
齐口裂腹鱼上溯洄游过程中的感官圈解译研究
2022-08-23 10:11邓月曦唐锡良严忠銮王小明卢晶莹安瑞冬
中国环境科学 2022年8期
邓月曦,唐锡良,严忠銮,王小明,卢晶莹,安瑞冬,李 嘉*
齐口裂腹鱼上溯洄游过程中的感官圈解译研究
邓月曦1,唐锡良2,严忠銮2,王小明2,卢晶莹3,安瑞冬1,李 嘉1*
(1.四川大学水利水电学院,水力学与山区河流开发保护国家重点实验室,四川 成都 610065;2.中国三峡建工(集团)有限公司,四川 成都 610041;3.中国长江三峡集团有限公司,湖北 武汉 430010)
选取西南地区典型鱼类齐口裂腹鱼(,(32.32±2.73)cm)作为实验对象,测试鱼类在上溯过程中应对不同水流环境时的游泳行为,解译不同大小网格尺寸下鱼类的游泳速度、转向次数以及转角和转角差与真实游泳轨迹的匹配程度.结果表明:采用小于或等于0.30m尺寸的网格时,分析得出的鱼类上溯运动平均游泳速度、转向次数以及转角和转角差均与实际情况无显著性差异,另外鱼类更偏向于在0.20~0.40m/s的流速区间内以和水平方向呈-49º~49º的运动搜索范围进行上溯运动.
网格尺寸;生态水流;鱼类行为;感官圈
解析鱼类上溯洄游过程中应对复杂流场的行为响应,如趋流行为及逃逸行为等[1-2],是优化过鱼设施设计[3]、保护洄游性鱼类及恢复河流天然属性的重要研究内容[4-5].基于鱼类个体行为学与水动力学耦合的数学模型为模拟洄游性鱼类通过自然水流障碍或水工建筑物提供了理论支撑,同时也为生物粒子模拟构建提供了数据支持[6].鱼的侧线系统能够获取鱼体周围的水流信号,评估游泳区域内的水动力条件,进而作出相应的行为反应[7]. Goodwin等[8]基于此提出了感官圈的概念,即鱼类的上溯洄游轨迹模拟定义为以感官圈的形式进行搜索上溯.感官圈特征主要包括网格尺寸划分、水动力条件趋向性、感官搜索范围、感官搜索角度4个部分,鱼类在合适的网格单元内以感官搜索范围在搜索角度范围内对水动力条件进行分析,选择偏好的水力条件进行上溯.其中感官搜索范围(SQD)选择生物响应范围和CFD模型搜索范围中的较大值,它与鱼体体长呈正相关关系,体长越长的鱼能够检索到更大范围内的水动力学条件[9].Gao等[10]建立了欧拉-拉格朗日鱼类运动模型,但该模型仅考虑湍流动能作为水力刺激因子运用感官圈进行鱼类的上溯模拟.Tan等[11]进一步建立考虑湍流动能、应变率和速度等多种水力刺激的鱼类运动轨迹模型,模拟获得鲢鱼的水力刺激范围.Zielinski等[12]考虑了感官搜索角度的范围,假设鱼只被刺激向上游动,即鱼不回溯,以鱼位置上游180°的半圆弧内进行网格节点的搜索,以减少模拟计算量.Zhu等[13]将感官圈的概念运用于生物颗粒模型来预测大坝下鱼的上溯轨迹,生物颗粒模型将鱼视为颗粒,并将流量、速度梯度和湍流动能作为回溯行为的条件,利用控制方程简化鱼的行为,开发鱼的可追溯预测程序.
然而上述方法并没有考虑到网格尺寸的大小对鱼类上溯运动感官圈解译的影响,网格尺寸过大时无法准确解译鱼类有效游泳行为信息,导致模拟结果与真实行为差异较大,网格尺寸过小时虽能捕捉丰富行为信息,但在大尺度空间中使用时会使得模型计算时间长,效率低下,合理的网格尺寸大小有利于精准、有效的解译鱼类上溯行为.另一方面,目前在模拟鱼类上溯运动时常对鱼类感官圈定义为360°整圆方向,但鱼类的实际运动有可能是呈现的扇形搜索范围,进一步确定搜索范围的区间可以减少预测鱼类轨迹时的计算量.据此本文研究了不同网格尺寸划分对齐口裂腹鱼上溯轨迹模拟准确程度的影响,选定合适的网格尺寸来反映鱼类上溯行为,此外,对齐口裂腹鱼在上溯运动过程中的响应流速、感官搜索角度的范围进行进一步探索.本研究为鱼类上溯应对湍流环境时的运动轨迹模拟网格尺寸选取提供了一种研究思路,同时也为生物粒子模拟构建提供了数据支撑.
1 材料与方法
1.1 研究对象
齐口裂腹鱼()属于鲤科、裂腹鱼属的底栖鱼类,喜在较低的水温环境中活动,一般生活于急缓流交界处,有短距离的河川生殖洄游需求[14-15].实验所用齐口裂腹鱼来自四川水产研究所于2020年10月网捕的成熟鱼种,平均体长(32.32±2.73)cm,体重(304.21±77.15)g.实验过程中随机挑选生理活性较好的齐口裂腹鱼作为实验对象,为避免鱼类环境适应性影响实验结果的可靠性,实验鱼不重复使用.
1.2 实验装置
本研究在四川大学水力学与山区河流开发保护国家重点实验室自制的大型开放式浅水循环水槽(图1)内进行.水槽由钢化玻璃和钢架构造,长20.50m、宽4.00m、高1.00m.循环装置由4个不同的功能部分组成:上游蓄水池、适应区域、下游蓄水池和循环供水渠道.水槽的边墙以及底板均为光滑直线型,以减少流动摩擦和边界层效应.
在上游3个等宽(1.00m)入口处通过进口格栅将游泳测试区域水流分成低、中和高流速区3种流速大小不同的区域,模拟不同水流环境条件下的鱼类上溯运动.
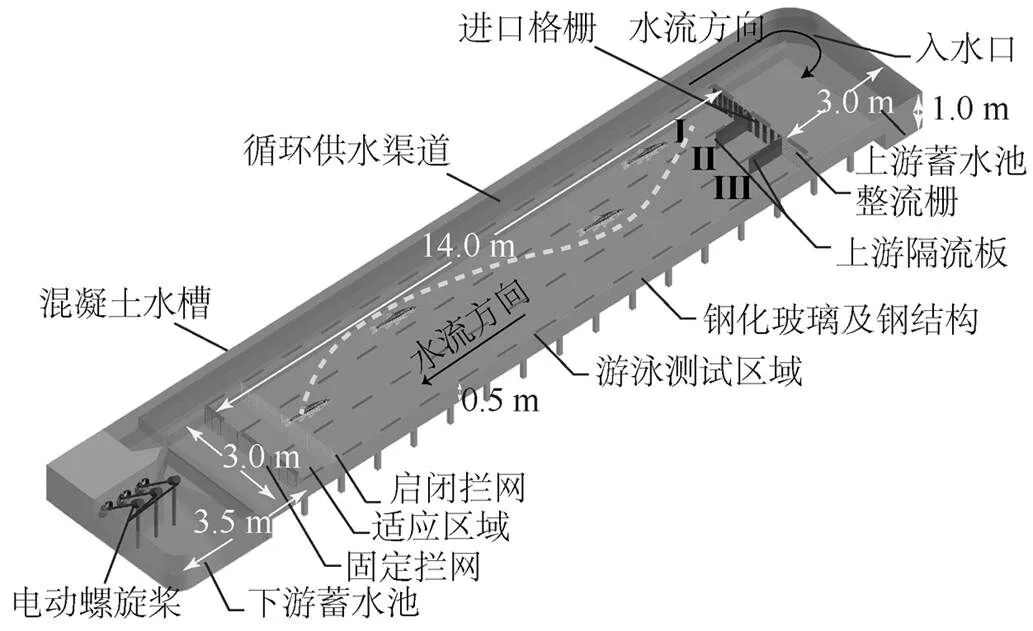
图1 鱼类上溯行为研究实验装置几何示意
1.3 实验方法
每次试验开始时,在适应区域放入1条健康的个体鱼,以适应水流温度和流速条件.适应0.50h后,抬升启闭拦网,让试验鱼在游泳测试区域自主游泳,同时使用4台高清摄像机(WIM SkyStar,30fps)连续记录鱼类上溯洄游游泳行为.试验结束条件为当测试鱼通过整个游泳测试区域并最终进入上游进口,或者测试鱼在1.00h内从未离开过适应区域,测试结束.
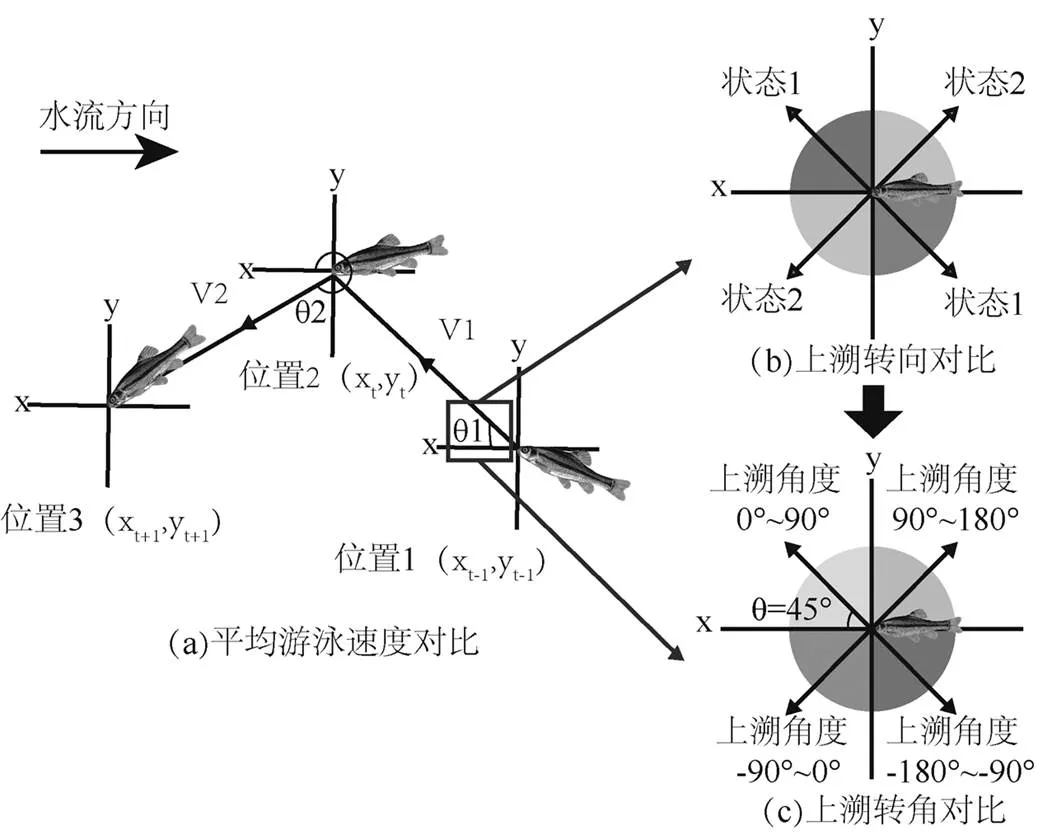
图2 实验方法示意
鱼的运动方向定义为深色和浅色两种状态方向,若运动方向出现颜色的转变,则判定为转向
本研究选择50尾齐口裂腹鱼进行实验,实验期间禁止在水槽周围活动,避免声音对实验鱼的干扰.后期通过LoggerPro视频处理软件以1/30(s)的帧率记录鱼的位置坐标数据.将鱼的轨迹点分别导入网格大小为0.20m×0.20m、0.25m×0.25m、0.30m× 0.30m、0.40m×0.40m、0.50m×0.50m、1.00m×1.00m的网格中进行分析.如图2所示,通过对比鱼类上溯运动平均游泳速度、转向次数以及转角和转角差得到合理的齐口裂腹鱼上溯运动网格分析尺寸.
此外,实验水槽的水流速度通过3D声学多普勒测速仪(ADV)进行测量,研究齐口裂腹鱼在上溯运动过程中对流速的偏好性,同时探究上溯搜索角度有无特定范围.
1.4 数据分析
为研究不同网格尺寸下鱼类游泳行为信息的连续及缺失程度,本研究采用游泳轨迹连续性、鱼类平均游泳速度、上溯轨迹转向、上溯轨迹转角及转角差5类指标对网格有效性进行评估.其中相邻轨迹点之间的鱼类平均速度如式(1)所示:

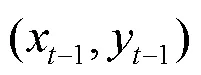

图2c中给出了转角的定义,转角是以横坐标的正方向作为参考方向的,为利于研究统计,转角范围以[0º,360º]表示;转角差则定义为相邻两次转角之间的差值,其中顺时针方向为正(+),称为右旋,反之为负(-),称为左旋.
转角差Δ的计算方法如下式:



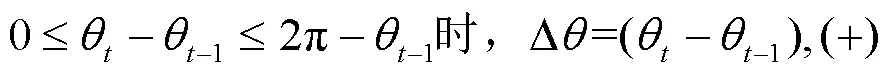

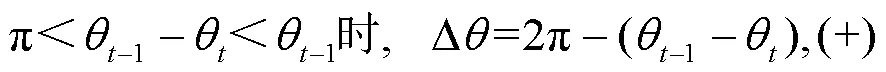
采用SPSS 20.0进行相关数据统计分析,不同网格尺寸下鱼的平均游泳速度采用单因素方差(ANOVA)分析.统计值均以平均值±标准误(Mean±SE)表示,显著水平为<0.05.
2 结果与分析
2.1 确定适宜网格尺寸
2.1.1 轨迹连续性 对鱼类运动的研究表明,当测试鱼进入试验水体环境时,它很快就能找到自己喜好的水深(或动水压力),然后在同一水层中游动.事实上,Rodriguez等[16]记录的真实鱼的轨迹也是在平面上呈现的,即二维上的,因此研究可忽略垂直上的变化.本文采用二维水流速度进行分析(图3),LoggerPro处理得到的单尾齐口裂腹鱼游泳速度区间为=1.47m/s左右,参考库朗数(o)的定义[17],当时间步长为1秒时,网格的大小不应大于×Δ,即不应大于1.47m.因此确定研究网格尺寸为0.20,0.25,0.30,0.40,0.50,1.00m.
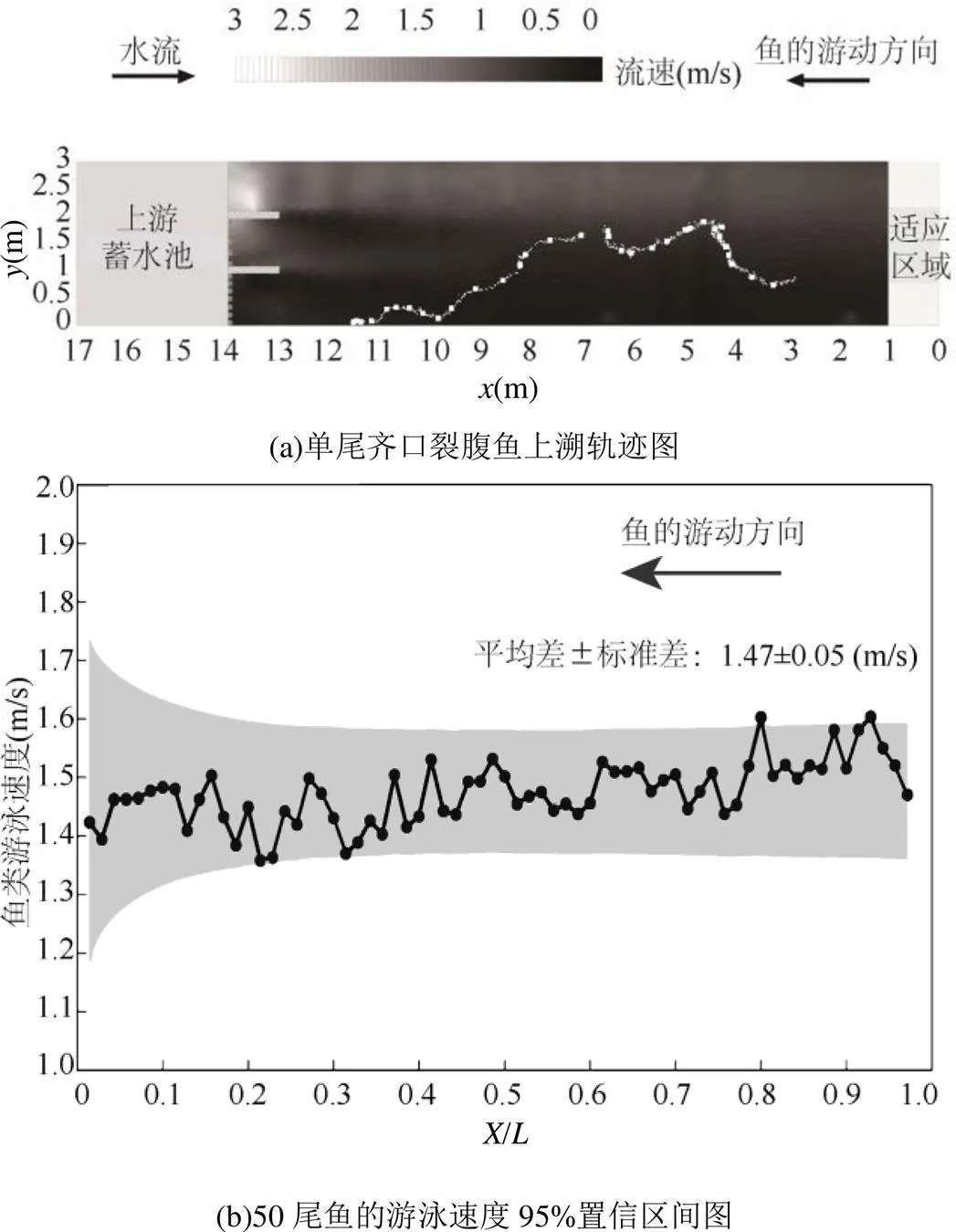
图3 鱼类上溯轨迹图及游泳速度置信区间图
i>X/表示鱼类运动坐标与测试长度范围(13m)的比值
将齐口裂腹鱼的轨迹点导入不同尺寸大小的网格进行筛选,导入轨迹点圆圈的直径为鱼的平均体长32.32cm,采用计算流体力学的定义,圈内不包含有网格节点的即视为不能被网格识别,需删除该轨迹点.最终得到的鱼类上溯轨迹点如图4所示,当网格尺寸小于0.30m时,轨迹点连续;当尺寸为0.40m时,轨迹点图出现部分断裂;而当尺寸大于0.40m时,图像无法完整地描述鱼类的上溯运动过程.
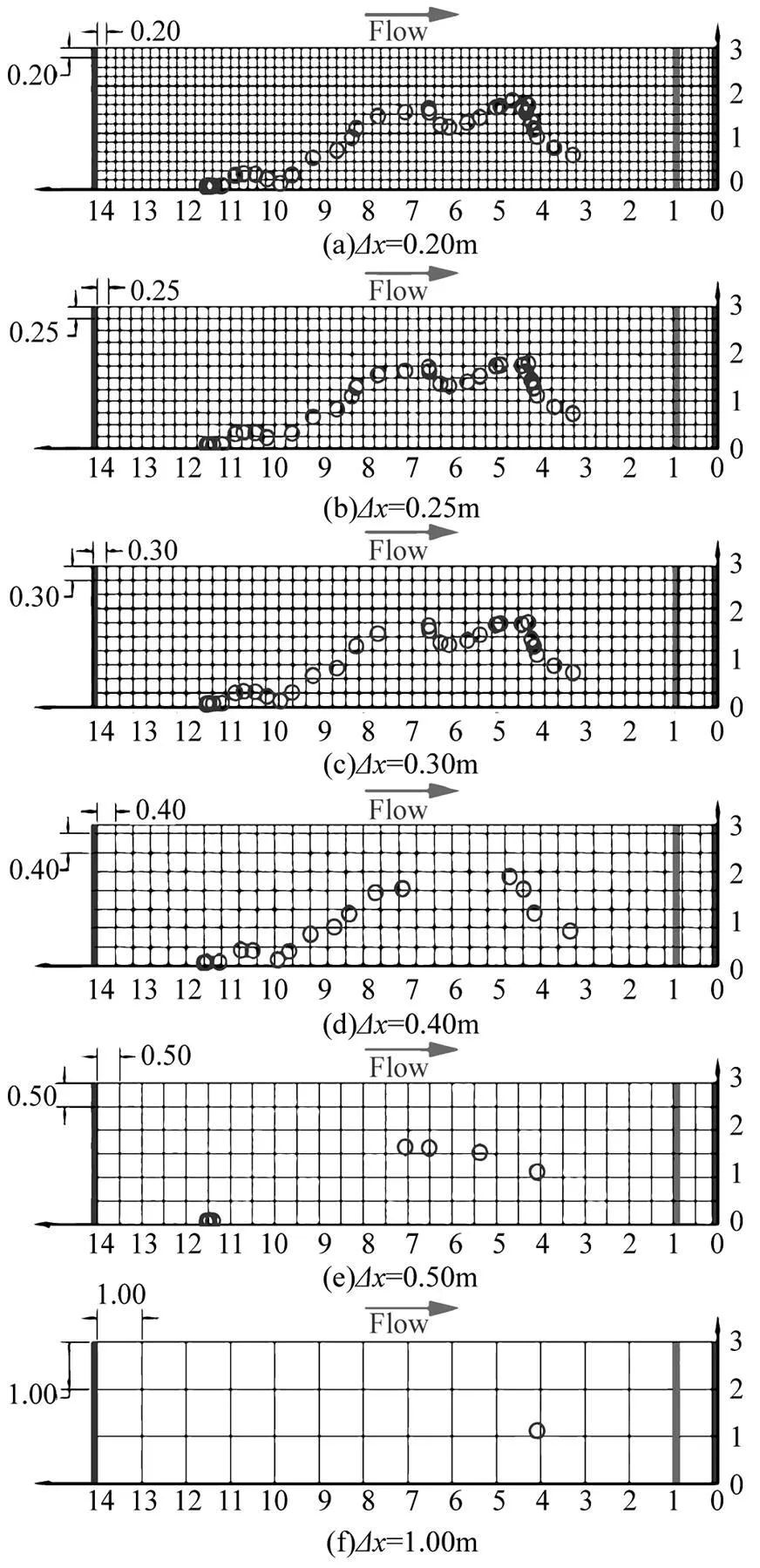
图4 不同网格尺寸下单尾鱼游泳轨迹解译图
D表示网格尺寸
2.1.2 鱼类平均速度对比 通过筛选出的各点以及记录的间隔时间计算鱼的平均游泳速度,结果如图5所示,采用小于0.30m的网格尺寸分析齐口裂腹鱼的平均游泳速度与不进行网格划分时没有显著性差异(>0.05);采用0.40,0.50,1.00m的网格尺寸计算结果彼此之间均具有显著性差异(0.05),且与真实数据也具有显著性差异(0.05).表明当网格尺寸大于0.30m时,对齐口裂腹鱼的上溯运动描述已经出现偏差.

图5 50尾鱼的游泳速度显著性差异分析
2.1.3 上溯轨迹转向对比 以1尾鱼为例,将筛选出的鱼的运动轨迹点依次连线,当网格尺寸为0.25m时(图6a),齐口裂腹鱼在上溯过程中共计转向9次,而网格尺寸增大到0.50m时(图6b),鱼共计转向3次,当转向次数明显减少时,鱼的运动轨迹不连贯,轨迹点不能正确描述上溯运动状态.统计50尾鱼上溯过程的转向次数,如图6c所示,当网格尺寸小于0.30m时,鱼的运动转向次数与实际相似,但网格尺寸大于0.30m时转向次数明显减少、异常值增多,与实际情况不符,不能正确反应鱼的上溯运动轨迹特征.
2.1.4 上溯轨迹转角及转角差对比 由图7a、b可见,以1尾鱼为例,当网格尺寸由0.25m变为0.50m时,齐口裂腹鱼上溯过程中的转角次数明显减小,转角大小也发生较大转变,因此可以通过统计鱼类转角的次数分布和数目,判别上溯运动轨迹的真实性.
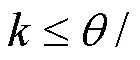

图6 鱼类上溯转向统计
如图8所示,鱼类转角的分布规律与网格尺寸的大小无关,上溯轨迹左旋与右旋大约平分,右旋略大于左旋.网格尺寸为0.20和0.25m时齐口裂腹鱼上溯运动的转角、转角差次数与真实数据情况极为相似,网格尺寸为0.30m时转角、转角差次数略小于真实情况,而网格尺寸为0.40和0.50m时转角、转角差次数减少了二分之一,1.00m时转角、转角差次数更是减少了四分之一,不能准确描述到上溯运动轨迹.
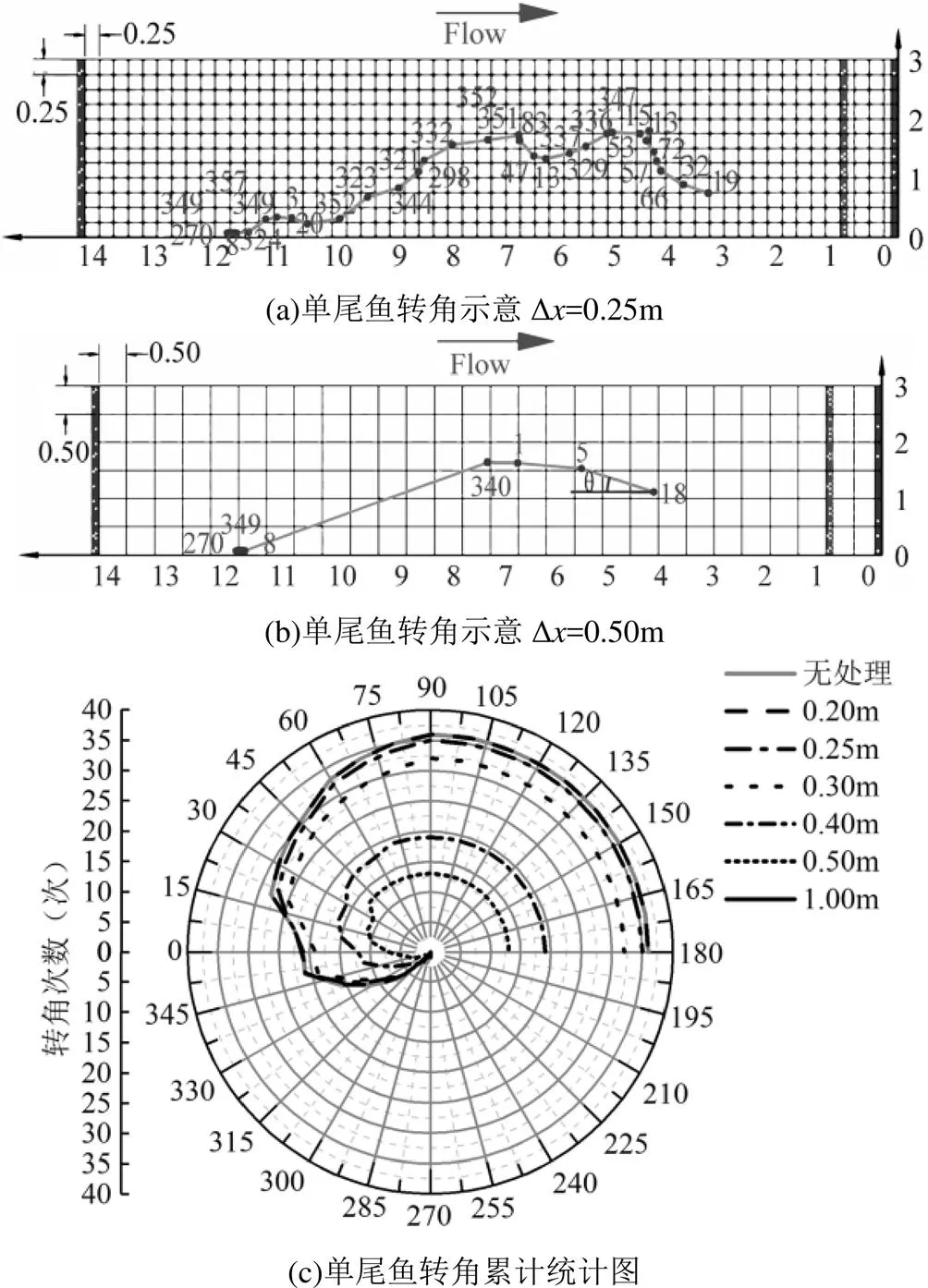
图7 单尾鱼转角示意
2.2 流速趋向性
上述研究表明,0.30m是网格尺寸分析的合理界限,因此选取此网格下的轨迹点,以1s为时间步长,分析鱼类上溯运动的流速趋向性以及上溯运动角度搜索范围.
本文使用声学多普勒测速仪提取流场的流速分布,将50尾齐口裂腹鱼的轨迹点对应水流流速通过双线性插值法计算得出,统计不同流速对应的出现相对频率如图9所示.齐口裂腹鱼对于流场流速的选择具有一定的趋向性,它更偏向于0.20~0.40m/s的流速区间内进行上溯运动.

图9 50尾鱼流速趋向
2.3 感官上溯搜索角度范围
以10°作为统计离散程度,统计鱼类上溯运动时不同转角范围的次数,如图10所示,50尾鱼共计转角4423次,取75%的置信区间,鱼的搜索角度范围是-49°~49°.因此,认为齐口裂腹鱼偏好以与水平方向呈98°的扇形角度搜索范围进行上溯运动的.
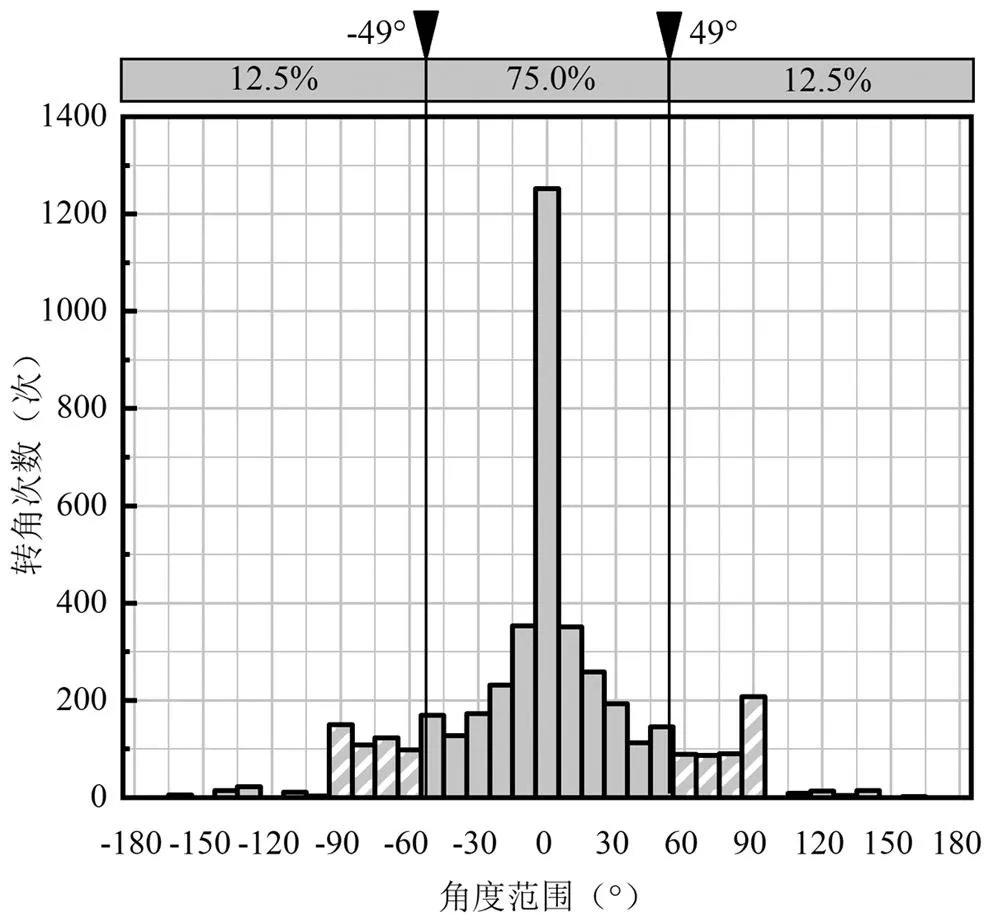
图10 50尾鱼感官上溯搜索角度
3 讨论及展望
鱼类的运动在时空上具有离散个体的差异性,其游泳轨迹的模拟研究大致可分为两个过程,首先是感知并评估感官圈内的各种水动力学因子[19],然后根据反馈的信息作出上溯方向的决策.如Liao等[20]根据细鳞裂腹鱼、圆口铜鱼和长薄鳅3种鱼类游泳速度与游泳行为之间的关系,提出了6种对应不同游泳行为策略的流速范围区间,基于此构建了与鱼类游泳特征相对应的三维游泳轨迹模型;蒋永强等[21]综合分析流速、湍流动能和湍动耗散率对两种不同体长鲑鱼上溯的影响,并依据上溯鱼类游泳速度与周围水力因子的相关性拟定函数关系,从而定量计算鱼类上溯轨迹,并发现湍流动能的影响最为显著,湍动能耗散率影响最小;Tan等[11]利用基于个体的IBM模型来模拟鲢鱼运动轨迹,该模型综合了鱼类响应的水力刺激因子(湍流动能、速度和应变率)以及鱼类个体运动的规则;Arenas等[22]则进一步指出鲑鱼可以响应除流速外的更多水力刺激因子,如温度、偏振光、地球磁场、存在的障碍等.本研究齐口裂腹鱼上溯应对复杂紊流时网格尺寸划分对轨迹解译的影响结果表明当时间间隔为1s,采用小于或等于0.30m尺寸的网格时,分析得出的鱼类上溯运动平均游泳速度、转向次数以及转角和转角差均与实际情况无显著性差异,而网格尺寸大于0.30m时鱼类游泳行为与实际运动轨迹有显著性差异,因此0.30m的网格尺寸可以较合适地反应鱼类上溯行为,同时这一网格尺寸大小也最接近鱼的体长.本研究以体长范围为(32.32±2.73)cm,体重范围为(304.21±77.15)g的齐口裂腹鱼为研究对象得出了与体长范围相关的适宜感官圈范围.由于同种鱼处于不同生长阶段时具有不同游泳行为策略,如转向角度及游泳能力等具有差异性[23-24],故研究结果可为体长相近范围内的同种鱼提供参考,后续研究可以利用该研究方法结合不同鱼类的不同鱼体范围探索适应的感官圈范围.水力学数值模拟计算中,紊流条件越复杂则网格划分越精密,而水流平顺处网格则相对稀疏,在对鱼体周身的水力学环境进行紊流数值模拟时,仅根据水力学条件划分的网格可能不足以满足鱼类捕捉有效水力学信息的要求,需参考鱼类感官范围对水力学网格进行局部加密使得鱼体能够捕获有效的刺激因素完成搜索上溯.此外研究发现,齐口裂腹鱼在上溯运动过程中对流速、感官搜索角度的范围有一定的趋向性,它更偏向于在0.20~0.40m/s的流速区间内以和水平方向呈-49°~49°的扇形感官搜索角度范围进行上溯洄游.
鱼类上溯行为涉及到鱼类的生态行为学与水力学的耦合,是一个比较前沿的学科交叉问题,鱼类在上溯过程中所表现出来的行为也是极其复杂的,对其上溯行为的研究也是非常具有难度的[25],在跟踪单尾鱼样本中首先遇到的困难是问题描述中的组合视角.鱼道的水力信息被认为是一种刺激物,属于欧拉流场,而鱼对这些刺激物的响应属于拉格朗日参照系.运用Eulerian-Lagrangian-Agent方法(ELAM)对模拟鱼类复杂的生态过程具有增值效益[26-27].
生物粒子模型将鱼视作粒子来模拟其上溯行为,其原理为首先对鱼类所在区域的环境流场进行网格划分,其中每个网格节点包含一定的水力学信息,包括上述的流速、流速梯度和湍动能等,然后鱼类感知并评估侧线感知范围内的偏好水力学条件[28],据此调整自身游泳策略进行搜索上溯.模型中鱼类偏好水力学条件可根据AHP层次分析法通过探索鱼类游泳行为对水力学条件的响应关系进行确定[13],此外,利用生物粒子模型预测的鱼类游泳轨迹结果还受数值模拟的网格精度、感官圈搜索范围、水质参数、鱼体理化状态及各种环境因子的影响,各种因子共同影响着鱼类自主游泳行为.如鱼类长时间处于较大紊动和切应力环境下会丧失感官方向,最终导致其游泳能力降低[29];袁佺等[30]指出长薄鳅游泳行为中的回避率随着水体环境总溶解气体过饱和(TDG)的提高而提高;Morrice等[31]得出了河流流量与潮汐对大鳞大马哈鱼幼鱼迁移路径、河口停留时间和生长率具有显著性的影响;LaBone等[32]采用三维耦合水动力水质模型生成的DO场运动算法,提出了基于环境水体溶解氧水平的3种鱼类垂直感观距离.因此,在后续模型构建的过程中,需综合考虑鱼类感官圈内的各种水力学、水质及鱼类理化状态参数的影响以优化生物粒子模型,真实客观模拟鱼类在不同水流环境中的运动行为,为建设高效率过鱼设施及评估过鱼效率提供参考.
4 结论
4.1 采用小于或等于0.30m尺寸的网格解译鱼体行为时,鱼类上溯运动平均游泳速度、转向次数以及转角和转角差均与实际情况无显著性差异.
4.2 鱼类偏向于在0.20~0.40m/s的流速区间内以和水平方向呈-49°~49°的运动搜索范围进行上溯运动.
[1] Silva A T,Lucas M C,Castro-Santos T,et al. The future of fish passage science,engineering,and practice [J]. Fish and Fisheries,2018,19(2):340-362.
[2] 李敏讷,朱海峰,金志军,等.加速流对鲢、鳙幼鱼下行过程中游泳行为的影响 [J]. 水生生物学报,2018,42(3):571-577.
Li M N,Zhu H F,Jin Z J,et al. The effect of accelerating flow on swimming behavior of juvenile aristichthys nobilis and hypophthalmichthys molitrix during downstream migration [J]. Acta Hydrobiologica Sinica,2018,42(3):571-577.
[3] 孙 林,易文敏,张兴磊,等.变流场情景下基于鱼类游泳特性的上溯通道分析[J]. 水力发电学报,2021,40(10):1-9.
Sun L,Yi W M,Zhang X L,et al. Analysis of upstream migrating channels based on characteristics of fish swimming in variable flow field [J]. Journal of Hydroelectric Engineering,2021,40(10):1-9.
[4] 陈凯麒,常仲农,曹晓红,等.我国鱼道的建设现状与展望[J]. 水利学报,2012,43(2):182-188.
Chen K Q,Chang Z N,Cao X H,et al. Recent progress in studies of overland flow resistance [J]. Journal of Hydraulic Engineering,2012,43(2):182-188.
[5] 林俊强,彭期冬,黄真理.河流鱼类鱼卵运动的水力学研究进展[J]. 水利学报,2015,46(7):869-876.
Lin J Q. Peng Q Y,Huang Z L. Review on hydraulics research of fish eggs’ movement in rivers [J]. Journal of Hydraulic Engineering,2015,46(7):869-876.
[6] Zielinski D P,Voller V R,Sorensen P W. A physiologically inspired agent-based approach to model upstream passage of invasive fish at a lock-and-dam [J]. Ecological Modelling,2018,382:18-32.
[7] Montgomery J,Coombs S,Halstead M. Biology of the mechanosensory lateral line in fishes [J]. Reviews in Fish Biology and Fisheries,1995,5(4):399-416.
[8] Goodwin R A,Nestler J M,Anderson J J,et al. Forecasting 3-D fish movement behavior using a Eulerian–Lagrangian–agent method (ELAM) [J]. Ecological Modelling,2006,192(1/2):197-223.
[9] Coombs S. Signal detection theory,lateral-line excitation patterns and prey capture behaviour of mottled sculpin [J]. Animal Behaviour,1999,58(2):421-430.
[10] Gao Z,Andersson H I,Dai H,et al. A new Eulerian–Lagrangian agent method to model fish paths in a vertical slot fishway [J]. Ecological Engineering,2016,88:217-225.
[11] Tan J,Tao L,Gao Z,et al. Modeling fish movement trajectories in relation to hydraulic response relationships in an experimental fishway [J]. Water,2018,10(11):1511.
[12] Zielinski D P,Voller V R,Sorensen P W. A physiologically inspired agent-based approach to model upstream passage of invasive fish at a lock-and-dam [J]. Ecological Modelling,2018,382:18-32.
[13] Zhu L,Li J,Deng Y,et al. Based on a biological particle model to predict the trace behavior of fish [J]. Water Supply,2021,21(8):4044- 4057.
[14] 郭静瑜,廖伯文,周其航,等.泥沙浓度对齐口裂腹鱼生存的影响试验研究[J]. 泥沙研究,2020,45(4):7-12.
Guo J Y,Liao B W,Zhou Q H,et al. Experimental study on influence of sediment concentration to survival of schizothorax prenanti [J]. Journal of Sediment Research,2020,45(4):7-12.
[15] 刘 军.长江上游特有鱼类受威胁及优先保护顺序的定量分析 [J]. 中国环境科学,2004,24(4):395-399.
Liu J. A quantitative analysis on threat and priority of conservation order of the endemic fishes in upper reaches of the Yangtze River [J]. China Environmental Science,2004,24(4):395-399.
[16] Rodriguez Á,Bermúdez M,Rabuñal J R,et al. Optical fish trajectory measurement in fishways through computer vision and artificial neural networks [J]. Journal of Computing in Civil Engineering,2011,25(4): 291-301.
[17] 田 康,张 尧,李金龙,等.基于OpenFOAM几何流体体积方法的波浪数值模拟[J]. 上海交通大学学报,2021,55(1):1.
Tian K,Zhang Y,Li J L,et al. Numerical waves simulation using geometrical VOF method based on OpenFOAM [J]. Journal of Shanghai Jiaotong University,2021,55(1):1.
[18] 廖琦宇.基于全局轨迹分析的视频分割与合成算法研究[D]. 合肥:中国科学技术大学,2015.
Liao Q Y. Research on video segmentation and merging based on global trajectory analyze [D]. Hefei: University of Science and Technology of China,2015.
[19] Bian L. The representation of the environment in the context of individual-based modeling [J]. Ecological Modelling,2003,159(2/3): 279-296.
[20] Liao L,Chen M,An R,et al. Identifying three-dimensional swimming corridors for fish to match their swimming characteristics under different hydropower plant operations: Optimization of entrance location for fish-passing facilities [J]. Science of the Total Environment,2022,822:153599.
[21] 蒋永强,杨忠勇,石小涛,等.基于多重水力学因子的竖缝式鱼道中鱼类上溯轨迹模拟 [J]. 生态学杂志,2018,37(4):1282-1290.
Jiang Y Q,Yang Z Y,Shi X T,et al. The simulation of fish migratory trajectory in a vertical slot fishway based on multihydraulic indices [J]. Chinese Journal of Ecology,2018,37(4):1282-1290.
[22] Arenas A,Politano M,Weber L,et al. Analysis of movements and behavior of smolts swimming in hydropower reservoirs [J]. Ecological Modelling,2015,312:292-307.
[23] Shi X,Ke S,Tu Z,et al. Swimming capability of target fish from eight hydropower stations in China relative to fishway design [J]. Canadian Journal of Fisheries and Aquatic Sciences,2022,79(1):124-132.
[24] 蔡 露,侯轶群,金 瑶,等.鱼游泳能力对体长的响应及其在鱼道设计中的应用 [J]. 农业工程学报,2021,37(5):209-215.
[25] Steel E A,Guttorp P,Anderson J J,et al. Modeling juvenile salmon migration using a simple Markov chain [J]. Journal of Agricultural,Biological,and Environmental Statistics,2001,6(1):80-88.
[26] Kulić T,Lončar G,Kovačević M,et al. Application of agent-based modelling for selecting configuration of vertical slot fishway [J]. Građevinar,2021,73(3):235-247.
[27] Benson T,De Bie J,Gaskell J,et al. Agent-based modelling of juvenile eel migration via selective tidal stream transport [J]. Ecological Modelling,2021,443:109448.
[28] Kanter M J,Coombs S. Rheotaxis and prey detection in uniform currents by Lake Michigan mottled sculpin (Cottus bairdi) [J]. Journal of Experimental Biology,2003,206(1):59-70.
[29] 齐 亮,杨 宇,王 悦,等.鱼类对水动力环境变化的行为响应特征 [J]. 河海大学学报(自然科学版),2012,40(4):438-445.
Qi L,Yang Y,Wang Y,et al. Fish behavior characteristics in response to change of hydrodynamic environment [J]. Journal of Hohai University,2012,40(4):438-445.
[30] 袁 佺,袁 嫄,王远铭,等.长薄鳅对过饱和总溶解气体的回避特征研究 [J]. 水生态学杂志,2017,38(4):77-81.
Yuan Q,Yuan Y,Wang Y M,et al. Avoidance characteristics of leptobotia elongate to total dissolved gas supersaturation [J]. Journal of Hydroecology,2017,38(4):77-81.
[31] Morrice K J,Baptista A M,Burke B J. Environmental and behavioral controls on juvenile Chinook salmon migration pathways in the Columbia River estuary [J]. Ecological Modelling,2020,427:109003.
[32] LaBone E,Justic D,Rose K,et al. Modeling fish movement in 3-D in the Gulf of Mexico hypoxic zone [J]. Estuaries and Coasts,2019,42(6):1662-1685.
Interpretation of sensory query distance during upstream migration of.
DENG Yue-xi1,TANG Xi-liang2,YAN Zhong-luan2,WANG Xiao-ming2,LU Jing-ying3,AN Rui-dong1,LI Jia1*
(1.State Key Laboratory of Hydraulics and Mountain River Engineering,College of Water Resources and Hydropower,Sichuan University,Chengdu 610065,China;2.China Three Gorges Construction Engineering Corporation,Chengdu 610041,China;3.China Three Gorges Corporation,Wuhan 430010,China).,2022,42(8):3882~3889
In this paper,an endemic fish located in Southwest China,(32.32±2.73)cm,was selected as the research object to test the volitional swimming behavior during the upstream migration in an open-channel flume with alternative flow regimes,and to interpret the extent to which swimming speed,turning times,angle and angle difference under different mesh sizes can match with the real swimming trajectory. The results showed that when the mesh size was less than or equal to 0.30m,there was no significant difference between the average swimming speed,the number of turns,the angle and the difference of turns with real swimming data. In addition,fish are more inclined to swim in the velocity ranged from 0.20 to 0.40m/s with the search angle ranged from -49º to 49º in horizontal direction.
mesh size;eco-hydraulics;fish behavior;sensory query distance
X171
A
1000-6923(2022)08-3882-08
2022-01-10
国家自然科学基金资助项目(52122904)
* 责任作者,教授,lijia@scu.edu.cn
邓月曦(1998-),女,四川绵阳人,四川大学硕士研究生,主要从事环境与生态水力学研究.
猜你喜欢
初中生学习指导·提升版(2022年4期)2022-05-11
中学生数理化·八年级物理人教版(2022年4期)2022-04-26
中学生数理化·七年级数学人教版(2021年9期)2021-11-20
大众科学(2020年7期)2020-10-26
儿童时代·幸福宝宝(2020年9期)2020-09-08
模具制造(2019年10期)2020-01-06
小天使·六年级语数英综合(2018年1期)2018-10-08
中国建筑金属结构(2018年4期)2018-05-23
探索科学(2017年5期)2017-06-21
