
微塑料影响抗性基因的传播与水平基因转移
2022-08-24 00:19:56吴效俭施国静王莹莹
中国环境科学 2022年8期
吴效俭,施国静,王莹莹*
微塑料影响抗性基因的传播与水平基因转移
吴效俭1,2,3,施国静1,2,3,王莹莹1,2,3*
(1.南开大学环境科学与工程学院,天津 300350;2.环境污染过程与基准教育部重点实验室,天津 300350;3.天津市城市生态环境修复与污染防治重点实验室,天津 300350)
微塑料作为一种新兴污染物,在多种环境介质(大气、土壤、水体、沉积物)中均有广泛检出.微塑料能够通过表面形成的生物膜结构携带抗性基因,对人体健康造成的潜在风险已经引起了国内外学者的广泛关注.但是,微塑料在抗性基因的长距离传播以及水平转移方面所发挥的功能,以及微塑料传播耐药性所导致的生态与环境效应还尚不明晰.本文重点对不同环境中的微塑料与抗性基因的污染现状进行了综述讨论,对微塑料传播抗性基因的主要方式与过程进行了对比分析,最后概述了微塑料参与调控抗性基因水平转移的机制,并对未来相关研究的重点方向进行了展望,以促进微塑料污染问题的科学解决.
微塑料;塑料;抗生素抗性基因;塑料际;污染特征
塑料作为一种人工合成或半人工合成的高聚物,被广泛应用于工业生产与日常生活中.2015年的塑料全球年产量已达到4.07亿t[1].大量塑料制品的使用会导致塑料废弃物的产生,预计在2050年,产生的塑料废弃物会突破2´107t[2].塑料废弃物进入环境后,经过逐级裂解,形成的粒径在5mm以内的塑料碎片被称为微塑料[3].在各类环境,如土壤[4-5]、大气[6-7]、水体与沉积物中[3,8-9],都有微塑料的广泛分布.由于尺寸较小,部分微塑料会进入浮游动物体内,这些含有微塑料的浮游动物被处于更高营养级的鱼、虾等水生动物捕食[10-11],进而在食物链中逐级积累,发生富集作用[12-14].当人类食用这些海鲜与水产品时,微塑料随食物进入人体消化系统,对人体健康具有潜在威胁[15].
环境中的微塑料表面会被微生物所定殖,形成生物膜结构,即不同种类的微生物构成的群落[16-17].通过生物膜,微塑料可以携带耐药细菌(ARB).由于耐药细菌拥有抗生素抗性基因(ARGs),因此能够对不同种类的药品产生抗性[18-20].携带耐药细菌的微塑料可以在不同类型的环境中迁移,进而产生诸多问题,其中抗性基因的水平基因转移(HGT)是尤为严重的一个方面.水平基因转移被认为是细菌耐药性传播的主要驱动因素[21],抗性基因可以通过质粒,整合子,转座子等可移动元件(MGEs)进行迁移[22],造成细菌耐药性的传播与扩散[23-24].因此揭示微塑料对抗性基因水平转移的机理,将有助于进一步明晰抗性基因的携带与传播作用,以及微塑料参与调控不同环境中微塑料的迁移过程对环境安全与人体健康的影响.
本文主要从细菌耐药性的角度,重点讨论了不同环境中微塑料与抗性基因的相关研究进展,旨在分析以耐药细菌为代表的微生物与微塑料污染之间的联系,以及两者作为一个整体所产生的环境生态效应.
1 环境中微塑料的赋存与抗性基因污染
微塑料与抗生素抗性基因被认为是近年来的环境新兴污染物[25-26].携带抗性基因的细菌可以从多个方面影响人体健康,例如通过饮用水、空气、开放水域等传播导致人体感染[27].微塑料作为一种可供微生物定殖的微生境,也具有携带并传播抗性基因与人体致病菌的功能.一些人体条件致病菌,例如,等会在微塑料表面富集[17,20].由于微塑料的难降解性,其在环境中有较长的赋存时间,进而增加人体对抗性基因的暴露风险.在不同环境介质(土壤环境、大气环境、水体与沉积物环境)中,由微塑料所携带的抗性基因的种类与丰度信息见表1.

表1 不同环境介质中微塑料携带的抗性基因种类与丰度
注: -为无丰度数据.
1.1 大气环境中的微塑料与抗性基因
在城市区域,空气、道路扬尘、室内灰尘中分布有大量微塑料和抗性基因[2,36].室内的装饰品、织物等会释放出大量微塑料,在室内空气中漂浮,据研究,城市室内环境的微塑料浓度可达16个/m3[37].随着门窗开启与空气流通,室内环境中的微塑料会扩散到室外空气中.汽车轮胎经过长期磨损后会释放出微塑料,部分进入空气中,这种现象在车流量较大的城市主干道路附近更加明显.漂浮在空气中的微塑料易吸附空气中漂浮的抗生素抗性基因,与其共同发生迁移.
由于尺寸较小和重量较轻,微塑料很容易在风力作用下向较偏远的区域发生迁移,在一些受人为活动影响较小区域的空气中,也发现了气载微塑料的存在[38-39].一项针对法国山脉附近的气载微塑料的研究发现,微塑料通过大气传输的距离能够达到95km[40].抗性基因在大气环境中的分布方面存在一定规律,全球尺度的大气环境中,β-内酰胺类抗生素抗性基因bla,以及喹诺酮类抗性基因具有较高的丰度[39].在空气重度污染地区,碳氢霉烯类抗性的基因bla-1占有大气总抗性基因丰度的30%[41].
漂浮在空气中的微塑料与抗性基因极易被人体吸入[42],导致呼吸系统的疾病[43],损害人体健康.每个成人每日吸入的微塑料颗粒数约为20个[44].在人体肺部组织中探测到了合成纤维的存在[45],且婴儿和儿童的微塑料吸入量远高于成人[6].空气气溶胶中存在着各类携带抗性基因的人体条件致病菌,如,等[39,46].当这些致病菌以及抗性基因黏附到微塑料表面进入人体肺部后,容易造成肺部感染等疾病.由于目前关于大气环境中微塑料所携带的抗性基因方面的研究还较少,因此对于评估抗性基因与微塑料的联合毒性效应还存在较大难度,同样地,对于抗性基因随气溶胶中的微塑料进行长距离迁移的潜力也有待进一步探究.
1.2 土壤环境中的微塑料与抗性基因
土壤环境,尤其是农田土壤中,存在大量微塑料,这是由于耕作过程中广泛使用的地膜等塑料制品在光照、风蚀、撞击等作用下,物理化学性质发生改变,逐渐裂解形成微塑料.特定的土壤微生物能够在微塑料表面形成细菌群落,导致微塑料携带的细菌种类和抗性基因组成与周围环境中的细菌群落均存在差异[47-48].土壤环境具有区别于大气与水环境的独特性,土壤的团粒结构组成成分相对复杂,其中各类杀虫剂、农药和重金属的存在导致土壤细菌同时受到多种污染物的影响,当细菌对外源刺激做出响应与反应,会进一步影响土壤细菌群落中抗性基因的丰度[49].土壤中的微塑料能够吸附土壤中的各类污染物,如溴代阻燃剂[50]、重金属[51]、抗生素[52],是一个独特的微生境.具有编码抗生素与重金属耐性基因的细菌能够通过外排泵[53]、降低膜通透性[54]、转化或降解污染物[55]等方式,更好地适应生境,从而定殖到微塑料表面.因此,土壤中的微塑料会携带并富集抗性基因,进而影响土壤环境中的抗性基因组成[30,34].
在污水处理厂中,汇聚了大量不同来源的生活、医疗、农业养殖污水,存在不同类型的微塑料与抗性基因[56].携带抗性基因的微塑料进入土壤的另一个重要来源是污水处理厂的活性污泥[57-58].经过絮凝与沉淀过程,污水中的微塑料进入活性污泥中[59],之后随污泥一同被填埋、焚烧、作为农田肥料,从而进入到土壤中[57],构成了土壤环境中抗性基因的重要点源污染.研究显示,污水处理厂污泥中的PVC微塑料是传播致病菌以及抗性基因的媒介[60].一项针对好氧颗粒污泥系统的研究显示,进入污泥中的微塑料会使体系内携带抗性基因的细菌分裂速率发生变化,对于赋存于污泥中[32]、渗滤液中[61]和接收污水处理厂经过处理后的废水的河流中[62]抗性基因的增殖情况均有明显影响.
污水处理厂的活性污泥是微塑料进入土壤环境的重要来源,但目前各反应阶段(如沉淀、絮凝、生化处理、消毒等)对微塑料以及表面抗性基因的种类与丰度变化的影响还了解甚少.因此,有必要针对关键的分离与转化步骤对微塑料的去除效率,以及表面生物膜中抗性基因的赋存情况进行研究,为评估特定步骤的物理化学条件对微塑料传播抗性基因的能力的影响.
1.3 水环境中的微塑料与抗性基因
海洋汇聚了河流、湖泊以及来自大气沉降的微塑料,被认为是一个重要的微塑料的汇[28,63].目前,多项研究证实了各个大洋、河流、内陆湖、集水区等水域[64-67]中均有微塑料的存在,甚至一些受人为活动影响较小的地区,如格陵兰岛附近的海域[68]也有微塑料的检出.微塑料能够携带并富集水体中的抗性基因[57,69-72].多项关于水体中微塑料表面生物膜的研究显示,无论是抗性基因的种类还是丰度,微塑料携带抗性基因的能力均高于水体,其表面形成的生物膜结构对抗性基因具有富集作用[17,19,57,69,71-72].一些抗性基因的细菌宿主(,与)只在微塑料表面被发现,而没有出现在其他自然介质表面(如岩石和叶片中),是微塑料所具有的材料特性所导致的[17].
目前的大部分研究关注的是水介质中的微塑料表面抗性基因的赋存情况,而对于沉积物的研究还较少,这可能是受到采样设备与条件的限制,以及沉积物环境中微塑料表面生物膜较难收集导致[73].由于微塑料在老化、生物污损等因素的作用下,会在水体环境中发生垂直迁移,从而进入沉积物中.沉积物环境中生活着多种底栖动物,微塑料所携带的抗性基因与这些生物的相互作用也有待进一步研究.
1.4 大气、土壤、水体环境中微塑料表面抗性基因污染的异同点
在水环境、土壤环境以及空气环境中的微塑料均被发现能够携带抗性基因,但存在于不同环境介质中微塑料表面形成的生物膜的群落组成与生物量存在差异,导致了其携带抗性基因能力的不同.其中,空气环境中微塑料携带抗性基因的情况与土壤、水体环境中存在很大差异,主要表现在微塑料表面生物膜的生物量较低,携带的抗性基因种类较少,丰度较低.这主要是单位体积空气中细菌的丰度小于单位体积的水或者土壤所导致的.
而土壤与水环境中蕴含着较丰富的营养物与多种细胞代谢所需的矿物质元素,能够支持存在于水、土介质中具有不同生理特性的微生物对微塑料表面的定殖过程,一些适应微塑料生境的细菌在微塑料表面逐渐形成生物量高、物种数量繁多、结构致密的生物膜系统,微塑料是众多细菌以群落为单位进行繁殖和演替的微型生境.微塑料表面以生物膜形式存在的细菌群落对环境因子的剧烈变化具有更强的抵抗能力.不同细菌有各自精细的分工,专营特定的生理反应与代谢活动,产生的各类次生代谢物可以释放到胞外,进入生物膜基质中,供给群落中的其他细菌成员使用.细菌群落作为整体,具有更加多样化的功能.部分细菌通过携带特定基因片段,可以利用转化、外排泵等方式将药物对细菌菌体的杀伤力降至最小,从而发挥出耐药性.在水体与土壤环境中的微塑料携带的抗性基因具有更高的多样性以及丰度.研究证实,微塑料表面生物膜中耐药细菌以及抗性基因的丰度比水体环境中至少高几个数量级[69,74].
现有的研究显示,土壤与水环境中微塑料表面携带的抗性基因的情况有所差异,这主要是由于土壤与水体环境中占有优势主导地位的细菌群落不同所导致的.具有不同分类学地位的细菌宿主会编码不同的抗性基因序列.水体与土壤中微塑料具有各自独特的表面性质特征,对表面的细菌群落产生特异性选择作用.例如,多项研究显示水体环境中微塑料对特定类群的细菌具有富集作用[75-76]另一方面,水体与土壤环境的多项环境因子均具有差异性,导致微塑料中含有的添加剂的析出程度不同,这也是造成水与土中微塑料携带抗性基因类别具有差异的重要原因.
污水处理厂、耕地、人体呼吸道等生境中,均存在核心抗性基因的现象[77-79].例如,研究收集了来自种植玉米、花生和大豆耕地的105份土壤样品,发现即使这些土壤中所生长的植物种类、土壤养分条件以及地理位置(样本之间相距距离最长2200km)不同,有一些基因在超过50%的样本中均被检测到,例如多重耐药型抗性基因,-05,以及-04等[80].微塑料是赋存于大气、土壤与水体环境中一类高度分化的生态位,但是目前对其中赋存的核心抗性基因的组成方面的了解还较少.多数研究区域面积较小,且种类单一,例如湖泊的近岸区、农田土壤的局部,得到的规律性结论在推广到其他环境时存在一定的困难.另外,多数研究受到微塑料生物膜生物量以及采样方法等条件的限制,所涵盖的样本量通常较少,对于解析抗性基因的丰度关系方面存在一定困难.因此,未来有必要在多环境介质、较大区域尺度范围内进行联合研究,以此为基础探讨微塑料对富集核心抗性基因的现象以及其中包含的规律.
2 微塑料表面生物膜的成因与影响条件
在自然环境中,多数土壤介质以及几乎所有水体中的固体表面均能够支持生物膜的生长[81].微塑料表面生物膜的形成条件主要包括以下几个方面:
2.1 固体介质表面性质
在微塑料生物膜形成的最初始阶段,需要固体介质表面为环境中存在的种类繁多的细菌提供附着与定殖的空间.只有细菌成功定殖到微塑料表面后,才能启动生物膜后续形成过程(图1).因此,固体介质微塑料的存在,是生物膜形成的基础.由于部分塑料材质的医疗用品,如导尿管,在制作加工时表面使用了聚两性离子涂层,具有抑菌效果[82].为了减少致病菌在导尿管上形成生物膜,部分导尿管的塑料材质表面还经过纳米尺度的结构修饰,细菌无法很好地黏附到管壁上,也就无法形成生物膜,从而降低尿路感染引起的炎症效应[83].相比于未经物理与化学改性修饰的普通微塑料,表面覆盖有特殊涂层,经过修饰的塑料制品经过裂解后形成的微塑料更加难以被细菌定殖,形成生物膜的能力较弱,携带抗性基因的能力较差.
环境中通过鞭毛等结构进行自由移动的细菌接触到微塑料表面后,松散地吸附到材料上.这个阶段的黏附是可逆的,也就是说,一部分细菌还会离开微塑料表面,重新回归为浮游状态.而部分细菌通过开始表达与黏性蛋白相关的基因[84],合成相关黏附素,将菌体紧密地固定到微塑料表面,同时鞭毛等主管移动的附件结构的活动性也发生变化,细菌对表面的吸附进入不可逆的状态[85],至此,细菌完成对微塑料表面的定殖过程,这些细菌被称作先锋种.先锋种细菌在微塑料表面进行细胞分裂与繁殖,改变了介质的表面性质,为后续其他种类细菌在微塑料表面形成细菌群落提供了先导条件.
2.2 营养与水文条件
在经历了先锋种的定殖后,逐渐有更多种类的细菌开始在微塑料表面稳定存在,由于微塑料作为一类微型生境,能够提供的空间与营养是有限的,因此周围环境中介质的更新速度决定了细菌获取营养以支撑群落发展的速率以及规模,也对生物膜的形貌特征具有决定性作用[86].有研究显示,在水体环境中,液体的流速影响了固体介质表面生物膜的物理结构,主要表现为在一定范围内,流速越高,生物膜的生物量越大[87].这主要是水流能够为生物膜带来更高浓度的营养物质,同时将产生的多余代谢物带走,不断进行局部环境的更新.但是,当水流流速过高时,生物膜结构无法承受水流剪切力作用,表面的细菌会从生物膜中剥离,导致生物量的减少[88-89].在特定的河流、湖泊等环境中,水流的速度大致在一定范围内进行波动,其中微塑料表面生物膜的生物量以及群落中细菌总量稳定在特定水平,其携带抗性基因的能力是相对稳定的.
2.3 其他环境因子
多种环境因子,例如,光照条件、温度、pH值、培养时间等均会从不同方面对生物膜中细菌的生长造成影响,从而改变生物膜的群落特征.研究显示,在黑暗条件下,蓄水库中生物膜的厚度在30μm以下,在有光照的条件下,蓄水库中生物膜的厚度有了大幅度的增加,约为50~130μm[90].温度对生物膜的生长速率有十分重要的影响.当水体温度过低时,细菌的细胞进入休眠状态,只进行最基本的代谢活动,生物膜的组成不再发生大的变化[91].研究显示,当微塑料在污水处理厂污泥中的存在时间超过60d时,微塑料所携带的抗性基因的丰度有显著增加[60,92],这表明微塑料在特定生境中的赋存时间对携带抗性基因的细菌在介质表面的增殖以及向周围环境中的扩散产生的影响是直接的,进而间接影响该生境中抗性基因的组成与丰度.
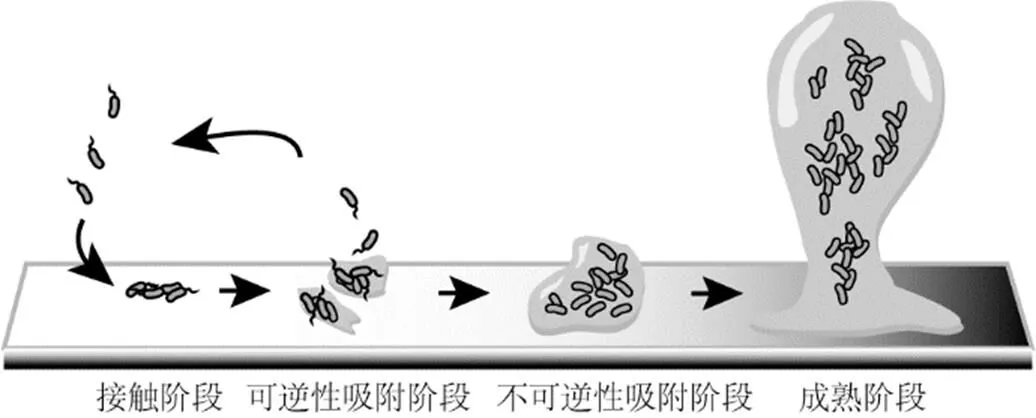
图1 微塑料表面生物膜的形成条件
在微塑料所处的特定生境中,所有环境因子共同对生物膜群落的变化与演替施加影响.经过各个细菌类群的比例动态变化的阶段后,逐渐达到群落结构相对稳定的状态,主要表现为养分分配的平衡,对外界环境因子的剧烈变化有一定的抵抗能力.这个稳定的状态并非是各个种属的细菌比例保持一成不变,而是生物膜群体能够随着外界环境条件随时调整自身结构形成的动态平衡.但是,目前的研究多关注于微塑料表面经过长时间培养后形成的生物膜结构,对于其动态形成过程中群落组成的变化以及抗性基因的丰度波动还需要更加详细的探究.
3 微塑料影响抗性基因传播的途径
3.1 微塑料的理化性质影响抗性基因传播
通过野外调查与实验室模拟,多项研究表明,抗性基因在微塑料表面生物膜中的赋存与传播受到微塑料理化性质的影响,主要包括介质的化学组成(材质)、粒径和老化程度等方面.
3.1.1 微塑料的化学组成影响抗性基因的传播 一项针对渗滤液中微塑料的研究显示,聚氯乙烯(PVC)、聚丙烯(PP)、聚苯乙烯(PS)所富集的抗性基因种类与丰度具有差异[92],这可能是由于不同的化学组成影响了微塑料表面所定殖的菌群的结构,从而导致所携带的抗性基因的差异.
塑料高聚物的种类也同样影响微塑料对于抗性基因的丰度富集效果.例如,河口环境与渗滤液微宇宙系统中的聚乙烯(PE)微塑料生物膜中的抗性基因的丰度均高于PP微塑料[92-93].不同化学组成的微塑料中所含有的添加剂成分也不同,对于不同细菌类群在微塑料表面形成生物膜的影响具有差异,其促进或抑制效应需要针对具体细菌类群的情况进行分析.
3.1.2 微塑料粒径与老化程度影响抗性基因传播 有研究通过批次培养实验,对不同粒径范围的微塑料表面生物膜群落组成进行了探究,分别对比了粒径范围在106~125μm与355~425μm的PS微塑料,粒径范围在125~250μm与250~500μm的PE微塑料,两组实验结果显示,粒径范围并未显著影响微塑料的表面微生物群落结构,其携带的抗性基因的差异也较小[94].
微塑料进入环境后,经历一系列物理、化学、生物过程导致其表面性质发生改变的现象称为老化.物理方面,在机械磨损作用下会发生分子链的断裂;化学方面,在紫外线照射下,微塑料表面发生光氧化,涉及官能团的改变;微生物作用下,其表面的粗糙度、电荷含量以及表面自由能变得不同,含有的添加剂更容易析出,且更容易吸附来自外部的污染物,如抗生素类的药品以及重金属等.微塑料老化后,材料的疏水性发生变化,表面形成的生物膜中更容易积累重金属、有机污染物、抗生素等物质[49,95].这些物质对细菌具有定向选择压力,不仅会导致耐药性细菌的增加,也会直接影响耐药细菌中质粒携带的抗性基因发生转移的速率(图2).
研究显示,微塑料表面吸附的污染物对于微塑料表面抗性基因的丰度具有促进作用[33].在甲氧苄氨嘧啶的作用下,抗性基因水平转移与垂直转移速率均显著增加[96];重金属与多环芳烃的存在,也会对抗性基因的转移效率产生影响[97].从塑料中分离出的细菌具有更强的制造胞外多聚物的能力,因此,当具有这些能力的细菌定殖于塑料表面时,有助于形成致密的生物膜结构[98-99].塑料防污涂层中含有的铜元素与锌元素能够使微塑料富集更高丰度的大环内酯类-林可胺类-链霉素类(MLS)抗性基因[100].在种植蔬菜的农田土壤中经过自然老化的微塑料吸附的抗生素与重金属含量更高,也增加了可移动性遗传元件的丰度,这对于微塑料表面抗性基因的传播有较多贡献[101].
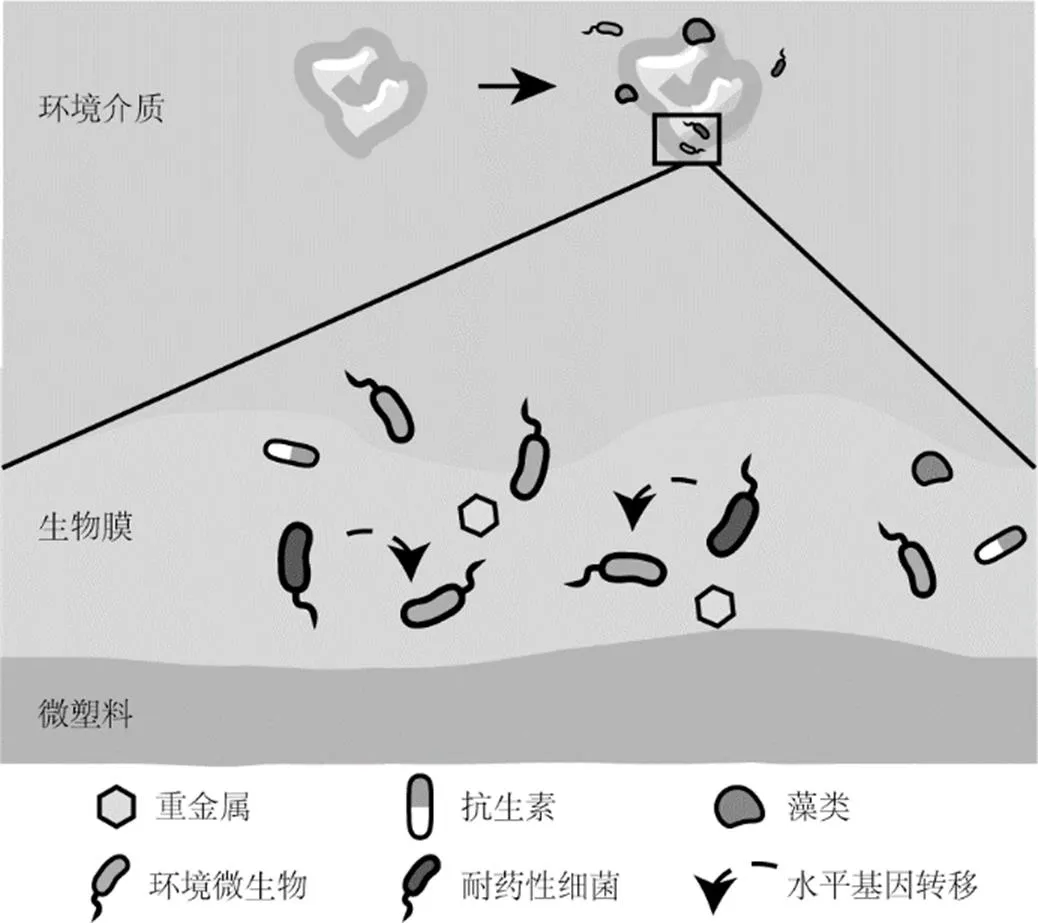
图2 微塑料表面生物膜影响抗性基因的传播
3.2 微塑料在环境介质中的迁移影响抗性基因传播
3.2.1 微塑料在环境介质内部的迁移 在水体环境内部,微塑料起初多数存在于表层水体,之后在表面生物膜的影响下向深层水体以及沉积物中迁移.当微塑料表面的生物膜被部分以细菌为食的浮游生物摄取后,微塑料的密度发生变化,可能会重新悬浮到水体表层.该过程不断循环,伴随着微塑料的沉降与再悬浮,所携带的抗性基因也随之进入水体环境的各个组分中[102].土壤中微塑料的移动主要是依靠土壤动物翻动土壤,微塑料随着土壤颗粒共同运动,其运动路径大体遵循由浅层土壤进入深层土壤的趋势.大气环境中存在的常常是密度较轻的微塑料,其运动受到风力、降水等因素的影响较多[103-104].
3.2.2 微塑料在多环境介质之间的迁移 河流是微塑料从陆源向水体环境中迁移的重要媒介.污水处理厂接收的废水中含有的大量微塑料无法被完全去除,依旧存在于水介质中,随着末端排放进入河流中.河流中携带着大量微塑料汇入海洋.据统计,全球90%的微塑料是通过河流输送到海洋、内陆湖等环境中的[105].
另一方面,大气的全球循环是水体、空气、土壤中的微塑料向其他介质中迁移的重要渠道.在风力与气流的作用下,微塑料进入空中,之后在沉降作用下进入陆地与海洋环境[103-104],但是目前气载微塑料的流通过程与通量的计量还需要结合模型进行解析与评估.
以生物膜形式存在的细菌,其生活方式区别于浮游态的细菌.胞外多聚物结构对于细菌菌体的黏附聚集、形成特定的空间结构、抵抗外界环境因子波动以及细菌群落内部的交流和耐药性方面,都具有十分重要的作用.致密的胞外多聚物可以加强微塑料表面细菌之间的联系,能够增加包括质粒、噬菌体、基因岛、转座子和插入序列等在内的可移动遗传元件的丰度,促进不同种属的细菌之间抗性基因的交流.已有研究显示,水环境中微塑料携带的耐药性细菌丰度是水体中的100倍[106],微塑料生物膜中的细菌获得质粒的频率约为浮游态细菌的两倍[107],质粒携带抗性基因的序列在环境微生物与耐药性细菌之间转移,增加了普通细菌获得耐药性的可能性.其他实验数据也表明微塑料携带的抗性基因具有较高的基因转移速率[69,106].微塑料作为一个汇集了耐药性细菌、污染物、抗生素的微生境,细菌之间的联系与交流都强于环境中浮游状态的细菌,对环境抗性基因组也会产生较大的影响.微塑料通过在多种介质之间的移动,会导致携带的抗性基因在表面生物膜与所经过的环境微生物之间发生交换.这种抗性基因的传播效应可以在长距离尺度发生[49,108].
生物膜作为一个具有高度自组织结构的整体,部分细菌类群数量始终处于不断变化中,这样能够更好地适应不断变化的外界环境.处于不同演替阶段的微塑料表面生物膜在传播抗性基因方面所发挥的功能有何种不同还有待进一步探索.另一方面,土壤固体团粒结构以及水流流体具有不同的介质特点,介质中存在的污染物的种类以及可迁移性方面也会影响抗性基因的传播,相关的具体过程以及内部调控机制也尚未明晰.最后,针对微塑料在不同生态系统中发生老化的过程,对于携带抗性基因的能力的影响的评估也十分重要,能够为评估微塑料的环境行为以及生态风险评估提供理论支持.
4 微塑料影响抗性基因水平转移的过程与机制
微塑料所携带的可移动遗传元件为抗性基因的水平基因转移提供了基础[109].整合子能够捕获并整合外源性基因,并将其转变为功能性基因的表达单位,与转座子或者接合性质粒共同使抗性基因在不同细菌之间发生水平传播[110].例如,抗性基因1位于I类整合子的3’-保守末端(3’-CS),当整合子在不同细菌之间移动时,所携带的1也会随之转移[111].此外研究报道在微塑料表面有部分可移动遗传元件能够同时携带重金属抗性基因(MRGs)和抗生素抗性基因,且两者的丰度具有显著相关关系[70].目前,对微塑料如何调控抗性基因水平转移方面的机制还有待进一步了解.对近5年报道的几类主要水平基因转移方式进行总结分析,微塑料在这些过程中所发挥的功能与调控作用见图3.
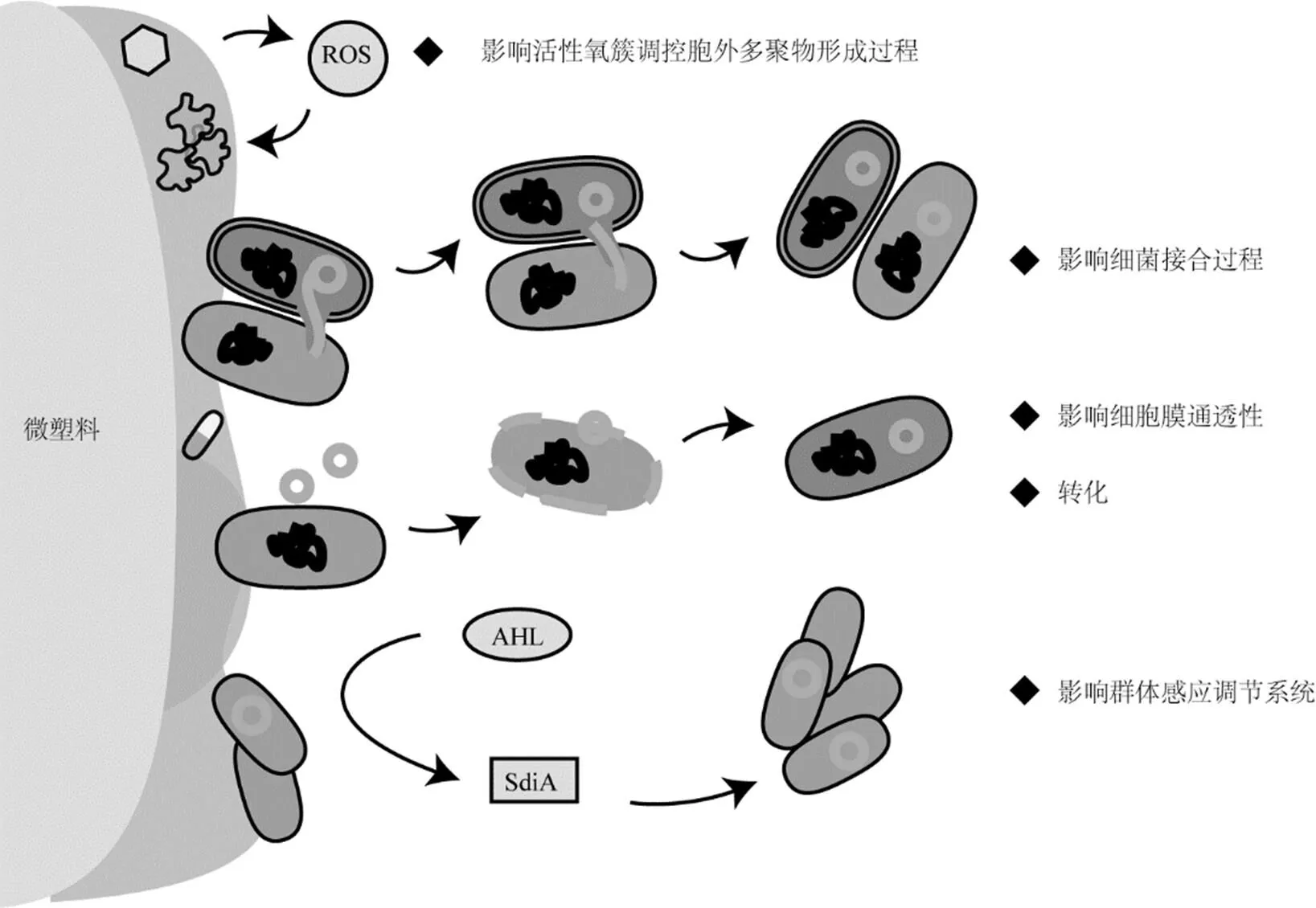
图3 微塑料影响抗性基因转移的机制
4.1 微塑料影响细菌的接合过程
与浮游态的细菌相比,微塑料表面的细菌具有更高的基因转移频率[109].抗性基因能够通过接合的方式发生水平转移.接合(Conjugation)是供体菌通过性菌毛将DNA传递给受体菌,从而将抗性基因整合到受体菌的基因组中,使其表达出相应的抗性性状.接合方式介导的基因转移以供体菌通过性菌毛与受体菌发生直接接触为基础[23],微塑料会对接合过程的发生频率产生影响.聚苯乙烯微塑料会极大程度地促进MG1655的pKJK5质粒向sp.细菌以及天然湖泊中细菌的接合转移,这表明微塑料不仅会影响模式细菌在纯培养体系中的接合频率,也会影响自然环境中的细菌的接合转移[107].微塑料表面可以吸附其他类型的污染物,例如纳米材料(ENMs)[112]以及药物和个人护理品(PPCPs)[113]等.这些吸附的污染物(如ZnO、Al2O3、GO、Ag、CuO、TiO2、NaNO2、TCS等)已被证实能够不同程度地提高细菌的接合效率[114-119].
4.2 微塑料影响活性氧簇调控胞外多聚物形成过程
微塑料表面附着的生物膜具有胞外多聚物结构,含有的多糖、核酸、蛋白质、脂质以及其他小分子,对细菌相互作用与信息交流具有重要作用[120].当胞外多聚物的含量较高时,会促进细菌的黏附作用,而胞外多聚物含量较低时,则会通过静电排斥力的作用抑制细胞黏附过程[121].由于胞外多聚物的形成过程受到胞内活性氧簇(ROS)浓度的影响[122],活性氧簇的浓度又受到污染物暴露的影响[123-124].微塑料表面吸附的污染物(如重金属离子等),会通过影响细菌胞内活性氧簇浓度水平的方式,间接影响影响表面生物膜的形成.由于适度的活性氧簇暴露会引发细胞一系列保护性措施,也有利于增加水平基因转移发生的频率[125];但是,当所接触的污染物含量较高时,过量的活性氧簇也会导致细胞损伤甚至死亡,导致接合频率的下降[126].
4.3 微塑料影响细胞膜通透性与群体感应调节系统
与浮游细菌相比,微塑料表面以生物膜状态存在的细菌,其营养策略、细胞间的交流、细胞膜通透性均具有不同的特征,这对抗性基因的转移也会构成影响.当细胞膜通透性较大时,疏水的脂质屏障被削弱,促进抗性基因的释放与吸收.暴露于微塑料表面污染物的细菌细胞膜容易形成孔洞,使供体菌的性菌毛更容易与受体菌接触,从而发生接合作用[127].
群体感应调节系统(QS)支持了生物膜中细胞与细胞之间的通讯,能够通过信号分子获取自身以及其它细菌种群的数量信息,当细菌的密度达到一定阈值时,会通过启动相关基因来调控细菌群落的密度水平[128].调控方式主要是依靠特定信号分子的浓度实现的,如N-酰基高丝氨酸内酯(AHLs).这些信号分子也参与了生物膜中抗性基因水平转移的过程.但是不同种类的AHLs的调控作用存在差异,例如,C4-HSL信号分子会通过启动AHL效应器SdiA,抑制部分细菌的种间接合转移[129];而另一项研究显示C6-HSL信号分子对细菌E. coli HB101向E. coli NK5449的属内接合过程产生促进作用[130].因此微塑料生物膜中的群体效应的信号分子如何调控抗性基因传播的机制还需进一步的研究.
5 总结与展望
5.1 目前的研究已经证实携带抗性基因的微生物能够以微塑料为载体进入到不同营养级的生物体内,与肠道菌群发生相互作用,导致抗性基因的传播、进化.未来还需要重点研究由微塑料携带的抗性基因、环境中的由浮游生物携带的抗性基因、以及生物肠道菌群内的抗性基因的共有部分以及同源性,以揭示抗性基因在不同介质中的扩散潜力以及进化速度.
5.2 目前关于大气环境中微塑料携带的抗性基因方面的研究还较少,评估抗性基因与微塑料的联合毒性效应还存在较大难度,因此抗性基因随气溶胶中的微塑料进行长距离迁移的潜力亟需进一步探究,以便全面评估由吸入等途径进入人体的微塑料所产生的人体健康效应.
5.3 当前研究多关注抗性基因在微塑料表面的赋存情况,对于微塑料导致抗性基因水平转移的频率增加的机理与调控机制还尚未明晰,因此需要研究微塑料参与抗性基因水平转移的各个步骤,从而对抗性基因的传播、功能以及生态作用提供全面的概述,全面加深微塑料迁移过程对生态稳定性的影响.
[1] Rhodes C J. Plastic pollution and potential solutions [J]. Science Progress,2018,101(3):207-260.
[2] Wright S L,Ulke J,Font A,et al. Atmospheric microplastic deposition in an urban environment and an evaluation of transport [J]. Environment International,2020,136:105411.
[3] Isobe A,Iwasaki S,Uchida K,et al. Abundance of non-conservative microplastics in the upper ocean from 1957 to 2066 [J]. Nature Communications,2019,10(1):417.
[4] Accinelli C,Abbas H K,Shier W T,et al. Degradation of microplastic seed film-coating fragments in soil [J]. Chemosphere,2019,226:645-650.
[5] Beriot N,Peek J,Zornoza R,et al. Lowdensity-microplastics detected in sheep faeces and soil: A case study from the intensive vegetable farming in Southeast Spain [J]. Science of the Total Environment,2021,755:142653.
[6] Abbasi S,Keshavarzi B,Moore F,et al. Distribution and potential health impacts of microplastics and microrubbers in air and street dusts from Asaluyeh County,Iran [J]. Environmental Pollution,2019,244:153-164.
[7] Amato-Lourenco L F,Galvao L D S,De Weger L A,et al. An emerging class of air pollutants: Potential effects of microplastics to respiratory human health? [J]. Science of the Total Environment,2020,749:141676.
[8] Cozar A,Echevarria F,Ignacio Gonzalez-Gordillo J,et al. Plastic debris in the open ocean [J]. Proceedings of the National Academy of Sciences of the United States of America,2014,111(28):10239-10244.
[9] Isobe A,Uchiyama-Matsumoto K,Uchida K,et al. Microplastics in the Southern Ocean [J]. Marine Pollution Bulletin,2017,114(1):623- 626.
[10] Desforges J P W,Galbraith M,Ross P S. Ingestion of microplastics by zooplankton in the Northeast Pacific Ocean [J]. Archives of Environmental Contamination and Toxicology,2015,69(3):320-330.
[11] Martinez-Gomez C,Leon V M,Calles S,et al. The adverse effects of virgin microplastics on the fertilization and larval development of sea urchins [J]. Marine Environmental Research,2017,130:69-76.
[12] Frias J P,Otero V,Sobral P. Evidence of microplastics in samples of zooplankton from Portuguese coastal waters [J]. Marine Environmental Research,2014,95:89-95.
[13] Berglund E,Fogelberg V,Nilsson P A,et al. Microplastics in a freshwater mussel (Anodonta anatina) in Northern Europe [J]. Science of the Total Environment,2019,697,134192.
[14] Adika S A,Mahu E,Crane R,et al. Microplastic ingestion by pelagic and demersal fish species from the Eastern Central Atlantic Ocean,off the Coast of Ghana [J]. Marine Pollution Bulletin,2020,153,110998 .
[15] Abbasi S,Turner A. Human exposure to microplastics: A study in Iran [J]. Journal of Hazardous Materials,2021,403,123799.
[16] Flemming H C,Wingender J. The biofilm matrix [J]. Nature Reviews Microbiology,2010,8(9):623-633.
[17] Wu X J,Pan J,Li M,et al. Selective enrichment of bacterial pathogens by microplastic biofilm [J]. Water Research,2019,165,114979.
[18] Zhang X X,Zhang T,Fang H H P. Antibiotic resistance genes in water environment [J]. Applied Microbiology and Biotechnology,2009,82(3):397-414.
[19] Wang J,Qin X,Guo J,et al. Evidence of selective enrichment of bacterial assemblages and antibiotic resistant genes by microplastics in urban rivers [J]. Water Research,2020,183,116113.
[20] Galafassi S,Sabatino R,Sathicq M B,et al. Contribution of microplastic particles to the spread of resistances and pathogenic bacteria in treated wastewaters [J]. Water Research,2021,201,117368.
[21] Stevenson C,Hall J P,Harrison E,et al. Gene mobility promotes the spread of resistance in bacterial populations [J]. Isme Journal,2017,11(8):1930-1932.
[22] Liao H,Friman V P,Geisen S,et al. Horizontal gene transfer and shifts in linked bacterial community composition are associated with maintenance of antibiotic resistance genes during food waste composting [J]. Science of the Total Environment,2019,660:841-850.
[23] Ochman H,Lawrence J G,Groisman E A. Lateral gene transfer and the nature of bacterial innovation [J]. Nature,2000,405(6784):299-304.
[24] Andersson D I,Hughes D. Microbiological effects of sublethal levels of antibiotics [J]. Nature Reviews Microbiology,2014,12(7):465-478.
[25] Bai Y,Xu R,Wang Q P,et al. Sludge anaerobic digestion with high concentrations of tetracyclines and sulfonamides: Dynamics of microbial communities and change of antibiotic resistance genes [J]. Bioresource Technology,2019,276:51-59.
[26] Liu Z B,Klumper U,Liu Y,et al. Metagenomic and metatranscriptomic analyses reveal activity and hosts of antibiotic resistance genes in activated sludge [J]. Environment International,2019,129:208-220.
[27] Feng J,Li B,Jiang X,et al. Antibiotic resistome in a large-scale healthy human gut microbiota deciphered by metagenomic and network analyses [J]. Environmental Microbiology,2018,20(1):355- 368.
[28] 刘晓薇,方芳,李兰兰,等.磺胺甲恶唑胁迫对2种微塑料上细菌群落和抗性基因影响的初步研究[J]. 生态毒理学报,2022,17:1-11.
Liu X W,Fang F,Li L L,et al. Preliminary study on antibiotic resistance genes and bacterial communities on two types of microplastics under sulfamethoxazole stress [J]. Asian Journal of Ecotoxicology,2022,17:1-11.
[29] 周昕原,王言仔,苏建强,等.微塑料对河水抗生素抗性基因的影响 [J]. 环境科学,2020,41(9):4076-4080.
Zhou X Y,Wang Y Z,Su J Q,et al. Microplastics-induced shifts of diversity and abundance of antibiotic resistance genes in river water [J]. Environmental Science,2020,41(9):4076-4080.
[30] Lu X M,Lu P Z,Liu X P. Fate and abundance of antibiotic resistance genes on microplastics in facility vegetable soil [J]. Science of the Total Environment,2020,709,136276.
[31] Shi J,Wu D,Su Y,et al. Selective enrichment of antibiotic resistance genes and pathogens on polystyrene microplastics in landfill leachate [J]. Science of the Total Environment,2021,765,142775.
[32] Dai H H,Gao J F,Wang Z Q,et al. Behavior of nitrogen,phosphorus and antibiotic resistance genes under polyvinyl chloride microplastics pressures in an aerobic granular sludge system [J]. Journal of Cleaner Production,2020,256,120402.
[33] Zhao Y,Gao J,Wang Z,et al. Responses of bacterial communities and resistance genes on microplastics to antibiotics and heavy metals in sewage environment [J]. Journal of Hazardous Materials,2021,402: 123550-.
[34] Yan X,Yang X,Tang Z,et al. Downward transport of naturally-aged light microplastics in natural loamy sand and the implication to the dissemination of antibiotic resistance genes [J]. Environmental Pollution,2020,262,114270.
[35] Di Cesare A,Pinnell L J,Brambilla D,et al. Bioplastic accumulates antibiotic and metal resistance genes in coastal marine sediments [J]. Environmental Pollution,2021,291,118161.
[36] Chen G,Feng Q,Wang J. Mini-review of microplastics in the atmosphere and their risks to humans [J]. Science of the Total Environment,2020,703,135504.
[37] Vianello A,Jensen R L,Liu L,et al. Simulating human exposure to indoor airborne microplastics using a Breathing Thermal Manikin [J]. Scientific reports,2019,9(1),8670.
[38] Evangeliou N,Grythe H,Klimont Z,et al. Atmospheric transport is a major pathway of microplastics to remote regions [J]. Nature Communications,2020,11(1),3381.
[39] Li J,Cao J J,Zhu Y G,et al. Global survey of antibiotic resistance genes in air [J]. Environmental Science & Technology,2018,52(19): 10975-10984.
[40] Allen S,Allen D,Phoenix V R,et al. Atmospheric transport and deposition of microplastics in a remote mountain catchment [J]. Nature Geoscience,2019,12(5):339-344.
[41] Zhang T,Li X Y,Wang M F,et al. Time-resolved spread of antibiotic resistance genes in highly polluted air [J]. Environment International,2019,127:333-339.
[42] Zhang Q,Xu E G,Li J,et al. A review of microplastics in table salt,drinking water,and air: Direct human exposure [J]. Environmental Science & Technology,2020,54(7):3740-3751.
[43] 周 帅,李伟轩,唐振平,等.气载微塑料的赋存特征、迁移规律与毒性效应研究进展 [J]. 中国环境科学,2020,40(11):5027-5037.
Zhou S,Li W X,Tang W P,et al. Progress on the occurrence,migration and toxicity of airborne microplastics [J]. China Environmental Science,2020,40(11):5027-5037.
[44] Liu K,Wang X H,Fang T,et al. Source and potential risk assessment of suspended atmospheric microplastics in Shanghai [J]. Science of the Total Environment,2019,675:462-471.
[45] Prata J C. Airborne microplastics: Consequences to human health? [J]. Environmental Pollution,2018,234:115-126.
[46] Li J,Zhou L T,Zhang X Y,et al. Bioaerosol emissions and detection of airborne antibiotic resistance genes from a wastewater treatment plant [J]. Atmospheric Environment,2016,124:404-412.
[47] Zhang M,Zhao Y,Qin X,et al. Microplastics from mulching film is a distinct habitat for bacteria in farmland soil [J]. Science of the Total Environment,2019,688:470-478.
[48] Chen H,Wang Y,Sun X,et al. Mixing effect of polylactic acid microplastic and straw residue on soil property and ecological function [J]. Chemosphere,2020,243,125271.
[49] Koelmans A A,Bakir A,Burton G A,et al. Microplastic as a vector for chemicals in the aquatic environment: Critical review and model- supported reinterpretation of empirical studies [J]. Environmental Science & Technology,2016,50(7):3315-3326.
[50] 孙 璇,俞安琪,王学松,等.富里酸在聚苯乙烯微塑料上的吸附行为 [J]. 中国环境科学,2022,42(1):285-292.
Sun X,Yu A Q,Wang X S,et al. Adsorption behaviors of fulvic acid onto polystyrene microplastics. China Environmental Science,2022,42(1):285-292.
[51] 范秀磊,常卓恒,邹晔锋,等.可降解微塑料对铜和锌离子的吸附解吸特性 [J]. 中国环境科学,2021,41(5):2141-2150.
Fan X L,Chang Z H,Zou Y F,et al. Adsorption and desorption properties of degradable microplastic for Cu2+and Zn2+[J]. China Environmental Science,2021,41(5):2141-2150.
[52] Wang J,Liu X,Li Y,et al. Microplastics as contaminants in the soil environment: A mini-review [J]. Science of the Total Environment,2019,691:848-857.
[53] Zgurskaya H I,Nikaido H. Multidrug resistance mechanisms: drug efflux across two membranes [J]. Molecular microbiology,2000,37(2): 219-225.
[54] Tabak M,Scher K,Hartog E,et al. Effect of triclosan on Salmonella typhimurium at different growth stages and in biofilms [J]. FEMS Microbiology Letters,2007,267(2):200-206.
[55] Carey D E,Mcnamara P J. The impact of triclosan on the spread of antibiotic resistance in the environment [J]. Frontiers in Microbiology,2015,5:780.
[56] Sun J,Dai X H,Wang Q L,et al. Microplastics in wastewater treatment plants: Detection,occurrence and removal [J]. Water Research,2019,152:21-37.
[57] Raju S,Carbery M,Kuttykattil A,et al. Transport and fate of microplastics in wastewater treatment plants: implications to environmental health [J]. Reviews in Environmental Science and Bio-Technology,2018,17(4):637-653.
[58] Zhang Z Q,Chen Y G. Effects of microplastics on wastewater and sewage sludge treatment and their removal: A review [J]. Chemical Engineering Journal,2020,382,122955.
[59] Rolsky C,Kelkar V,Driver E,et al. Municipal sewage sludge as a source of microplastics in the environment [J]. Current Opinion in Environmental Science & Health,2020,14:16-22.
[60] Zhao Y F,Gao J F,Wang Z Q,et al. Responses of bacterial communities and resistance genes on microplastics to antibiotics and heavy metals in sewage environment [J]. Journal of Hazardous Materials,2021,402,123550.
[61] Shi J H,Wu D,Su Y L,et al. (Nano)microplastics promote the propagation of antibiotic resistance genes in landfill leachate [J]. Environmental Science-Nano,2020,7(11):3536-3546.
[62] Eckert E M,Di Cesare A,Kettner M T,et al. Microplastics increase impact of treated wastewater on freshwater microbial community [J]. Environmental Pollution,2018,234:495-502.
[63] 王佳佳,赵娜娜,李金惠.中国海洋微塑料污染现状与防治建议 [J]. 中国环境科学,2019,39(7):3056-3063.
Wang J J,Zhao N N,Li J H,et al. Current situation of marine microplastics pollution and prevention proposals in China [J]. China Environmental Science,2019,39(7):3056-3063.
[64] Ivar Do Sul J A,Costa M F. The present and future of microplastic pollution in the marine environment [J]. Environmental Pollution,2014,185:352-364.
[65] Zhao S,Zhu L,Li D. Microplastic in three urban estuaries,China [J]. Environmental Pollution,2015,206:597-604.
[66] 徐 沛,彭谷雨,朱礼鑫,等.长江口微塑料时空分布及风险评价 [J]. 中国环境科学,2019,39(5):2071-2077.
Xu P,Peng G Y,Zhu L X,et al. Spatial-temporal distribution and pollution load of microplastics in the Changjiang Estuary [J]. China Environmental Science,2019,39(5):2071-2077.
[67] 王志超,窦雅娇,周 鑫,等.冰封期岱海微塑料赋存与环境因子的关系及风险评价 [J]. 中国环境科学,2022,42(2):889-896.
Wang Z C,Dou Y J,Zhou X,et al. Relationship between microplastics occurrence and environmental factors and risk assessment during ice-covered period of the Daihai Lake [J]. China Environmental Science,2022,42(2):889-896.
[68] Jiang Y,Yang F,Zhao Y,et al. Greenland Sea Gyre increases microplastic pollution in the surface waters of the Nordic Seas [J]. Science of the Total Environment,2020,712,136484.
[69] Lu J,Zhang Y,Wu J,et al. Effects of microplastics on distribution of antibiotic resistance genes in recirculating aquaculture system [J]. Ecotoxicology and Environmental Safety,2019,184,109631.
[70] Yang Y Y,Liu G H,Song W J,et al. Plastics in the marine environment are reservoirs for antibiotic and metal resistance genes [J]. Environment International,2019,123:79-86.
[71] Guo X P,Sun X L,Chen Y R,et al. Antibiotic resistance genes in biofilms on plastic wastes in an estuarine environment [J]. Science of the Total Environment,2020,745,140916.
[72] Wang S,Xue N,Li W,et al. Selectively enrichment of antibiotics and ARGs by microplastics in river,estuary and marine waters [J]. Science of the Total Environment,2020,708,134594.
[73] Van Cauwenberghe L,Devriese L,Galgani F,et al. Microplastics in sediments: A review of techniques,occurrence and effects [J]. Marine Environmental Research,2015,111:5-17.
[74] Zhang Y X,Lu J,Wu J,et al. Potential risks of microplastics combined with superbugs: Enrichment of antibiotic resistant bacteria on the surface of microplastics in mariculture system [J]. Ecotoxicology and Environmental Safety,2020,187,109852.
[75] Amaral-Zettler L A,Zettler E R,Mincer T J. Ecology of the plastisphere [J]. Nature Reviews Microbiology,2020,18(3):139-151.
[76] Zettler E R,Mincer T J,Amaral-Zettler L A. Life in the "Plastisphere": Microbial communities on plastic marine debris [J]. Environmental Science & Technology,2013,47(13):7137-7146.
[77] Mac Aogain M,Lau K J X,Cai Z,et al. Metagenomics reveals a core macrolide resistome related to microbiota in chronic respiratory disease [J]. American Journal of Respiratory and Critical Care Medicine,2020,202(3):433-447.
[78] Munck C,Albertsen M,Telke A,et al. Limited dissemination of the wastewater treatment plant core resistome [J]. Nature Communications,2015,6,8452.
[79] Lau K J,Mac Aogain M,Narayana J K,et al. Functional metagenomics reveals a core resistome and potential resistance reservoirs in chronic respiratory disease [J]. American Journal of Respiratory and Critical Care Medicine,2020,202(3):433–447.
[80] Du S,Shen J P,Hu H W,et al. Large-scale patterns of soil antibiotic resistome in Chinese croplands [J]. Science of the Total Environment,2020,712,136418.
[81] Percival S L,Knapp J S,Edyvean R,et al. Biofilm development on stainless steel in mains water [J]. Water Research,1998,32(1):243- 253.
[82] Chang Y,Yandi W,Chen W Y,et al. Tunable bioadhesive copolymer hydrogels of thermoresponsive poly(n-isopropyl acrylamide) containing zwitterionic polysulfobetaine [J]. Biomacromolecules,2010,11(4):1101-1110.
[83] Cheng Y F,Feng G P,Moraru C I. Micro- and nanotopography sensitive bacterial attachment mechanisms: A review [J]. Frontiers in microbiology,2019,10,191.
[84] Allison D G. The biofilm matrix [J]. Biofouling,2003,19(2):139-150.
[85] O'toole G,Kaplan H B,Kolter R. Biofilm formation as microbial development [J]. Annual Review of Microbiology,2000,54:49-79.
[86] Nguyen H,Ybarra A,Basagaoglu H,et al. Biofilm viscoelasticity and nutrient source location control biofilm growth rate,migration rate,and morphology in shear flow [J]. Scientific reports,2021,11(1): 16118.
[87] Zhang W,Sileika T S,Chen C,et al. A novel planar flow cell for studies of biofilm heterogeneity and flow-biofilm interactions [J]. Biotechnology and Bioengineering,2011,108(11):2571-2582.
[88] Paramonova E,Kalmykowa O J,Van Der Mei H C,et al. Impact of hydrodynamics on oral biofilm strength [J]. Journal of Dental Research,2009,88(10):922-926.
[89] Liu Y,Tay J H. Metabolic response of biofilm to shear stress in fixed-film culture [J]. Journal of Applied Microbiology,2001,90(3): 337-342.
[90] Rao T S,Rani P G,Venugopalan V P,et al. Biofilm formation in a freshwater environment under photic and aphotic conditions [J]. Biofouling,1997,11(4):265-282.
[91] Hostacka A,Ciznar I,Stefkovicova M. Temperature and pH affect the production of bacterial biofilm [J]. Folia Microbiologica,2010,55(1): 75-78.
[92] Su Y L,Zhang Z J,Zhu J D,et al. Microplastics act as vectors for antibiotic resistance genes in landfill leachate: The enhanced roles of the long-term aging process [J]. Environmental Pollution,2021,270,116278.
[93] Guo X P,Sun X L,Chen Y R,et al. Antibiotic resistance genes in biofilms on plastic wastes in an estuarine environment [J]. Science of the Total Environment,2020,745,140916.
[94] Parrish K,Fahrenfeld N L. Microplastic biofilm in fresh- and wastewater as a function of microparticle type and size class [J]. Environmental Science-Water Research & Technology,2019,5(3): 495-505.
[95] Guo X,Liu Y,Wang J. Sorption of sulfamethazine onto different types of microplastics: A combined experimental and molecular dynamics simulation study [J]. Marine Pollution Bulletin,2019,145:547-554.
[96] Li X,Mei Q,Chen L,et al. Enhancement in adsorption potential of microplastics in sewage sludge for metal pollutants after the wastewater treatment process [J]. Water Research,2019,157:228-237.
[97] Imran M,Das K R,Naik M M. Co-selection of multi-antibiotic resistance in bacterial pathogens in metal and microplastic contaminated environments: An emerging health threat [J]. Chemosphere,2019,215:846-857.
[98] Lagarde F,Olivier O,Zanella M,et al. Microplastic interactions with freshwater microalgae: Hetero-aggregation and changes in plastic density appear strongly dependent on polymer type [J]. Environmental Pollution,2016,215:331-339.
[99] Lagana P,Caruso G,Corsi I,et al. Do plastics serve as a possible vector for the spread of antibiotic resistance? First insights from bacteria associated to a polystyrene piece from King George Island (Antarctica) [J]. International Journal of Hygiene and Environmental Health,2019,222(1):89-100.
[100] Flach C F,Pal C,Svensson C J,et al. Does antifouling paint select for antibiotic resistance? [J]. Science of the Total Environment,2017,590:461-468.
[101] Lu X M,Lu P Z,Liu X P. Fate and abundance of antibiotic resistance genes on microplastics in facility vegetable soil [J]. Science of the Total Environment,2020,709,136276.
[102] Andrady A L. Microplastics in the marine environment [J]. Marine Pollution Bulletin,2011,62(8):1596-1605.
[103] Bergmann M,Mutzel S,Primpke S,et al. White and wonderful? Microplastics prevail in snow from the Alps to the Arctic [J]. Science Advances,2019,5(8):eaax1157.
[104] Klein M,Fischer E K. Microplastic abundance in atmospheric deposition within the Metropolitan area of Hamburg,Germany [J]. Science of the Total Environment,2019,685:96-103.
[105] Schmidt C,Krauth T,Wagner S. Export of plastic debris by rivers into the sea [J]. Environmental Science & Technology,2017,51(21): 12246-12253.
[106] Zhang Y,Lu J,Wu J,et al. Potential risks of microplastics combined with superbugs: Enrichment of antibiotic resistant bacteria on the surface of microplastics in mariculture system [J]. Ecotoxicology and Environmental Safety,2020,187,109852.
[107] Arias-Andres M,Kluemper U,Rojas-Jimenez K,et al. Microplastic pollution increases gene exchange in aquatic ecosystems [J]. Environmental Pollution,2018,237:253-261.
[108] Schwarz A E,Ligthart T N,Boukris E,et al. Sources,transport,and accumulation of different types of plastic litter in aquatic environments: A review study [J]. Marine Pollution Bulletin,2019,143:92-100.
[109] Pinilla-Redondo R,Cyriaque V,Jacquiod S,et al. Monitoring plasmid-mediated horizontal gene transfer in microbiomes: Recent advances and future perspectives [J]. Plasmid,2018,99:56-67.
[110] Cheng D,Ngo H H,Guo W,et al. Contribution of antibiotics to the fate of antibiotic resistance genes in anaerobic treatment processes of swine wastewater: A review [J]. Bioresource Technology,2020,299,122654.
[111] Luo Y,Wang Q,Lu Q,et al. An ionic liquid facilitates the proliferation of antibiotic resistance genes mediated by class I integrons [J]. Environmental Science & Technology Letters,2014,1(5):266-270.
[112] Li P H,Zou X Y,Wang X D,et al. A preliminary study of the interactions between microplastics and citrate-coated silver nanoparticles in aquatic environments [J]. Journal of Hazardous Materials,2020,385,121601.
[113] Xiong Y C,Zhao J H,Li L Q,et al. Interfacial interaction between micro/nanoplastics and typical PPCPs and nanoplastics removal via electrosorption from an aqueous solution [J]. Water Research,2020,184,116100.
[114] Qiu Z,Yu Y,Chen Z,et al. Nanoalumina promotes the horizontal transfer of multiresistance genes mediated by plasmids across genera [J]. Proceedings of the National Academy of Sciences of the United States of America,2012,109(13):4944-4949.
[115] Zou W,Li X,Lai Z,et al. Graphene oxide inhibits antibiotic uptake and antibiotic resistance gene propagation [J]. Acs Applied Materials & Interfaces,2016,8(48):33165-33174.
[116] Wang X,Yang F,Zhao J,et al. Bacterial exposure to ZnO nanoparticles facilitates horizontal transfer of antibiotic resistance genes [J]. Nanoimpact,2018,10:61-67.
[117] Guo M T,Tian X B. Impacts on antibiotic-resistant bacteria and their horizontal gene transfer by graphene-based TiO2&Ag composite photocatalysts under solar irradiation [J]. Journal of Hazardous Materials,2019,380,120877.
[118] Zhang S,Wang Y,Song H,et al. Copper nanoparticles and copper ions promote horizontal transfer of plasmid-mediated multi-antibiotic resistance genes across bacterial genera [J]. Environment International,2019,129:478-487.
[119] Lu J,Wang Y,Jin M,et al. Both silver ions and silver nanoparticles facilitate the horizontal transfer of plasmid-mediated antibiotic resistance genes [J]. Water Research,2020,169,115229.
[120] Karygianni L,Ren Z,Koo H,et al. Biofilm matrixome: Extracellular components in structured microbial communities [J]. Trends in Microbiology,2020,28(8):668-681.
[121] Tsuneda S,Aikawa H,Hayashi H,et al. Extracellular polymeric substances responsible for bacterial adhesion onto solid surface [J]. FEMS Microbiology Letters,2003,223(2):287-292.
[122] Han P P,Shen S G,Guo R J,et al. ROS is a factor regulating the increased polysaccharide production by light quality in the edible cyanobacterium nostoc flagelliforme [J]. Journal of Agricultural and Food Chemistry,2019,67(8):2235-2244.
[123] Li G,Chen X,Yin H,et al. Natural sphalerite nanoparticles can accelerate horizontal transfer of plasmid-mediated antibiotic- resistance genes [J]. Environment International,2020,136,105497.
[124] Wang Y,Lu J,Engelstadter J,et al. Non-antibiotic pharmaceuticals enhance the transmission of exogenous antibiotic resistance genes through bacterial transformation [J]. Isme Journal,2020,14(8):2179- 2196.
[125] Van Acker H,Coenye T. The role of reactive oxygen species in antibiotic-mediated killing of bacteria [J]. Trends in Microbiology,2017,25(6):456-466.
[126] Han X,Lv P,Wang L G,et al. Impact of nano-TiO2on horizontal transfer of resistance genes mediated by filamentous phage transduction [J]. Environmental Science-Nano,2020,7(4):1214-1224.
[127] Wang Q,Mao D,Luo Y. Ionic liquid facilitates the conjugative transfer of antibiotic resistance genes mediated by plasmid RP4 [J]. Environmental Science & Technology,2015,49(14):8731-8740.
[128] Papenfort K,Bassler B L. Quorum sensing signal-response systems in Gram-negative bacteria [J]. Nature Reviews Microbiology,2016,14(9): 576-588.
[129] Lu Y,Zeng J,Wu B,et al. Quorum sensing N-acyl homoserine lactones-sdia suppresses escherichia coli-pseudomonas aeruginosa conjugation through inhibiting tral expression [J]. Frontiers in Cellular and Infection Microbiology,2017,7:7.
[130] Zheng J,Chen T,Chen H. Antibiotic resistome promotion in drinking water during biological activated carbon treatment: Is it influenced by quorum sensing? [J]. Science of the Total Environment,2018,612:1-8.
Effects of microplastics on the spread and horizontal gene transfer of antibiotic resistance genes.
WU Xiao-jian1,2,3,SHI Guo-jing1,2,3,WANG Ying-ying1,2,3*
(1.College of Environmental Science and Engineering,Nankai University,Tianjin 300350,China;2.Key Laboratory Processes and Environmental Criteria (Ministry of Education),Tianjin 300350,China;3.Tianjin Key Laboratory of Environmental Remediation and Pollution Control,Tianjin 300350,China).,2022,42(8):3957~3968
As an emerging pollutant,microplastics are ubiquitous in different types of environments. The antibiotic resistance genes (ARGs) carried by microplastic biofilms have adverse effects on human health,which has aroused wide concerns. However,the role of microplastics in the process of long migration and horizontal gene transfer of ARGs is still not clear. The environmental impacts of the resistance dissemination caused by microplastics also need further investigation. The current status of microplastic pollutants and the ARGs they carry was summarized. The pathways by which microplastics spread ARGs and the regulation of ARG transfer via microplastics are also compared and discussed. Future research perspectives are proposed to provide possible approaches for solving the problem of antibiotic resistance dissemination by microplastics.
microplastic;plastic;antibiotic resistance genes;plastisphere;pollution characteristics
X171.5
A
1000-6923(2022)08-3957-12
2022-01-11
国家自然科学基金委面上项目(31870485);天津市自然科学基金项目(19JCZDJC39600);天津市研究生科研创新项目(2019YJSB052)
* 责任作者,教授,wangyy@nankai.edu.cn
吴效俭(1995-),女,天津人,南开大学博士研究生.主要从事抗性基因的宏基因组、宏转录组检测与定量分析相关研究.发表论文2篇.
猜你喜欢
现代临床医学(2021年5期)2021-11-02 05:21:18
昆明医科大学学报(2021年4期)2021-07-23 01:22:08
空间科学学报(2021年1期)2021-05-22 01:36:34
环境保护与循环经济(2017年5期)2018-01-22 02:56:44
Coco薇(2016年8期)2016-10-09 16:58:11
中国果菜(2016年9期)2016-03-01 01:28:44
中国蔬菜(2015年9期)2015-12-21 13:04:40
少儿科学周刊·少年版(2015年3期)2015-07-07 20:57:25
中华皮肤科杂志(2014年4期)2014-12-19 12:55:59
西安建筑科技大学学报(自然科学版)(2014年4期)2014-11-12 05:15:34
