
刺五加群落多样性海拔梯度变化及相似性
2022-08-28 02:35高金辉韩家永张厚良林国英张莹艾志强刘继云
森林工程 2022年4期
高金辉,韩家永,张厚良,林国英,张莹,艾志强,刘继云
(黑龙江省林业科学院伊春分院,黑龙江 伊春 153000)
0 引言
刺五加(Acanthopanaxsenticosus)为五加科(Araliaceae)五加属(Eleutherococcus)多年生落叶小灌木,主要分布于我国东北三省、河北省和山西省,是针阔叶混交林下的重要组成树种。刺五加全株皆可入药,具有益气健脾、补肾安神的作用,药效可媲美人参(Panaxginseng),已被收录于《中国药典》[1]。然而因其优良的药用功能和疗效,资源消耗与日俱增,也因其具有较高经济价值而带来大量不合理采挖活动,野生资源遭到严重破坏,使其变成渐危物种[2]。此外,在天然条件下,刺五加种子萌发的实生苗很少,现存大量刺五加种群多为无性植株组成,这样容易使基因多样性减少,导致出现绝种风险。为了保护刺五加适生环境和恢复野生资源,从群落生态学角度研究群落多样性和演替梯度变化,诠释最优群落分布格局、结构和稳定程度,具有重要现实意义。
植物群落物种多样性研究可以反映植物群落生长环境差异、植物群落结构差异以及群落物种丰富度和均匀度,其随环境梯度的变化现已成为国内外群落生态学家争相研究的热点[3-8]。海拔梯度变化直接改变着山地生态环境的各项因素,如水分、温度、湿度和土壤结构等[9],很多研究发现海拔与群落多样性呈负相关或单峰型关系[10-12]。刺五加所生长森林群落的典型特征是其空间异质性明显、水热等环境因素差异较大,导致群落中林木的空间分布、物种组成和群落生长发育均呈现动态变化[13],故探索其群落多样性变化和群落之间相似性随海拔梯度的变化规律十分必要。近年来,关于刺五加的研究大部分集中在化学成分分析、药理作用和育种栽培等领域[14-16],基因、生物多样性和遗传结构等方面也有一些探索[17-20],但刺五加群落多样性与海拔之间关系的研究鲜见报道。
鉴于此,本研究以黑龙江省小兴安岭南坡山地分布的典型刺五加群落为研究对象,对不同海拔刺五加群落的物种组成进行样地调查,探究刺五加群落物种多样性与海拔梯度之间的变化关系,并分析海拔因素对群落物种多样性的影响,旨在深入探讨刺五加群落物种多样性的垂直分布格局,为刺五加野生资源的合理利用和群落物种多样性保护提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
小兴安岭位于黑龙江省北部,西北以五大连池-黑河一线与大兴安岭相接,东南达松花江谷地,东北以黑龙江为界,东接三江平原,西以小兴安-铁力-巴彦为界,与松嫩平原相邻,地势东南高、西北低,地貌表现出明显成层性,属低山丘陵地形。小兴安岭具有得天独厚的自然生态条件,年均气温-2~2 ℃,年均降雨量500 mm,无霜期100 d左右。繁衍生长着红松(Pinuskoraiensis)、落叶松(Larixgmelinii)、樟子松(Pinussylvestrisvar.mongolica)和“三大硬阔”[胡桃楸(Juglansmandshurica)、水曲柳(Fraxinusmandshurica)、黄檗(Phellodendronamurense)]等许多珍贵树木。植被类型为典型温带针阔混交林,生物资源相对贫乏,生物多样性程度相对较低。
1.2 样地设置和调查方法
2020年5—9月,对小兴安岭南坡刺五加典型群落进行野外调查,采用样地法,沿海拔梯度利用 GPS 定位进行样地布设,尽量控制海拔以外的其他环境因素。样地调查区分别为嘉荫(海拔93 m)、双丰(海拔300 m)、五营(海拔447 m)、伊美(海拔469 m)、新青(海拔799 m)、桃山(海拔1 055 m),每个调查区设置3块样地,6个海拔梯度共设置18块样地。每块样地设置一个面积20 m×20 m样方,在样方内按梅花点布设5个5 m×5 m灌木样方,再在每个灌木样方内选取5个1 m×1 m草本样方,共设置18个样方、90个灌木样方和450个草本样方。调查内容包括样地经纬度、海拔和坡度等基本概况以及乔木、灌木、草本植物的种数、株高、地径、冠幅和株数等。研究样地描述见表1。
1.3 群落物种多样性和群落结构测度方法
采用Microsoft Office Excel 2003、SPSS 25和Forstat 3.0软件进行数据记录、处理和数据统计分析。
1.3.1 群落α多样性
Simpson多样性指数:
Shannon多样性指数:
Margalef指数:
d1=(S-1)/lnn。
Menhiniek指数:
式中:S为研究样本中的物种总数;pj为某一物种出现的频率;n为观察到的个体总数。
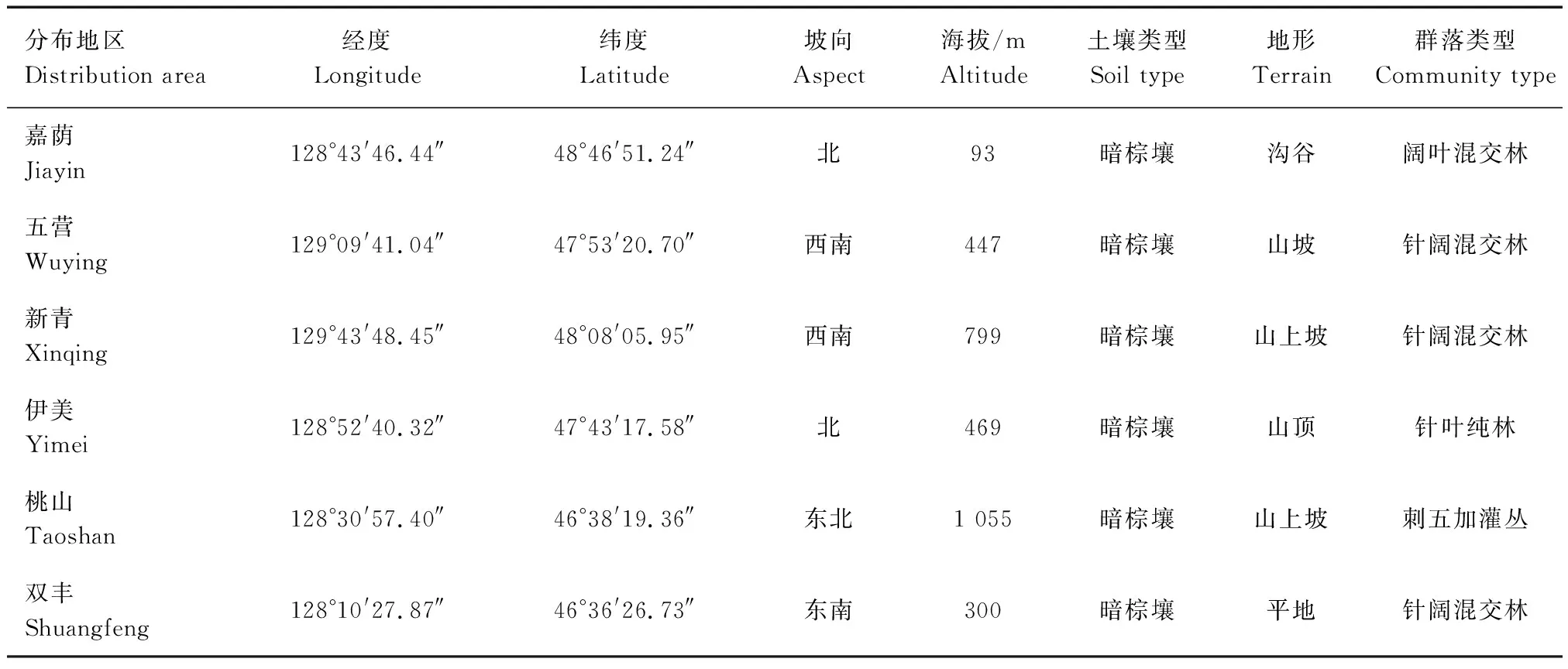
表1 研究样地描述Tab.1 Description of the sample plots
Margalef指数和Menhiniek指数均是从群落的物种数量和个体总数方面描述种的丰富度,但2种指数对其丰富度反映的敏感性不同。
Simpson均匀度指数:
Shannon均匀度指数:
Jh=H/lnS∈(0,1]。
均匀度指数是指群落的实测多样性与最大多样性的比值。两种均度指数分别用Simpson多样性指数和Shannon多样性指数计算,前者得出群落中随机选取两个个体属于同一个物种的概率高低,后者描述物种数目及物种中个体分配的均匀性。
优势度指数
式中:nj为第j种的个体数。
1.3.2 群落β多样性
Cody指数:
Sorenson指数:
式中:a为2个相邻海拔梯度样地内共有的物种数;b为沿海拔梯度变化而消失的物种数;c为沿海拔梯度变化而增加的物种数。
1.3.3 群落相似性
采用Jaccard相似性公式计算刺五加群落相似性系数
式中:G为2块样地之间共有的物种数;A、B为某一群落的物种数。
2 结果与分析
2.1 刺五加群落结构特征
刺五加群落结构数据见表2。
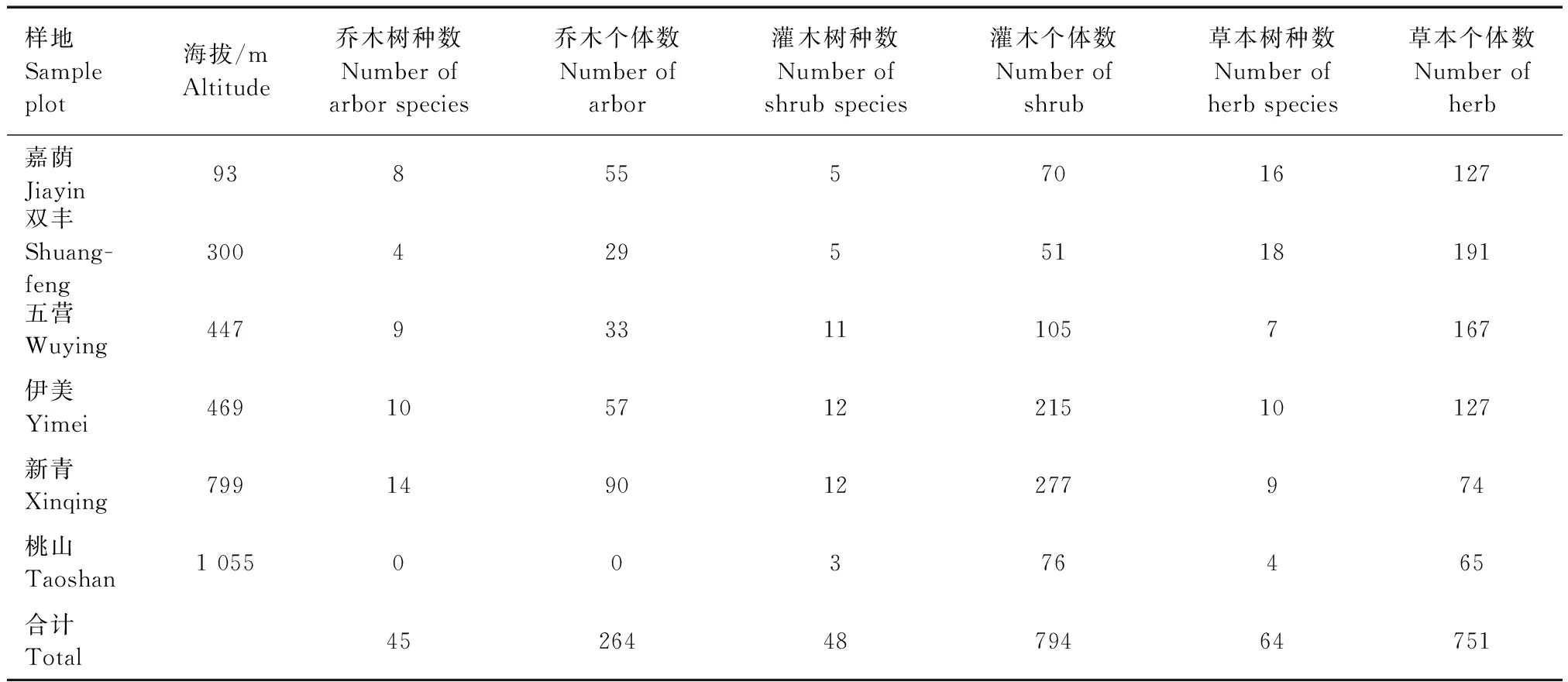
表2 研究样地物种统计
刺五加群落中,随着海拔升高,乔木、灌木树种数呈先增高后降低趋势,海拔799 m的新青样地乔木树种数最多,达14种;海拔469 m的伊美样地和海拔799 m的新青样地灌木树种种类最多,均达12种,这与新青样地特殊的地形条件有关,人为干扰极少。乔木和灌木树种个体数随着海拔升高均呈先降低再升高后降低趋势,与物种数的变化趋势基本相似。草本种类和数量变化趋势与乔木和灌木不同,随着海拔升高大致呈降低趋势,海拔300 m的双丰样地草本种类和数量最多,达18种191株,主要原因可能是较少的乔木和灌木种类和数量导致林分郁闭度较低,为林下草本提供了充足的光照和营养空间。桃山样地位于小兴安岭最高峰平顶山上,刺五加分布在全光条件下,群落结构只有灌木和草本,特殊的地形和气候条件导致桃山样地各生长型种类和数量均有限,适生物种较少,未表现出一定规律性。
综上所述,随着海拔升高,刺五加群落中乔木、灌木和草本树种物种组成变化不明显,只有海拔达1 000 m以上,物种组成变化较大,原有物种几乎全部消失,随之出现蹄叶橐吾(Ligulariafischeri)、小花风毛菊(Saussureaparviflora) 和北乌头(Aconitumkusnezoffii)。刺五加种群作为群落的优势种群,除海拔300 m和1 055 m无花楷槭(Acerukurunduense)、海拔93 m和1 055 m无东北茶藨子(Ribesmandshuricum),其他海拔梯度上均有较大比例分布,作为群落中的次优势种存在。海拔、立地条件和生境条件等小空间尺度因子对群落类型影响作用明显。
2.2 刺五加群落生物多样性的垂直变化
2.2.1 群落α多样性沿海拔梯度的垂直变化
小兴安岭植物区系属北部边缘类型,发生在温带针阔叶混交林植被所要求的最低热量的极限条件下,大多数物种基本达天然分布的最北界,物种丰富度和多样性相对较低。植物群落物种组成决定群落多样性特征,也反映物种种群的适生环境。群落α多样性各指数从不同角度表征植物群落内的物种组成情况,表3为不同海拔刺五加群落乔木层、灌木层、草本层和群落α多样性各指数。

表3 刺五加群落α多样性指数
乔木层α多样性结果显示,除双丰(海拔300 m)和桃山(海拔1 055 m)外,其他各样地的各指数沿海拔梯度没有变化。灌木层α多样性结果显示,各样地的各指数沿海拔梯度呈先升高后降低趋势,嘉荫(海拔93 m)、双丰(海拔300 m)和桃山(海拔1 055 m)样地各指数次于五营(海拔447 m)、伊美(海拔469 m)和新青(海拔799 m)。草本层α多样性结果显示,各指数沿海拔梯度呈降低趋势,新青(海拔799 m)样地各指数表现最差,双丰(海拔300 m)样地各指数表现最好,可能是因为双丰样地紧临农业施业区,人为干扰较大,破坏了样地群落多样性,而桃山样地由于海拔较高,刺五加群落不存在乔木层,导致群落多样性数据异常,但纵观各样地各指数,在群落层面上均未表现出沿海拔梯度的较大变化。海拔400~500 m范围内,达到刺五加最为适宜的气候条件,群落结构特征完整,各指数表现最佳[21]。
五营样地乔木层群落和双丰样地草本层群落物种多样性高,物种丰富,均匀度较高,而且乔木和草本在组成上占优势,这与林分结构、立地条件和人为破坏程度有直接关系。伊美样地灌木层群落物种多样性高,且分布均匀,与其他层片相比具有明显优势,该样地乔木层优势树种为红松,林分树种组成简单,且伴生树种数量较少,分布不均匀,明显影响林下植物的物种分布和生长。
2.2.2 群落α多样性指数间的相关性分析
群落α多样性各指数对群落物种多样性的贡献度不同,表4为刺五加群落物种α多样性指数间的相关系数。可以看到,海拔与群落物种α多样性各指数间均呈负相关,与丰富度指数d2在0.05显著性水平上负相关,即随着海拔梯度升高,各群落多样性指数均呈下降趋势。刺五加群落α多样性各指数中,多样性指数D和H之间、多样性指数D与均匀度指数Jd之间、多样性指数H与均匀度指数Jd之间、多样性指数H与物种优势度C之间,呈P<0.01水平极显著正相关性。从海拔对刺五加群落α多样性各指数的贡献上看,物种丰富度贡献率最大,由大到小依次为:种的丰富度指数、种的均匀度指数和物种多样性指数。

表4 刺五加群落物种α多样性指数间相关系数Tab.4 The correlation coefficient between α diversity index of Acanthopanax senticosus community
2.2.3 群落β多样性沿海拔梯度的垂直变化
群落β多样性可表征沿海拔梯度更替物种替代的程度和不同群落间的物种多样性,反映不同海拔梯度群落间物种组成的差异性,不同海拔梯度群落共有种越少,群落β多样性越大。从刺五加群落β多样性指数(表5)可以看出,Cody指数βc随着海拔升高呈先降低后升高趋势,最低点出现在中海拔地区(447~469 m),为6.8,这说明2个海拔之间物种组成相似,物种更新速率较慢。Sorenson指数Cs随着海拔升高呈先升高后降低趋势,最高峰仍出现在中海拔地区(447~469 m),为0.81,是因为2个相邻海拔地区共有物种数最多,说明相邻中海拔地区群落间的差异性最小。海拔差距在300 m时,群落间物种组成差异较明显,生境被物种隔离程度较高。
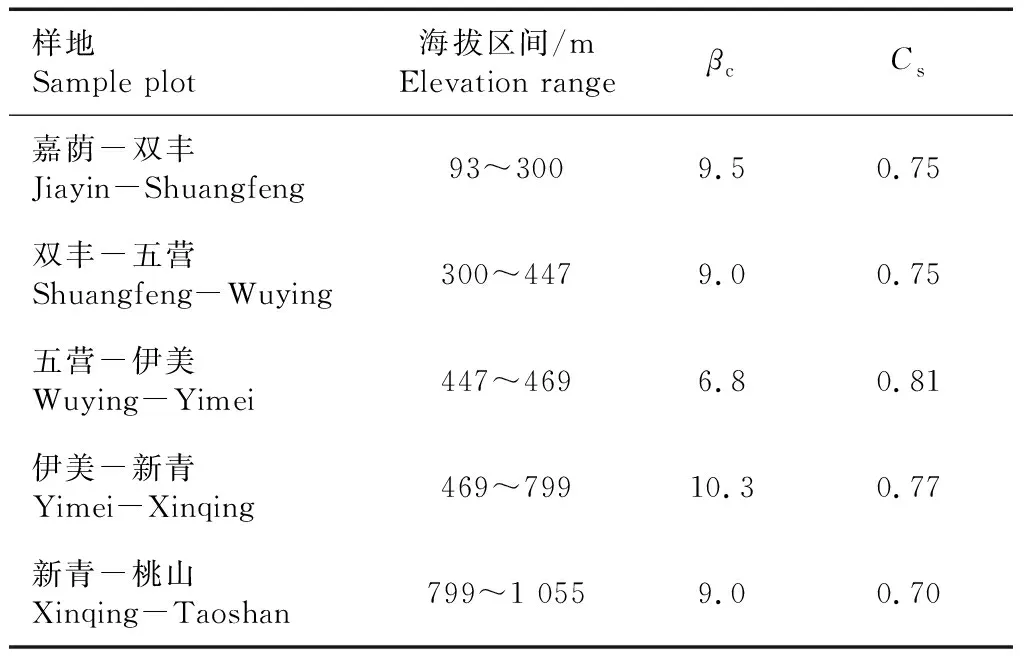
表5 刺五加群落物种β多样性指数Tab.5 Species β diversity index of Acanthopanax senticosus community
2.3 不同海拔刺五加群落的相似性系数
从表6可以看出,五营样地(海拔447 m)与伊美样地(海拔469 m)之间的群落相似性系数为0.37,在所有海拔样地之间最高,可能是由于两样地海拔最为接近,与其他样地相比拥有重叠植物种类数最多;伊美样地为红松针叶纯林,五营样地为阔叶混交林,虽然刺五加伴生种有些相同,但乔木层和草本层在物种组成上还是存在较大差异,导致群落相似性系数不高,只呈现中等不相似水平。其次五营样地(海拔447 m)与新青样地(海拔799 m)之间的群落相似性系数为0.32,呈中等不相似水平;剩余各群落之间的相似性系数为0~0.25,呈极不相似水平。桃山样地(海拔1 055 m)由于特殊的生境条件,物种种类明显与其他差异较大,与其他样地的群落相似性系数均较低。总体上看,海拔梯度相差越大,群落相似性越低;不同海拔刺五加群落之间呈现不相似水平,说明不同海拔梯度的样地生境变化较大,导致群落结构性差异较大,物种多样性变化也越明显。

表6 刺五加群落相似性系数Tab.6 The similarity index of Acanthopanax senticosus community
3 结论
(1)刺五加群落沿海拔梯度的物种组成变化为以灌木为主、草本植物为辅。刺五加为优势种群,次优势种群随着海拔升高发生相应改变。除海拔300 m和1 055 m无花楷槭、海拔93 m和1 055 m无东北茶藨子,其他海拔梯度上均有较大比例分布,作为群落中的次优势种存在。随着海拔升高,刺五加群落中乔木、灌木和草本树种物种组成变化不明显,只有海拔达到1 000 m以上,原有物种几乎全部消失,随之出现蹄叶橐吾、小花风毛菊和北乌头,说明海拔、立地条件和生境条件等多因素控制刺五加群落物种组成。
(2)海拔梯度、纬度梯度等多种环境梯度对群落α多样性具有较大影响。相关学者极度重视海拔梯度这个环境梯度格局中的主导梯度[22],五营样地乔木层群落和双丰样地草本层群落物种多样性高,物种丰富,均匀度较高,而且乔木和草本在组成上占优势,这与林分结构、立地条件和人为破坏程度有直接关系。伊美样地灌木层群落物种多样性高,且分布均匀,与其他层片相比具有明显优势,该样地乔木层优势树种为红松,林分树种组成简单,且伴生树种数量较少,分布不均匀,明显影响林下植物的物种分布和生长,但纵观各样地各指数在群落层面上,均未表现出沿海拔梯度较大的变化。海拔400~500 m范围内,达到刺五加最为适宜的气候条件,群落结构特征完整,各项指数表现最佳。从海拔对刺五加群落α多样性各指数的贡献上看,物种丰富度的贡献率最大,海拔与各指数均呈负相关。
(3)物种β多样性指数差异不大,指数在海拔799 m时出现最大值,可见新青过渡带物种变化速率较快。
(4)桃山样地(海拔1 055 m)由于特殊的生境条件,物种种类明显与其他差异较大,与其他样地的群落相似性系数均较低。总体上看,海拔梯度相差越大,群落相似性越低;不同海拔刺五加群落之间呈现不相似水平,说明种群扩散与种群定居均受海拔影响,不同海拔梯度的样地生境变化较大,导致群落结构性差异较大,物种多样性变化也越明显。
(5)从刺五加物种多样性的海拔格局来看,海拔相近的区域,由于相似的地形条件致使物种多样性间差异不显著,研究未对小兴安岭南坡以外的海拔影响因子加以分析,但是较小海拔范围内其他因素的影响作用依然明显。因此,应加强针对刺五加群落物种多样性在大空间尺度上的差异、种群迁移和演替进程等在海拔影响因子等方面研究。
猜你喜欢
今日农业(2022年15期)2022-09-20
建筑与预算(2022年6期)2022-07-08
农村实用技术(2020年1期)2020-12-16
乡村科技(2018年8期)2018-02-13
现代园艺(2018年14期)2018-01-18
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
中成药(2016年4期)2016-05-17
特产研究(2014年4期)2014-04-10
中国民间疗法(2014年3期)2014-01-25
