋深处理下,胡杨幼苗气孔导度在黏壤土中最大,砂壤土中次之,砂土中最小,但不同地下水埋深处理下,不同土质间气孔导度差异性表现不同。可能原因是土质中砂粒含量减少,有利于植物不定根生长[36—37],而地下水埋深的加深,植物根系垂直分布越分散,两者的共同作用将有助于植物吸取更深层水分[38],从而影响植物水分状况,改变气孔开度。
气孔导度斜率会对外界生境条件的变化做出不同的响应。在不同水分生境下,同一物种具有不同的气孔导度斜率[39]。本研究中,由于土壤质地的差异,胡杨面临不同的水分条件,气孔导度斜率相应做出不同的响应。前人的研究表明,植物受到土壤水分胁迫时,对气孔导度的模拟可以通过改变干旱胁迫与光合作用/气孔导度之间作用关系来达到的[40—41],也可通过干旱胁迫与光合作用或者光合作用参数关系的变化,模拟光合作用的变化进而模拟气孔导度的变化[42—43]。但在植物受到严重干旱胁迫时,上述方法无法准确模拟光合作用和蒸腾作用[44—45]。本试验中,在对气孔导度的模拟发现,砂土中胡杨在地下水埋深90 cm时拟合结果差,可能原因是该条件下胡杨受到严重水分胁迫,不能准确模拟胡杨气孔导度变化,无法准确得到气孔导度斜率。而其余地下水埋深处理下,砂土中胡杨气孔导度斜率最大,且WUE最低。这可能表明在严重水分亏缺条件下,气孔的调控不符合优化气孔理论,气孔的变化仅为植物生存,并非为生长。Somerville等人[46]的研究表明在同样的生境下,气孔导度斜率低的植物具有较低的气孔导度,而小幅度降低气孔导度可提高水分利用效率。本研究的砂壤土中胡杨气孔导度斜率最小,而相较于地下水埋深30cm处理, 60与90 cm处理的气孔导度小幅降低,但水分利用效率却显著提高,此试验结果与前人研究相符。S3中胡杨幼苗Gs随着气孔导度斜率减小而增大。这可能是黏壤土水分充足,植物通过保持较高气孔导度加大水分消耗维持较高水平碳同化能力。气孔导度斜率与水分利用效率存在显著的线性关系,随着气孔导度斜率减小,水分利用效率增大,这与Lin等人[7]的研究结果相符,与Héroult等人[8]提出的理论公式相一致。
植物生长过程中,水分亏缺会对植物光合产生气孔限制与非气孔限,导致植物的光合作用下降,影响植物正常生长[47]。本研究中,不论地下水埋深变化,S1中胡杨日均光合作用显著低于S2与S3。已有研究表明[48—49],水分亏缺严重条件下,非气孔限制对光合作用占主导,且非气孔限制通常比气孔限制对光合速率的影响更大[40],与本实验结果相符合,即S1中胡杨水分亏缺严重,非气孔限制占主导,显著降低胡杨光合作用,且S1W3条件下非气孔限制最大,光合作用最小。S2与S3中胡杨日均光合作用随着地下水埋深加深逐渐增大。两种土质中,随着地下水埋深加深胡杨气孔限制作用有所加大,但非气孔限制作用减弱,而非气孔限制作用的影响更大[39],这可能是引起光合作用增强的原因。
5 结论
目前已有的研究主要集中于地下水埋深变化对植物气孔行为的影响,但忽略了植物在土壤质地变化时的响应差异。本研究发现,不同土壤质地中的胡杨气孔行为对地下水埋深变化的响应具有差异性。砂土中幼龄胡杨通过保持低Gs与高g1适应地下水埋深变化,光合作用受到较大的非气孔限制作用,导致胡杨在保持低水分消耗的同时WUE也保持在低水平。砂壤土中幼龄胡杨通过降低Gs和保持低g1适应地下水埋深变化,随着地下水埋深加深,胡杨在降低水分消耗的同时提高WUE,维持高效碳同化水平。黏壤土中幼龄胡杨通过保持高Gs和较低g1适应地下水埋深变化,随着地下水埋深加深,胡杨在增大水分消耗的同时具较高WUE,维持高碳同化能力。本文研究证明土壤质地改变了胡杨气孔行为对地下水埋深变化的响应,表明不仅仅是生境的水分条件,土壤质地也是影响胡杨生活史早期植物水分关系的重要因子。研究的结果丰富了我们对荒漠河岸带植物水分关系的认识,明确了土壤质地如何通过与地下水交互作用而限制或促进幼龄胡杨的生长,因而对深入认识河岸带幼龄胡杨空间分布以及胡杨自然更新的环境约束具有重要实践意义。
猜你喜欢
中国农业科学(2022年15期)2022-08-09
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2021年6期)2021-12-08
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
安全与环境工程(2021年2期)2021-04-02
农村实用技术(2020年9期)2020-11-19
节水灌溉(2020年9期)2020-09-16
湖北林业科技(2019年4期)2019-09-16
天津农业科学(2015年8期)2015-08-06
