
野芋提取物通过SFTA1P对肝癌细胞生物学行为的影响*
2022-08-19 03:51陈晓青袁发浒邱文洪
华中科技大学学报(医学版) 2022年4期
陈晓青, 袁发浒 , 黄 丹, 邱文洪
江汉大学医学院 1医学实验中心 2基础医学部,武汉 430056
肝癌是一种严重危害人类健康的恶性肿瘤。目前传统中药由于药性温和,不良反应较少,还可提高机体的免疫力,成为治疗肿瘤的重要方法之一[1]。研究报道地榆乙酸乙酯提取物可抑制肝癌HepG2细胞增殖,促进细胞凋亡[2]。南蛇藤提取物可抑制肝癌细胞的增殖与侵袭转移[3]。研究报道野芋提取物具有抗肿瘤作用[4],但其是否具有抗肝癌的作用及其机制尚不清楚。研究报道显示,lncRNA与肿瘤的进展相关,表面活性剂有关的1假基因(surfactant associated 1 pseudogene,SFTA1P)是一种lncRNA,在肺癌中异常表达[5]。SFTA1P在胃癌患者组织中低表达,与肿瘤大小、淋巴结转移和患者的预后密切相关;SFTA1P过表达可抑制胃癌细胞的增殖、迁移和侵袭[6]。但SFTA1P对肝癌细胞生物行为的影响尚未可知,本实验旨在研究野芋提取物对肝癌细胞生物行为的影响及其机制。
1 材料与方法
1.1 实验材料
肝癌细胞系HCC9204购自上海宾穗生物科技有限公司;野芋药材购自宝芝林大药房;胎牛血清、RPMI-1640培养液购自美国Gibco公司;细胞计数试剂盒8(CCK-8)购自武汉维克赛思科技有限公司;RIPA蛋白裂解液购自上海羽朵生物科技有限公司;Annexin Ⅴ-FITC/PI试剂盒、双荧光素酶报告基因检测试剂盒购自北京Solarbio公司;Trizol试剂、荧光定量试剂盒购自北京百奥莱博科技有限公司;Transwell小室购于美国BD公司。
1.2 野芋提取物的制备
取适量野芋干燥药材,粉碎后用体积浓度为80%的乙醇浸泡(乙醇与野芋用量比为2 mL∶1 g),在37℃条件下提取2 h,提取3次,减压浓缩去除提取液中的乙醇,得到浸膏,将浸膏用乙酸乙酯萃取,乙酸乙酯与浸膏的重量比为1 mL∶1 g;萃取3次,将萃取液减压浓缩后去除乙酸乙酯,得到萃取物即为野芋提取物,将其稀释至所需浓度备用。
1.3 细胞处理与分组
HCC9204细胞用RPMI-1640培养液常规培养,用浓度分别为5、10、15、20 mg/L的野芋提取物处理HCC9204细胞,作为野芋提取物低剂量、中剂量、中高剂量、高剂量组;正常培养的肝癌HCC9204细胞作为对照组。将pcDNA3.1、pcDNA3.1-SFTA1P转染至HCC9204细胞中,记为pcDNA3.1组、pcDNA3.1-SFTA1P组;将si-NC、si-SFTA1P转染至HCC9204细胞后用20 mg/L的野芋提取物处理,记为野芋提取物+si-NC组、野芋提取物+si-SFTA1P组。
1.4 CCK-8检测细胞存活率
取各组细胞,加入CCK-8试剂后孵育2 h,然后用酶标仪检测490 nm处吸光度(A)值。细胞存活率(%)为实验组和对照组A值的比值。
1.5 流式细胞术检测细胞凋亡
取各组细胞,按试剂盒说明加入相应剂量的Annexin Ⅴ-FITC和PI,混匀后避光孵育10 min;上流式细胞仪检测细胞凋亡率。
1.6 克隆形成实验检测细胞增殖
取各组细胞,消化后悬浮细胞,分别按每孔100个细胞接种于6孔板中,大约培养2周,有肉眼可见克隆时停止培养,然后用甲醇固定细胞,再用吉姆萨染色,计数>50个细胞的集落。
1.7 蛋白质印迹(Western blot)法检测Ki-67、Bcl-2、Bax、E-cadherin、N-cadherin蛋白表达
提取细胞总蛋白,并进行定量;然后进行SDS-PAGE电泳,再转至PVDF膜上,用5%的牛血清蛋白封闭1 h;加入一抗4℃过夜,加入二抗室温培养1 h,再用TBST洗涤3次,加入化学发光试剂显影,成像后用Image J分析蛋白条带的灰度水平,以GAPDH为内参计算蛋白表达水平。
1.8 Transwell检测细胞迁移
将200 μL细胞悬液加入Transwell小室的上室,培养24 h后先用多聚甲醛固定,再用结晶紫染色,然后在显微镜下观察计数。
1.9 实时荧光定量PCR(qRT-PCR)检测SFTA1P和miR-665的表达水平
提取总RNA,合成cDNA后按说明书操作进行PCR检测,最终相对表达量用2-ΔΔCt法计算。以GAPDH和U6为内参,SFTA1P上游引物序列:5′-CAGCATTCCAGGTGGGCTTT-3′,下游引物序列:5′-CCTTGTTTGGCTTACTCGTGC-3′;GAPDH上游引物序列:5′-GGGAGGCCAAAAGGGTCAT-3′,下游引物序列:5′-GAGTCCTTCCAC-GATACCAA-3′;miR-665上游引物序列:5′-ACCAGGAGGCTGAGG-3′,下游引物序列:5′-GAGCAGGCTGGAGAA-3′;U6上游引物序列:5′-CTCGCTTCGGCAGCACA-3′,下游引物序列:5′-GTGCAGGGTCCGAGGT-3′。
1.10 双荧光素酶报告实验检测SFTA1P对miR-665的靶向调控
构建含miR-665结合位点的SFTA1P的荧光素酶表达载体WT-SFTA1P和MUT-SFTA1P,将其分别与miR-NC和miR-665共转染至HCC9204细胞中。按照试剂盒说明书检测荧光素酶活性。
1.11 统计学方法

2 结果
2.1 野芋提取物对HCC9204增殖、凋亡、迁移的影响
与对照组比较,野芋提取物各剂量组HCC9204细胞的存活率显著降低,凋亡率显著升高,克隆形成数和迁移细胞数显著减少,Ki-67、N-cadherin、Bcl-2蛋白表达水平显著降低,Bax、E-cadherin蛋白表达水平显著升高(均P<0.05)(图1、表1)。
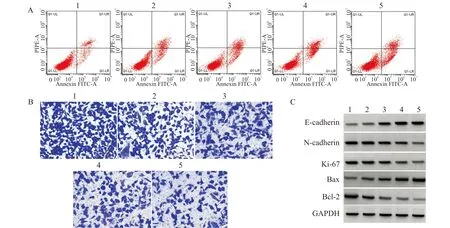
A:野芋提取物对HCC9204凋亡的影响;B:野芋提取物对HCC9204迁移的影响(结晶紫染色,×200);C:野芋提取物对HCC9204中相关蛋白表达的影响;1:对照组;2:野芋提取物低剂量组;3:野芋提取物中剂量组;4:野芋提取物中高剂量组;5:野芋提取物高剂量组图1 野芋提取物对HCC9204凋亡、迁移及相关蛋白表达的影响Fig.1 The effect of Colocasia Antiquorum extract on apoptosis and migration of HCC9204 and expression of related proteins

表1 野芋提取物对HCC9204增殖、凋亡、迁移的影响Table 1 Effect of Colocasia Antiquorum extract on proliferation and apoptosis of
2.2 野芋提取物对HCC9204中SFTA1P表达的影响
与对照组比较,野芋提取物各剂量组SFTA1P表达水平显著升高(均P<0.05)(图2)。

与对照组比较,*P<0.05图2 野芋提取物对HCC9204中SFTA1P表达的影响Fig.2 Effect of Colocasia Antiquorum extract on expression of SFTA1P in HCC9204
2.3 SFTA1P对HCC9204增殖、凋亡及迁移的影响
与pcDNA3.1组相比,pcDNA3.1-SFTA1P组SFTA1P表达水平显著升高,细胞存活率显著降低,凋亡率显著升高,克隆形成数和迁移细胞数显著减少,Ki67、N-cadherin、Bcl-2蛋白表达水平显著降低,Bax、E-cadherin蛋白表达水平显著升高(均P<0.05)(图3、表2)。

A:SFTA1P对HCC9204凋亡的影响;B:SFTA1P对HCC9204迁移的影响(结晶紫染色,×200);C:SFTA1P对HCC9204中相关蛋白表达的影响;1:pcDNA3.1组;2:pcDNA3.1-SFTA1P组图3 SFTA1P对HCC9204凋亡、迁移及相关蛋白表达的影响Fig.3 The effect of SFTA1P on apoptosis and migration of HCC9204 and expression of related proteins
表2 SFTA1P对HCC9204增殖、凋亡及迁移的影响Table 2 Effects of SFTA1P on the proliferation and apoptosis of
2.4 SFTA1P靶向miR-665
Starbase预测显示SFTA1P与miR-665存在结合位点(图4)。与miR-NC组相比,miR-665组中WT-SFTA1P荧光素酶活性显著降低[(1.00±0.07)vs.(0.18±0.02),P<0.05];而MUT-SFTA1P荧光素酶活性无显著差异[(0.99±0.07)vs.(1.02±0.07),P>0.05]。与pcDNA3.1组相比,pcDNA3.1-SFTA1P组中miR-665表达水平显著降低[(0.98±0.05)vs.(0.28±0.03),P<0.05]。

图4 SFTA1P靶向调控miR-665Fig.4 Targeted regulation of miR-665 by SFTA1P
2.5 抑制SFTA1P对野芋提取物处理的HCC9204增殖、凋亡、迁移的影响
与野芋提取物+si-NC组相比,野芋提取物+si-SFTA1P组SFTA1P表达水平显著降低,存活率显著升高,凋亡率显著降低,克隆形成数和迁移细胞数显著增加,Ki67、N-cadherin、Bcl-2蛋白表达水平显著升高,Bax、E-cadherin蛋白表达水平显著降低(P<0.05)(图5、表3)。

A:抑制SFTA1P对野芋提取物处理的HCC9204凋亡的影响;B:抑制SFTA1P对野芋提取物处理的HCC9204迁移的影响(结晶紫染色,×200);C:抑制SFTA1P对野芋提取物处理的HCC9204相关蛋白表达的影响;1:野芋提取物+si-NC组;2:野芋提取物+si-SFTA1P组图5 抑制SFTA1P对野芋提取物处理的HCC9204凋亡、迁移及相关蛋白表达的影响Fig.5 The effect of inhibition of SFTA1P on the apoptosis,migration and related protein expression of HCC9204 treated with Colocasia Antiquorum extract
表3 抑制SFTA1P对野芋提取物处理的HCC9204增殖、凋亡及迁移的影响Table 3 The effect of inhibition of SFTA1P on the proliferation,apoptosis and migration of HCC9204 treated with Colocasia Antiquorum
3 讨论
中医药在肿瘤预防和治疗过程中具有明显的效果,因为毒副作用较小,对防治肝癌复发、转移及延长生存期有一定的帮助,对肝癌治疗有重要价值[7-8]。研究报道,预知子种子提取物能抑制HepG2肝癌细胞的增殖及迁移[9]。半枝莲提取物可抑制肝癌HepG2细胞迁移及侵袭[10]。此外,已有研究证实野芋提取物具有抗肿瘤活性[4]。本实验为研究野芋提取物对肝癌细胞的影响,用不同浓度的野芋提取物处理肝癌细胞HCC9204,结果显示,细胞存活率显著降低,细胞凋亡率显著升高,克隆形成数、迁移细胞数显著减少,Ki-67、N-cadherin、Bcl-2蛋白表达水平显著降低,Bax、E-cadherin蛋白表达水平显著升高,呈剂量依赖性。说明野芋提取物可抑制肝癌细胞增殖、迁移,并促进肝癌细胞凋亡。
研究表明,lncRNA参与多种肿瘤的发生、发展及预后,也与肝癌密切相关[11]。研究报道SFTA1P与肺鳞状细胞癌预后相关[12];SFTA1P在肺鳞状细胞癌中低表达,上调其表达可促进肺鳞状细胞癌细胞凋亡,并增强对顺铂的敏感性[13]。SFTA1P在肺腺癌表达下调,上调其表达可抑制肺腺癌细胞迁移和侵袭[14]。经TCGA数据库分析,SFTA1P在肝细胞癌组织中呈现高表达[15]。因此,我们猜测SFTA1P异常表达与肝癌发展进程有关。我们的实验结果与上述研究一致,在肝癌细胞中,过表达SFTA1P导致细胞存活率显著降低,细胞凋亡率显著升高,克隆形成数、迁移细胞数显著减少,Ki-67、N-cadherin、Bcl-2蛋白表达水平显著降低,Bax、E-cadherin蛋白表达水平显著升高。表明过表达SFTA1P可抑制肝癌HCC9204细胞增殖、迁移,促进细胞凋亡。研究发现中医药提取物可通过调控lncRNA介导的通路而发挥抑癌作用。如槐耳提取物可通过调控H-19/miR-675-5p通路从而减缓乳腺癌进展[16];蒲公英根提取物可抑制胃癌细胞增殖和迁移,其机制可能与靶向lncRNA CCAT1有关[17]。因此,我们深入探究野芋提取物是否通过介导SFTA1P表达从而影响肝癌进展。进一步实验结果显示,野芋提取物处理的肝癌HCC9204细胞中SFTA1P表达水平显著升高,而抑制SFTA1P表达可逆转野芋提取物对HCC9204细胞增殖、迁移和凋亡的影响。说明野芋提取物可调控SFTA1P的表达,其可能通过SFTA1P影响肝癌细胞的进展。
多项证据表明,lncRNA可通过靶向miRNA参与肝细胞癌的发生及发展[18]。此外,有研究表明肝癌患者血清中外泌体miR-665水平升高,与肝癌进展相关,miR-665的水平可能有助于肝癌的临床诊断和预后判断[19]。miR-665通过靶向B型酪氨酸磷酸酶受体(tyrosine phosphatase receptor type B,PTPRB)减少Hippo信号传导,从而促进肝癌细胞的迁移、侵袭和增殖[20]。说明miR-665参与肝癌的发生发展过程。且Sun等[21]研究揭示,lncRNA LIMT可通过吸附miR-665从而促进肝癌细胞索拉菲尼耐药性及上皮间质转化进程。本实验结果同样显示,SFTA1P可在肝癌细胞中与miR-665靶向结合,且双荧光素酶报告实验表明SFTA1P可靶向调控miR-665的表达。提示SFTA1P可能通过调控miR-665的表达影响肝癌细胞的进展。
综上所述,野芋提取物可通过调控SFTA1P/ miR-665轴抑制肝癌HCC9204细胞增殖、迁移,促进细胞凋亡。
猜你喜欢
中老年保健(2022年2期)2022-08-24
中老年保健(2022年1期)2022-08-17
保健医苑(2022年5期)2022-06-10
中老年保健(2021年5期)2021-12-02
中老年保健(2021年12期)2021-11-30
天津医科大学学报(2021年4期)2021-08-21
昆明医科大学学报(2021年2期)2021-03-29
广东蚕业(2021年1期)2021-03-18
军民两用技术与产品(2021年10期)2021-03-16
山东医药(2020年9期)2020-05-20
