
Multifunctional, Sustainable, and Biological Non-Ureolytic Self-Healing Systems for Cement-Based Materials
2022-08-17 01:51MohmmFhimizehPooriPskhshLeeSuiMeJoshBnLeeTnSinghRmn
Engineering 2022年6期
Mohmm Fhimizeh, Poori Pskhsh,*, Lee Sui Me, Josh Bn Lee Tn, R.K. Singh Rmn
a School of Engineering, Monash University Malaysia, Bandar Sunway 47500, Malaysia
b School of Science, Monash University Malaysia, Bandar Sunway 47500, Malaysia
c Tropical Medicine and Biology Multidisciplinary Platform, Monash University Malaysia, Bandar Sunway 47500, Malaysia
d Department of Chemical Engineering & Department of Mechanical and Aerospace Engineering, Monash University, Clayton, VIC 3168, Australia
Keywords:Self-healing concrete Mechanical properties Durability Non-ureolytic pathways Cement composite Sustainability
ABSTRACT Microbially induced calcium carbonate (CaCO3) precipitation (MICP) has been investigated as a sustainable alternative to conventional concrete remediation methods for improving the mechanical properties and durability of concrete structures. To date, urea-dependent MICP is the most widely employed MICP pathway in biological self-healing concrete research as its use has resulted in efficient CaCO3 precipitation rates. NH3 is a byproduct of ureolysis, and can be hazardous to cementitious structures and the health of various species. Accordingly, non-ureolytic bacterial concrete self-healing systems have been developed as eco-friendly alternatives to urea-dependent self-healing systems. Non-ureolytic pathways can improve the physical properties of concrete samples and incorporate the use of waste materials;they have the potential to be cost-effective and sustainable. Moreover, they can be applied in terrestrial and marine environments. To date, research on non-ureolytic concrete self-healing systems has been scarce compared to that on ureolytic systems. This article discusses the advances and challenges in nonureolytic bacterial concrete self-healing studies and highlights the directions for future research.
1. Introduction
Rapidly expanding urban areas and increasingly erratic weather patterns due to anthropogenic greenhouse gas emissions demand more resilient and durable structures with minimal environmental footprint. Improving urban infrastructure has been emphasized as a primary challenge to achieve global sustainable development[1].Extensive research into alternative construction materials and strategies has presented a roadmap of sustainable construction practices that will enable the construction industry to adapt to the challenges in the current millennium. The goal is to create green structures with an extended service life and resilience to various stressors, thus dissociating the expanding construction sector from greenhouse gas emissions [2–4]. One of the proposed strategies is to use smart self-healing cement composite materials that are affected by changes in the structure and respond accordingly without human mediation.
Concrete self-healing systems have been proposed to resolve the structural and environmental problems encountered by traditional concrete structures.These problems include ①crack formation on structure surfaces due to the low tensile strength of concrete, freeze-thawing, and shrinkage; ②ingress of water and corrosive substances through the cracks and pores on the structure’s surface and dispersion through the interconnected voids of the cement matrix;③initiation of extreme structural degradation from an early stage;④inadequate capacity of concrete for autogenous self-healing with respect to time and repair scale; ⑤limitations of manual repair for visible and accessible surface cracks,including time and cost; and ⑥considerable environmental burden posed by the cement industry [5–9]. Concrete possesses an autogenous self-healing ability that occurs with ①the carbonation of calcium hydroxide (Ca(OH)2) to produce calcium carbonate(CaCO3), which can block small pores and heal small cracks;②hydration and swelling of the cement matrix around voids and cracks; and ③transport of dislodged particles into voids and cracks by water current [10]. However, Ca(OH)2supply is finite,and the corresponding rate of unadulterated carbonation is slow.Hence, researchers have attempted to improve the self-healing ability of concrete based on a better understanding of the impact of different curing conditions (e.g., curing under wet, wet–dry, or humid conditions) on concrete self-healing [11–13] and by incorporating autonomous self-healing systems containing chemical[14–17], biological [7,18–22], or mineral [23] healing agents. Note that concrete self-healing systems have been designed and tested on various cement-based materials.In this research field,concrete has been used as a general term encompassing different cement composites.
To overcome the limitations of autogenous concrete selfhealing, researchers have developed autonomous concrete selfhealing systems based on mineral[24],chemical[14–17],and biological healing agents [20,21,25]. These agents are delivered through ①direct addition [18,26], ②encapsulation [20,24], and③vascular systems [27–29]; they may also be immobilized on fibers [30,31]. Chemical-based systems may include a single healing agent, including cyanoacrylates [14] and epoxy [32], or they may rely on the reaction of multiple chemicals, such as twocomponent polyurethane foam [33]. Such chemicals may expand upon crack formation to block the entry of corrosive substances through the crack [33,34]. Chemical concrete self-healing systems offer remarkable advantages, such as a long shelf life for systems based on two chemicals and possibly fast reaction time. However,chemical self-healing systems may pose environmental contamination risks, interfere with future concrete recycling, and possibly result in inadequate cement bonding [35].
Over the past two decades, researchers have investigated biotechnological strategies to repair concrete structures [9,36–38]. Such techniques rely on microorganisms that precipitate CaCO3to heal cracks and block pores in concrete structures. They involve the manual introduction of bacterial spores and nutrients to the surface of cracked concrete samples or their incorporation into these samples as a self-healing mechanism.Calcium carbonate is a natural cement-compatible filler present in concrete that poses no adverse environmental impacts. When concrete cracks, water and atmospheric gases seep into the structure and activate dormant biological healing agents, as illustrated in Fig. 1 [26]. The healing agents fill these cracks by microbially induced CaCO3precipitation(MICP).Most bacteria precipitate CaCO3when calcium is present in the surrounding environment. The presumption is that bacteria precipitate CaCO3to strengthen their extracellular matrix and biofilms to better resist environmental stressors, such as high salinity and mechanical force,or simply as a means to store ions or adjust their concentrations [39]. Note that MICP can be achieved through various processes and under different environmental conditions. Bacteria suited for incorporation in concrete self-healing systems must be ①tolerant of the alkaline concrete environment(pH 9–13),②spore-forming,and ③capable of precipitating CaCO3in the cement environment[26,36,40].Spore formation is essential because vegetative bacterial cells cannot tolerate the concrete mixing process and the highly alkaline cement environment.
Unlike bacteria, spores can withstand harsh conditions while remaining dormant until favorable conditions (O2, water, and nutrient availability) are achieved. This feature can keep spores from germinating until structural faults in concrete structures allow the penetration of water and oxygen required for spore germination. The bacteria that satisfy the above criteria are categorized based on their utilization of predominant MICP pathways.
Biological concrete self-healing systems can be broadly categorized into two groups based on their underlying MICP pathway.Ureolytic systems use urea and a microbial precursor for CaCO3precipitation(Eqs.(1)and(2)),whereas non-ureolytic systems rely on various nutrients and precursors. Thus far, ureolytic systems have received more research attention than non-ureolytic designs,mainly because ureolytic bacteria have been found to induce rapid CaCO3precipitation under specific conditions[35,41–43].Ureolytic bacteria and urease enzymes are well-characterized, and urea hydrolysis is identified to be an efficient MICP pathway[44].However, ureolytic bacteria produce NH4+as a by-product of MICP(1 mol NH4+per mole of CaCO3) [36]. The pH increase caused by NH4+increases the CaCO3precipitation rate in ureolytic selfhealing concrete.However,if NH4+is in contact with nitrifying bacteria or conditions allow for its volatilization as NH3(g) (Eq. (3)),the dissolution of CaCO3may occur following a drop in pH, which can damage concrete structures [45]. Moreover, NH4+is hazardous to the health of various organisms, including humans [46]. The adverse environmental impact of ureolytic concrete self-healing systems can limit their application,hindering their commercialization. Although most MICP studies are based on ureolytic bacteria,these organisms require specific conditions to survive and adequately function, limiting their use in specific environments [41].Accordingly, non-ureolytic bacteria that do not require urea have been investigated and incorporated into concrete self-healing systems.
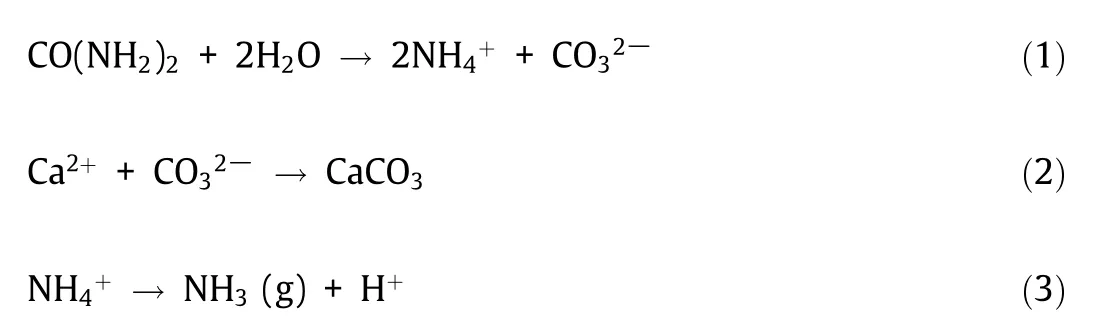
This article reviews the current non-ureolytic systems and their components,research findings,and advances in non-ureolytic biological self-healing concrete development. The remaining challenges that hinder the applicability of these systems and research gaps and unanswered questions are also discussed, and future research directions are highlighted.
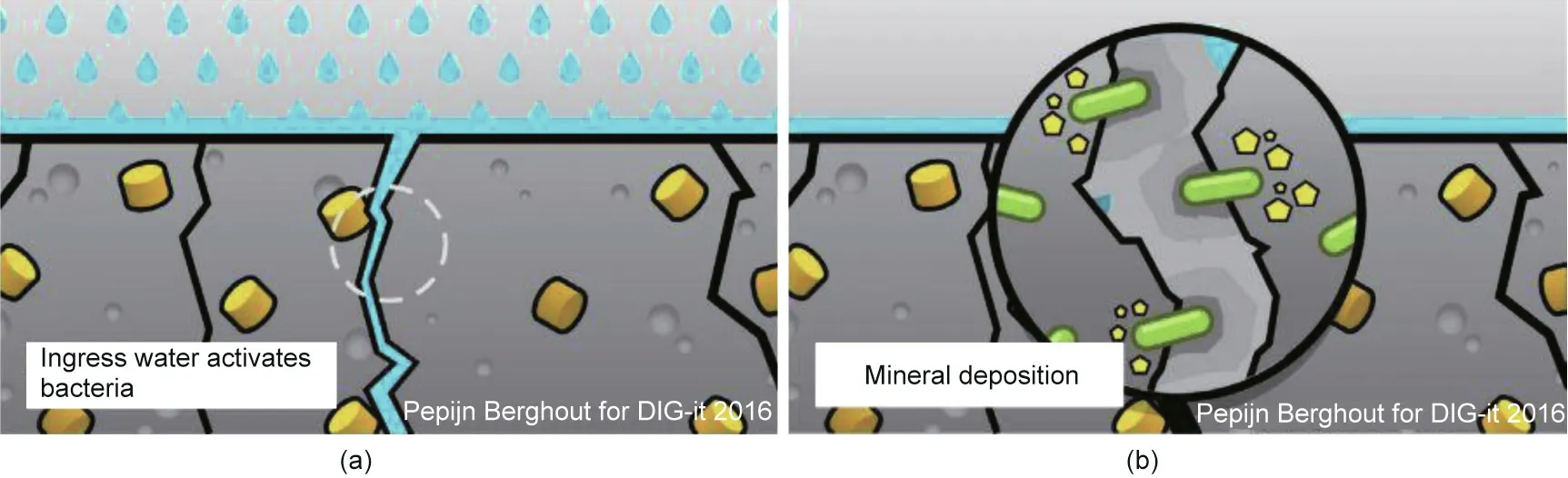
Fig.1. (a)Water flow through concrete cracks revives dormant bacteria,leading to(b)CaCO3 precipitation and crack sealing.Reproduced from Ref.[26] with permission of Elsevier Ltd., © 2017.
2. Non-ureolytic MICP pathways
To date,a wide range of non-ureolytic MICP pathways has been reported. These autotrophic pathways include ① nonmethylotrophic methanogenesis [47], ②oxygenic and anoxygenic photosynthesis [48], ③and heterotrophic pathways, such as conversion of organic salts [36,49], methane oxidation [50], sulfate reduction [51], and denitrification [52]. Generally, non-ureolytic MICP is influenced by ①Ca2+concentration,②dissolved inorganic carbon concentration, ③pH, and ④nucleation site abundance[43,53,54].These parameters,alongside bacteria population levels,bacterial strain, and environmental conditions, which alter bacterial activity (e.g., temperature), are primarily associated with the scale of CaCO3precipitation.The MICP pathways and their applicability to concrete self-healing have been thoroughly reviewed[55],and factors affecting MICP have also been previously examined[22,54]. Among the non-ureolytic MICP pathways described to date, only organic salt oxidation, denitrification, and photosynthesis-driven MICP are compatible with concrete selfhealing. Different problems have hindered the research on other pathways. For example, ammonification-dependent MICP results in significant NH3production; methane-reduction-dependent MICP releases hydrogen sulfide; and the action of sulfatereducing bacteria frequently results in metal corrosion, posing a serious threat to buildings [56–58]. Accordingly, researchers have not investigated pathways for concrete self-healing resulting in unwanted by-products. Newly identified non-ureolytic and alkaliphilic MICP-capable bacterial species that can be used for biological non-ureolytic bacterial concrete self-healing research can expand the described MICP pathways [59].
2.1. Oxidation of organic salts
Certain aerobic (AE) bacteria, which rely on the metabolism of organic compounds (e.g., lactate) for energy production, precipitate CaCO3as a by-product of their metabolic functions. Organic calcium salts, such as calcium lactate (Ca–L) and calcium acetate(Ca–A), provide Ca2+ions necessary for MICP, whereas bacteria can metabolize organic anions for energy production. Certain AE heterotrophic bacteriahave been investigated as biological healing agents for concrete.
Bacterial species such as Bacillus pseudofirmus (B. pseudofirmus)[20,36,40,49,60], Bacillus subtilis (B. subtilis) [61,62], Bacillus cohnii(B. cohnii) [40,63–65], Bacillus alkalinitrilicus (B. alkalinitrilicus)[66], Bacillus thuringiensis (B. thuringiensis) [67], and Bacillus halodurans (B. halodurans) [40] are resistant to alkaline cement environment. When applied to concrete, these bacteria successfully precipitate CaCO3, achieving various distinctive features in concrete self-healing.In vitro and in situ studies have reported efficient CaCO3precipitation by alkaliphiles such as B. pseudofirmus from Ca–L [36,49]. In concrete remediation research, MICP based on the oxidation of organic salts has been used as a surface treatment or self-healing agent. From 1 mol of Ca–L, bacteria can produce 1 mol of CaCO3and 5 mol of CO2(Eq. (4)) [68]. The resulting CO2can react with the abundant Ca(OH)2in the concrete matrix to yield CaCO3(Eq.(5)).One mole of Ca–L can result in the precipitation of 6 mol of CaCO3in the presence of non-limiting Ca2+ions(Eqs. (4) and (5)). The water released during this process contributes to cement hydration and bacterial activity. This pathway has been regarded as a sustainable alternative to ureolysis; it is free of hazardous by-products and can potentially utilize industrial or fermented waste as an organic substrate [69,70].
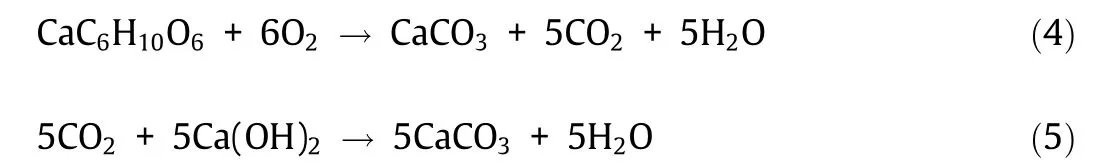
2.2. Denitrification
Unlike ureolysis and oxidation of organic salts,denitrification is independent of atmospheric O2because denitrifying bacteria utilize NO3-as the final electron acceptor for the oxidation of organic substrates to produce CO2, subsequently hydrated to HCO3-(Eqs.(6) and (7)) [71]. Hence, denitrifying bacteria can function in oxygen-deprived environments and precipitate CaCO3under alkaline conditions when Ca2+is available.The denitrification pathway involves four enzymes and intermediates,such as N2O,a substance with potential health hazards to vertebrate health that can act as a steel corrosion inhibitor [72]. Under optimal conditions, N2O does not accumulate in the system;however,the alkaline concrete environment may lead to N2O accumulation [73]. Denitrificationdependent MICP has been investigated for concrete repair and is recommended for low-oxygen environments, such as waterlogged soils, as it is the only anaerobic (AN) non-ureolytic MICP pathway suitable for concrete self-healing [74]. Calcium nitrate(Ca–N), a source of both calcium and nitrate, is often used in denitrification-dependent concrete self-healing systems [25,74].Pseudomonas aeruginosa(P.aeruginosa)and Diaphorobacter nitroreducens(D.nitroreducens)have shown resilience under dehydrated,nutrient-starved, and highly alkaline conditions;accordingly, they have been proposed and investigated for concrete remediation[75].
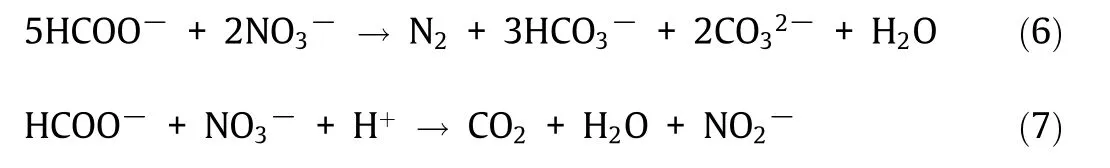
2.3. Photosynthesis
Bacteria can facilitate MICP through CO2uptake via autotrophic and heterotrophic pathways. The precipitation of CaCO3by cyanobacteria depends on the alkaline extracellular media resulting from the CO2uptake during photosynthesis[76].During photosynthesis, CO2is intracellularly concentrated through a biochemical CO2-sequestering mechanism, creating an alkaline environment around the bacterial cell.In the presence of calcium,CaCO3can then be precipitated outside the cell(Eqs.(8)and(9)).The cyanobacteria capacity to capture CO2as organic assimilates and CaCO3precipitates has rendered these bacteria as attractive potential agents for carbon capture and storage (CCS). By abating atmospheric CO2in heterotrophic bacteria,MICP occurrence has been attributed to the action of the carbonic anhydrase enzyme, which facilitates the hydration of CO2[77]. Cyanobacteria Synechococcus PCC8806 has shown tolerance to the concrete environment and can precipitate CaCO3under the environmental conditions of concrete[48].
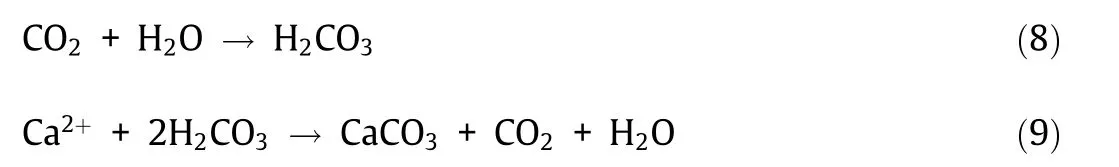
3. Non-ureolytic biological concrete self-healing systems
Non-ureolytic biological concrete self-healing systems have been less investigated than their urea-dependent counterparts.However, they are among the first biological concrete selfhealing systems tested and have been successfully applied in this new scientific field. They offer novel venues for scientific progress and engineering applications. The foundation of their progress could be summarized as follows.
∙Non-ureolytic bacteria can precipitate CaCO3through various metabolic pathways under a wide range of environmental conditions.This property enables the development of versatile healing systems that can rely on different substrates,function under different environmental conditions, and serve various purposes,such as simultaneous concrete self-healing and CCS.
∙Non-ureolytic bacterial concrete self-healing systems can contain more than one bacterial species in bacterial consortia (a mixture of different bacterial species). Although the same is true regarding ureolytic bacteria, non-ureolytic consortia can precipitate CaCO3through different non-ureolytic MICP pathways, exhibiting versatile and flexible self-healing performance. For example, a non-ureolytic consortium consists of organisms that can perform MICP under AE and AN conditions,satisfying the self-healing demands on the concrete surface and oxygen-depleted cracks deep inside a structure.
∙Unlike ureolytic bacteria, non-ureolytic MICP is free from harmful and unwanted by-products and does not require urea in self-healing systems. Although the quantity of NH3/NH4+leaching out of concrete structures containing ureolytic bacteria can seem minute,it may pose a severe threat in the case of large-scale or widespread applications.Concrete is considered the most abundant human-made material. Hence, the largescale application of novel concrete technologies, such as self-healing concrete, in the future is inevitable. At this point,the presence of NH3/NH4+, which is associated with ureolytic systems, can become a problem. In many ecosystems, NH3/NH4+is a natural component and is in equilibrium with other chemical and biological components.Alterations to their concentration may prove disastrous for the natural environment because reactive nitrogen species, such as NH3/NH4+, can lead to eutrophication and loss of biodiversity in forests[78],lakes,and streams [79] as well as deplete the stratospheric ozone[80]; overall, they contribute to anthropogenic global warming. Moreover, NH3/NH4+aggravates numerous health problems, such as cancer as well as respiratory and heart diseases [81].
∙Non-ureolytic biological concrete self-healing systems present an opportunity for sustainable production of self-healing systems that can totally rely on recycled or environmentally friendly components. Through this strategy, biological selfhealing systems can facilitate the introduction of wastederived materials into the market. Non-ureolytic systems can be devised locally based on the available resources in a particular region.
The research attention that has been devoted to non-ureolytic bacterial concrete self-healing is driven by the interest in identifying environmentally friendly alternatives to ureolytic MICP. These two biological approaches employ different bacterial strains, and non-ureolytic systems do not include urea. However, the rest of the components, such as nitrogen and calcium sources, carrier materials,and self-healing evaluation methods,are generally similar. The non-ureolytic systems reported to date are subsequently described and discussed.
3.1. Substrates for biological self-healing concrete
3.1.1. Supporting bacterial growth, activity, and CaCO3precipitation Healthy growth of the bacterial population must be supported before self-healing can commence. Fundamental elements, such as C, H, O, and N, are generally supplied to bio-concrete systems by adding complex media, such as yeast extract. This extract includes the water-soluble remnants of the autolytic digestion of yeast cells mainly composed of singular amino acids or shortchain peptides, nucleic acids, carbohydrates, sugars, vitamins,and salts. Yeast extract is a well-known nitrogen source, rich in B vitamins,and a verified inexpensive and safe microbiological medium that can support the growth of various microbes.Other microbiological medium ingredients, such as nutrient broth, meat extract(e.g.,beef extract),and peptone,can also be used to support bacterial activity in bio-concrete systems. Similar to yeast extract,the foregoing ingredients supply N in amino acids and peptides and contain carbohydrates,vitamins,and salts.The exact composition depends on the manufacturer and substrate source.The presence of Ca is essential for MICP because Ca concentration is a major factor that dictates the MICP rate. The abundance of Ca(OH)2in structures with fresh concrete indicates more bioavailable Ca for biomineralization. Therefore, the presence of a Ca source in bioconcrete systems is not required for early self-healing. As Ca(OH)2is extremely carbonated and its supply is finite, adding a Ca source to the bio-concrete system is essential for the longterm self-healing of aged carbonated cement-based structures[82]. Non-ureolytic bacteria can perform MICP using a variety of Ca substrates. Organic calcium salts, such as Ca–L and Ca–A, have been used as calcium and energy sources because bacteria can metabolize organic anions as carbon sources [83].
The impact of different calcium substrates on CaCO3precipitation and bacterial population growth has been documented. Xu et al.[64]compared the in vitro precipitation of CaCO3by B.cohnii(107cells∙mL-1) supplied with 0.1 mol∙L-1Ca–L, Ca–Glut, and calcium chloride as substrates. Both Ca–L and Ca–Glut are organic salts that function as calcium sources,whereas lactate ions provide carbon,and glutamate ions contain carbon and nitrogen.After 28 d of incubation,the authors reported twice as much precipitation in the cultures supplied with Ca–Glut (0.08 mol∙L-1) compared with those supplied with Ca–L; the lowest CaCO3precipitation rates were reported for cultures supplied with calcium chloride. The authors concluded that the choice of calcium source and initial concentration rather than the CaCO3precipitation rate were the two main factors influencing the total quantity of precipitated CaCO3. Cultures supplied with Ca–L contained higher cell concentrations after one week of incubation compared to cultures containing Ca–Glut (more than 90%). However, after 28 d, cultures supplied with Ca–Glut contained approximately 50% more cells than cultures containing Ca–L; these cultures had the lowest cell survival rate among all cultures up to 28 d. However, after 28 d,they sustained the same population levels as the cultures containing Ca–L. It should be noted that the initial calcium concentration(0.05–0.2 mol∙L-1) did not affect the survival results. Bacterial growth was accompanied by media acidification, with pH values dropping from 9.5 to 7.5–8.0 in cultures containing Ca–L and Ca–Glut. Tziviloglou et al. [83] pre-grew three Bacillus isolates in media, with each medium containing 0.1 mol∙L-1of Ca–L, Ca–A,and sodium glutamate. The authors reported that isolates pregrown on Ca–L and Ca–A could develop a preference for substrates in consecutive generations. The results indicate the formation of a metabolic memory,which may result in the upregulation of certain enzymes in cultures exposed to a nutrient source in previous generations. Metabolic memory can form at different levels of the genetic process and is a phenomenon found across all life domains[84].Future research can identify the causal genetic factors for the metabolic preference reported by Tziviloglou et al. [83]. Lors et al.[49]investigated non-ureolytic MICP by B.pseudofirmus using Ca–L(0.3 mol∙L-1) and yeast extract with and without Ca–N(0.2 mol∙L-1). Researchers reported the total lactate consumption in the presence of Ca–N but not without it. A tenfold increase in the population levels of B. pseudofirmus was observed in the presence of Ca–N. However, this did not increase the CaCO3precipitation,which could be explained by the lack of bioavailable Ca2+from Ca–N. Population levels can significantly influence the quantity of precipitated CaCO3[85]; hence, the addition of adequate concentrations of Ca–N for enhanced growth in MICP-based healing systems may be of interest. Moreover, inorganic calcium salts, such as Ca–N, can react with fresh concrete matrix, releasing free Ca2+,contributing to higher CaCO3precipitation and improving selfhealing performance. Superior bacterial CaCO3precipitation with Ca–L compared with using Ca–N has also been reported for ureolytic bacteria[69].Note that at room temperature,the water solubility of Ca–N is considerably higher than that of Ca–L(i.e.,121 and 4.8 g per 100 mL, respectively).
The effect of different organic calcium sources on concrete structures has also been examined;Ca–N and Ca–F are established concrete admixtures that improve the physical properties of concrete and provide anti-freezing protection [74]. Jonkers et al. [36]reported a slight improvement in the compressive strength of concrete samples containing Ca–L. Tziviloglou et al. [83] reported improved compressive strength in mortar samples containing Ca–L (up to 8%) and Ca–A (up to 13.4%) when supplied in 0.56%–2.24% of cement weight after 28 d of curing. Ducasse-Lapeyrusse et al. [86] found that after a month of curing, Ca–L and Ca–Gluc aided in sealing large fresh cracks (> 150 μm) with calcite and ettringite, which were self-healing products derived by increasing calcium and carbonate concentrations, respectively. Both Ca–Gluc and Ca–L had no significant sealing effect on small cracks. Further research into the microstructure of cement and concrete samples supplemented with organic calcium salts can reveal whether the impact of organic calcium salts is due to the increase in Ca2+concentration or whether organic anions perform a role in settling,gel formation, and cement matrix hydration.
3.1.2.Waste as nutrient or calcium source for non-ureolytic biological self-healing concrete
Non-ureolytic MICP is considered an environmentally friendly alternative to ureolytic MICP; hence, substrates necessary for non-ureolytic MICP must also be sourced through sustainable processes to harness a truly sustainable product. Incorporating waste materials has been suggested to lower the production cost of bioconcrete healing agents and increase the product sustainability factor [26,87–91]. For non-ureolytic systems, Ca–L has been a popular calcium substrate because it improves the physical properties of concrete and serves as a calcium and carbon source for non-ureolytic bacteria [36]. Ca–L can be sourced from fermented vegetable waste [70], recycled poly(lactic acid) (PLA) [92], and paper mill waste[93],possibly contributing to the economic feasibility and sustainable nature of non-ureolytic bacterial concrete self-healing agents. Mors and Jonkers [26] tested a non-ureolytic system containing lactic derivatives, such as lactic acid (a calcium source)and non-ureolytic bacterial spores; they reported that significant water tightness was regained by the cracked samples.Lactic acid has been found to improve cement hydration by enhancing the crystalline structure of cement and reacting with Ca(OH)2from fresh cement to produce Ca–L [94]. The use of Ca–N to sustain denitrifying bacteria has improved bacterial survival; however, it has also resulted in lower quantities of CaCO3precipitation than when Ca–L is employed [49]. The combined use of Ca–L and Ca–N also improved the functionality of non-nitrifying bacteria through survival preservation. Experts suggest that calcium and nitrate do not fully dissociate from each other and continue to be unavailable to the bacteria to some degree.The structural integrity of concrete is also improved by Ca–A, a known environmentally friendly de-icing agent sourced from vegetable waste conversion into acetic acid [95]. Jin et al. [95] converted more than 22% of the total organic carbon in vegetable waste into acetic acid and used oyster shells as an eco-friendly calcium source to obtain Ca–A. Another organic calcium salt, Ca–F, previously tested as a nutrient source for non-ureolytic bacterial concrete self-healing systems, can be sourced from chlor-alkali industrial waste [96].Waste incorporation into self-healing systems reduces production costs and keeps hazardous materials from landfills and the environment, further contributing to the sustainability of bio-based concrete self-healing systems. A novel strategy for integrating waste and waste-derived substances into biological self-healing concrete was reported by Vermeer et al. [97]. The technique included refining polyhydroxyalkanoate (PHA)-rich wastewater residue to be used as a bacterial nutrient in a non-ureolytic biological concrete self-healing system.With this approach,the researchers proposed an alternative use for PHAs that lack sufficient quality for conventional bioplastic application. The comparison of selfhealing results of specimens with and without waste-derived PHA as bacterial nutrients for non-ureolytic B.cohnii bacteria indicates that the former has higher crack-healing efficiency and reduction in water absorption.It should be noted that B.cohnii produced extracellular enzymes to metabolize the PHA. This research approach utilizes biological self-healing concrete as a niche field for waste-derived products whose use is not economically viable based on conventional applications of concrete. Moreover, it identifies a pathway in which biological self-healing concrete can contribute to sustainable resource management.
Song et al. [92] recycled PLA waste into Ca–L using a reusable inert ionic catalyst, deriving > 70% Ca–L yield [93]. Lactic acid was produced from the mixture of softwood pre-hydrolysate and paper mill waste, obtaining Ca–L as an unwanted by-product.Therefore, concrete self-healing systems can utilize waste materials from other industries, contributing to economic feasibility and sustainable development. Certain industrial waste materials,such as blast furnace slags and waste glass, have become established choices for cement replacement [98–101]. Research into similar waste materials, including furnace slag from steel, copper,zinc,and iron production,has shown promising results as potential cement replacements as they are rich in cementitious materials such as Ca and Mg oxides, which may be harnessed as biomineralization precursors [89,102,103]. Note that when such oxides are employed as a calcium source for MICP, they can cause undesirable cement expansion [104], which can be prevented by immobilizing the biological healing agents.Calcium oxides can also be derived from recycled eggshells [105] and shells of aquatic biomineralizing animals, such as mollusks and oysters [106,107].
Røyne et al. [108] described the microbial acid digestion of limestone by Bacillus safensis and subsequent re-precipitation by ureolytic bacteria in a process termed ‘‘the BioZEment approach”(Fig. 2). This technique sustainably converts limestone into an inexpensive calcium source for MICP(Stage I).the ureolytic bacteria then increase the slurry’s pH and precipitate calcite crystals(Stage II). The pH increase is mainly due to the presence of NH4+ions resulting from ureolysis. Even in the absence of NH4+, the interaction between slurry and amino acids from the added bacterial nutrients can increase the pH level to 7.The possibility of using this method may depend on which type of non-ureolytic bacteria is used and require a de-acidification step. We further conducted a life cycle assessment and studied the environmental benefits of this technique compared with conventional concrete production by assessing the global warming potential of the process [109].The findings showed that the BioZEment approach has a lower global warming potential than the conventional concrete production by more than 70%; however, it requires more land area and possibly causes the occurrence of eutrophication and NH3admission problems. The cost of using the BioZEment approach was estimated to be more than 10% of conventional concrete production.
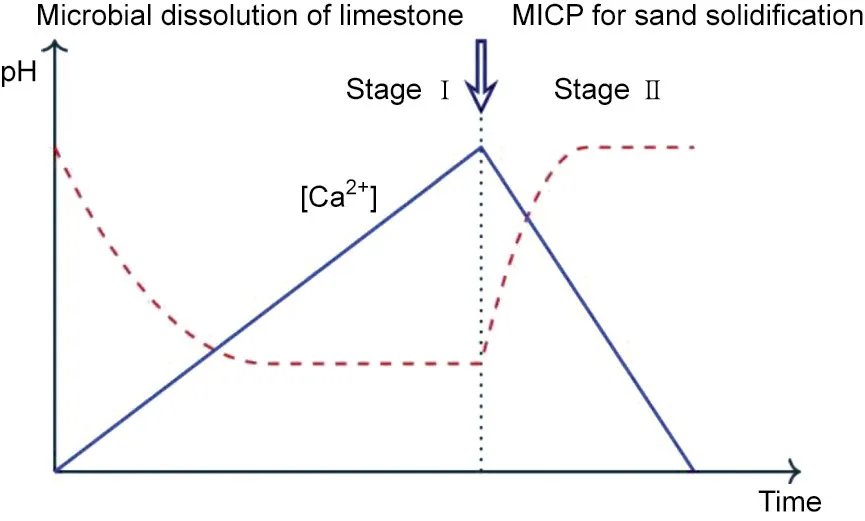
Fig. 2. Schematic of proposed two-stage limestone dissolution–recrystallization process (adapted from Ref. [108] and reproduced based on Creative Commons license).
The investigations described in this section demonstrate the utilization of various waste and by-product materials as bacterial nutrients or calcium sources for MICP of non-ureolytic selfhealing concrete. Moreover, research shows how non-ureolytic biological self-healing concrete can be a point of convergence for the construction industry, wastewater treatment sector, and businesses that produce organic refuse or metal-rich waste.This opportunity provides the researchers of non-ureolytic biological concrete self-healing systems with the prospect of expanding their work to include other industrial waste and by-products. Several practical steps that can benefit future research on alternative nutrient sources for non-ureolytic biological concrete self-healing are available. For instance, establishing a clear understanding of the metabolic processes performed by bacteria can aid researchers in predicting the impact of various factors (e.g., bacterial population,temperature, pH, carriers) on metabolism. Furthermore, this can help researchers in identifying the metabolites or remnants left after the digestion of nutrient sources to ensure that harmful byproducts, which can damage the concrete structure and carrier material, or inhibit bacterial growth, are not producedover time.Another critical step is a life-cycle assessment of the self-healing system that may be similar to the evaluation performed by Mors and Jonkers [26]; this assessment considers the environmental footprint of the alternative nutrient or calcium source. Moreover,it allows for a more meticulous examination of the system’s sustainability and can aid in determining elements that have not been previously considered in a system’s ecological footprint.
3.2. Immobilization and survival assessment of non-ureolytic healing agents
Although the carrier materials used for biological self-healing between ureolytic and non-ureolytic systems do not differ, the methods used to evaluate bacteria survival after immobilization and application vary.Rajczakowska et al.[110]thoroughly reviewed the impact of different carriers used in biological self-healing concrete on the physical properties of cement/concrete samples.In this study, a review of the techniques employed to evaluate bacterial survival after immobilization is presented.Researchers who investigate ureolytic systems frequently use the urease enzyme activity by the total ammonia nitrogen determination method to analyze ureolytic bacteria activity. As this method is not applicable to nonureolytic systems,several other methods have been used to determine the bacterial activity in non-ureolytic systems.
3.2.1. Direct enumeration
Direct enumeration of microbial cells involves the quantification of living microbes by growing the cells on specific growth media.Jonkers et al. [36] directly added 2.4×108spores of B. cohnii per cubic centimeter of cement samples and estimated the spore survival by crushing the samples after 9, 22, 42, and 153 d of curing.They used the most probable number technique to measure the bacterial population and reported that the bacterial activity after 9 d was 1% and then decreased to the minimum test detection limit(<500 cells∙cm-3)after four months.There is a possibility that the high mechanical force used to free the bacterial cells from the cement matrix during this test could have altered the results.
Sharma et al. [40]also directly added spores of B.pseudofirmus,B. cohnii, and B. halodurans into cement paste. Less than 5% of the initial B. pseudofirmus inoculum was recovered after 7 d, which decreased to less than 1%after 28 d of water immersion.The spore survival rate of B.cohnii and B.halodurans also decreased in a similar manner as that of B.pseudofirmus;however,their spore loss was more drastic. Overall, the direct addition of spores to cement is related to low spore survivability, which is possibly due to the highly alkaline cement environment, lack of access to nutrients or oxygen, or susceptibility to shear forces during mixing and settling.
Direct enumeration has also been used to assess the survival of bacteria immobilized in/on a solid carrier. A study incorporated B.pseudofirmus in expanded clay particles(ECPs)by vacuum impregnation (2.2×107colony-forming units per gram (CFU∙g-1) [111].The loaded clay particles were then coated with eight different materials and incorporated into concrete specimens. Viability assessment was conducted by direct enumeration of B. pseudofirmus bacteria from powdered biological self-healing concrete specimens of different ages. With this technique, the authors assessed the protective ability of various coatings and reported superior protection by coatings such as MgO and styrene–acrylate, which block the pores of ECPs. They also reported that the sensitivity of this technique was not optimal but was sufficient to distinguish high and low levels of protection.
3.2.2. NO3-/NO2-consumption
The measurement of the consumption of NO3-/NO2-has been a reliable tool for assessing the activity of bacteria with denitrifying ability. Ers¸an et al. [72] studied the survival of vacuumencapsulated pure cultures of NO3-reducing P. aeruginosa or D.nitroreducens in diatomaceous earth, expanded clay, and granular activated carbon after addition to mortar.They also performed this test for self-protecting NO3-reducing bacteria mixtures termed as the activated compact denitrifying core. Survival was then assessed by measuring the NO3-and NO2-consumptions of bacteria removed from fractured mortars by resuspension and lowfrequency sonication. Among the carrier materials used for pure cultures,expanded clay and activated carbon offered better protection to the bacteria than diatomaceous earth did. The concentrations of NO3-/NO2-were quantified using ion chromatography[72,112].
3.2.3. Measurement of O2consumption for bacterial activity
An established method for deducing bacterial activity is the measurement of O2uptake by bacteria. Researchers have used this method to show bacterial activity in self-healing systems and bioconcrete samples. Wiktor and Jonkers [66] vacuum-impregnated ECPs with Ca–L, yeast extract, and B. alkalinitrilicus. By employing O2consumption measurements, they found that the bacteria remained viable after nine months inside the cement. Sierra-Beltran et al.[63]impregnated lightweight aggregates(LWAs)with Ca–L, yeast extract, and B. cohnii spores and measured the O2consumption to elucidate the bacterial activity in the mortars. The results indicated bacterial activity after three months of casting the samples.Mors and Jonkers[26]evaluated the O2uptake of mortars incorporated with a bacterial lactic acid derivative for a nonureolyticself-healingsystem.BasedontherecordedO2consumption,they verified that the viability of samples with bacteria was superior to that of abiotic control specimens.From the O2consumption measurements,they also determined that the nutrient levels had reached a sufficiently low quantity that limited bacterial activity after a week.
The measurement of O2consumption has also been used to verify the viability of alginate-immobilized bacterial spores[113,114].Palin et al.[113]reported lower dissolved O2concentrations in the 1 mm boundary above the bacterial capsules submerged in artificial seawater at 8 °C compared with those of abiotic control specimens. The dissolved O2levels reached the lowest recorded concentration on the second day of submersion. This technique demonstrated bacterial activity and metabolism under cold marine conditions. However, the O2consumption assessment did not allow the collection of precise evidence for the survivability of the bacterial population and evidence regarding whether the bacterial spores remained inside the capsules or ventured out. Furthermore, the immobilization efficiency of the healing agents by calcium-alginate beads was unclear. The porous hydrogel tends to leach the healing agents during the calcium bath stage of capsule preparation and during the suspension. The described instances of O2measurement demonstrate that this technique can enable researchers to verify the general viability of immobilized healing agents and obtain an estimate of nutrient consumption; however, these measurements do not provide quantitative information regarding the surviving bacterial population. Additionally, no conclusions can be drawn regarding the success of the biological healing agents in the immobilization process (i.e.,the extent to which the agents provided immobilization or whether the vacuum encapsulation impacted the spore survival).The above studies have all provided qualitative proof of spore survival after immobilization, indicating that these survival assessment methods do not provide a precise measure of the bacterial survival or the impact of the immobilization process and the concrete environment on the healing agents.They only indicate that a detectable quantity of the bacteria has remained viable for concrete self-healing. Hence, the impact of immobilization processes on spore survival, the germination of viable spores, and the exact degree of protection(e.g.,carriers)of bacterial spores remain speculative. The qualitative results offer assurance that spores survive the various immobilization processes. However, for the largescale production of biological self-healing agents, a quantitative analysis of the impact of immobilization processes is necessary.
To resolve these problems, we attempted to quantitatively assess the impact of hydrogel immobilization on bacterial spores[20].We immobilized the spores of B.pseudofirmus in calcium alginate capsules using the ionic gelation method, which involved the crosslinking of alginate monomers with a divalent cation (e.g.,Ca2+). Then, an optimized de-crosslinking procedure was implemented to dissolve the alginate capsules, and the spores were quantified using direct enumeration. The results suggested that virtually all of the spores had been successfully encapsulated inside the capsules with no loss of viability or occurrence of leaching during the encapsulation process.Furthermore,the presence of nutrients did not impact the efficiency of the immobilization procedure.The de-crosslinking method could not dissolve the alginate capsules removed from cement, possibly due to residual cement.However, the survival of the bacteria in alginate capsules placed in cement can be assessed if the outermost layer can be removed,that is, if the coated alginate capsule is similar to the chitosancoated alginate capsule described by Gao et al. [60].
The impact of the immobilization procedure on biological selfhealing agents can be better understood by thoroughly analyzing the interaction between agents and protective carriers.This analysis can elucidate whether the carrier requires improvement to better retain or protect the healing agents to ultimately fine-tune the protective carriers of the material support and provide long-term functionality to the healing agents. Assuming that a hypothetical system is incorporated with 108spores∙g-1of a carrier, if 99.99%of the spores do not survive the immobilization or cement casting,106spores∙g-1of the carrier can still provide self-healing results in the form of enhanced mechanical performance, crack healing, and CaCO3precipitation. Moreover, the bacterial samples consume more O2than the control samples. Nevertheless, the efficiency of self-healing is significantly reduced.
Similarly,if a carrier cannot properly keep healing agents,such as yeast extract,shielded from the cement matrix,the self-healing efficiency and functionality can be reduced,and the cement matrix may sustain the deleterious impact posed by yeast extract or similar MICP precursors. Therefore, researchers cannot ignore the efficiency of immobilization techniques and carrier materials in protecting and holding bacterial spores and other healing agents.Note that the nature of certain carrier materials for protecting non-ureolytic self-healing agents may not allow the precise and direct assessment of the survivability and viability of healing agents.Palin et al.[114]used the O2measurement data to estimate a possible amount of CaCO3precipitation; however, they did not report the accuracy of the estimated quantity with respect to the obtained precipitation quantity; nevertheless, this approach can present a strategy for researchers. The correlation of in vitro characterization results(e.g.,the relationship between CaCO3precipitation quantity with estimates of such a performance indicator(e.g.,O2consumption)) can produce a more accurate survivability estimate. Further, the viability of healing agents may even aid in explaining the self-healing behavior of the system after mechanical characterization.
4. Effect of non-ureolytic bacterial concrete self-healing systems on mechanical, physical, and durability properties of concrete
Non-ureolytic biological concrete self-healing systems can protect the biological healing agents and provide the required medium for self-healing action. In contrast, biological healing agents perform the self-healing function following a trigger (e.g., structural fault or crack) that promotes the germination of dormant spores and results in the bioavailability of nutrients and O2. Overall,non-ureolytic biological self-healing systems can reinforce the concrete structure and improve its durability and mechanical and physical properties. Section 3 describes the relationship between the biological and carrier subsystems of non-ureolytic biological concrete self-healing systems and the protective function of the systems. This section describes the self-healing functions (e.g.,the impact of non-ureolytic self-healing systems on the mechanical and physical properties and the durability of concrete specimens) of non-ureolytic biological concrete self-healing systems.
The self-healing ability of non-ureolytic systems in concrete has been studied by comparing the variations in mechanical parameters(e.g.,compressive and flexural strengths)and physical properties(e.g.,changes in porosity and crack dimensions)of cement and concrete specimens.Variations in the type and quantity of bacteria,carrier material,nutrients,calcium additive,specimen composition,and curing conditions do not allow a robust comparison of reported results. Nonetheless, similar measurement techniques have been used to assess the durability as well as the mechanical and physical properties of test specimens before and after self-healing. This commonality allows for a meaningful interpretation of the impact of healing agents on the properties of the specimens in their early stages and the self-healing ability of different systems.
4.1. Mechanical properties
4.1.1. Compressive strength
The compressive strength of concrete has an established correlation with the durability of concrete structures. Inadequate compressibility can result in crack formation due to increased sensitivity to loads and pressure. Such cracks then facilitate the concrete structure’s degradation by increasing the exposure of the cement matrix and reinforcements to corrosive agencies (i.e.,similar to high-porosity concrete structures) [115]. Hence, the compressive strength of experimental concrete samples containing additives is typically evaluated to determine the impact of additives on the structural strength and concrete durability. This evaluation has been widely implemented to assess the impact of self-healing systems on early concrete properties and the degree to which self-healing systems enhance the structural strength of concrete over time.
Mors and Jonkers [26] showed that the direct addition of 6×108B.pseudofirmus spores has a retarding effect on the cement sample’s initial compressive strength. Over 28 d, the authors reported a decrease in the frequency of larger pores (0.8–1 μm in diameter) in cement specimens containing healing agents compared with that of control specimens in which the microcracks(0.01–0.1 μm)remain unblocked.The compressive strength of biotic mortars was initially 60% lower than that of abiotic mortars.After 14 d,their strengths became similar due to bacterial precipitation and remained unchanged up to 56 d.Mondal and Ghosh[62]incorporated 103, 105, and 107cells∙mL-1of B. subtilis spores into concrete.They reported higher compressive strengths for all bacterial samples than those of the control specimens after 3 d, continuing up to 28 d. During this time, the highest gain in compressive strength was observed in samples containing 105cells∙mL-1(a 27%increase compared with those of the control specimens). Lower water absorption rates were also observed, namely, 13%, 23%,and 27%, for the specimens with 103, 105, and 107cells∙mL-1,respectively, compared with those of the corresponding abiotic control samples.
Khaliq and Ehsan [61] incorporated 2.8×108cells∙mL-1of B.subtilis and Ca–L spores either directly or after LWA or graphite nanoparticle (GNP) immobilization and reported improved compressive strengths in bacterial samples compared with those of the control specimens after 3 d and up to 28 d, leading to a maximum compressive strength increase of 12% with LWAimmobilized bacteria and 9.8% with GNP-immobilized bacteria but only a 3% increase with direct bacteria addition. The strength improvement was attributed to the bacterial CaCO3precipitation and improved matrix packing due to aggregate replacement by LWA and GNP, leading to a decrease in the interfacial transition zone (ITZ) formation. Moreover, the interaction between the bacteria and carrier surface may also affect the healing rate, create nucleation sites, and promote biofilm formation that may possibly affect MICP in a manner resembling the impact of soil particles on CaCO3precipitation caused by soil bacteria [116]. In vitro MICP evaluation by B. subtilis using Ca–A revealed that B.subtilis promotes CaCO3precipitation by biofilm formation[117]. Prior research has also noted the importance of biofilm formation for MICP by B. subtilis [118] and B. cohnii [64]. Schwantes-Cezario et al. [117] examined in vitro CaCO3precipitation by B.subtilis using Ca–A (0.25% w/w). They reported that B. subtilis promotes CaCO3precipitation by biofilm formation and associated the rise in pH with biofilm development. Hence, attention to bacteria-carrier interactions can lead to beneficial discoveries,especially when the healing agent is a biofilm-forming bacteria.Such an approach may focus on factors affecting biofilm development, including water availability [119].
The reported improved compressive strength upon the direct addition of healing agents by Khaliq and Ehsan [61] differs from the findings of Jonkers et al. [36]; the latter suggests that the retarding effect of bacterial spores on the compressive strength of concrete may be species-dependent. This suggestion is possible because both studies used Ca–L as the primary nutrient source and comparable spore concentrations. Moreover, the data indicates a slight positive impact of 10% by Ca–L on compressive strength[36]. The comparison between the findings of Khaliq and Ehsan[61] and those of Ramachandran et al. [37] (who directly incorporated 7.2×107cells∙cm-3of ureolytic bacteria Bacillus pasteurii and found no compressive strength improvement) suggests that B.subtilis is a better choice for direct addition in terms of compressive strength. However, the direct addition of healing agents to concrete can be disadvantageous considering the structural integrity of concrete and self-healing efficiency.
Luo and Qian[120]compared the self-healing capacity of a system composed of non-ureolytic bacterial spores, Ca–L, Ca–N, and Ca–F. The authors reported the impact of bacteria/Ca–F (1% w/w)on the compressive strength of cement samples, namely, decreasing strength with increasing bacteria/Ca–F concentration. The effect of varying levels of calcium substrate on compressive strength was also evident from the reported compressive strength results of bacteria/Ca–L.The best performance was observed when 3%w/w of bacteria/Ca–L was added to cement samples,resulting in an initial loss in compressive strength; however, after 10 d, their compressibility surpassed those of the control samples. Bacteria with Ca–N decreased the compressive strength with increasing Ca–N levels. It was reported that the pore size distribution was associated with the calcium substrate used. The control specimen primarily had pores in the size range of 10–100 nm, but the addition of self-healing agents shifted the pore size distribution to the range of 100–1000 nm.The authors suggested this observation as a possible explanation for variations in compressive strength and reported an adverse impact on the cement’s setting time when bacteria/Ca–L was used; in contrast, bacteria/Ca–N and bacteria/Ca–F reduced the cement setting time. This pattern was also reported for the three substrates regarding their effect on the hydration kinetics of cement. These findings improve the understanding of the impact of self-healing systems and their components on cementitious structures and system optimization.
4.1.2. Flexural strength
The bending of concrete structure due to external forces often lead to crack formation because concrete has low flexural strength(usually 10%–20% of its compressive strength). Accordingly, the impact of concrete self-healing systems on flexural strength has been assessed as a self-healing benchmark.
In an earlier study,Xu and Yao[65]assessed the healing ability of 107cells∙mL-1B.cohnii spores and Ca–L or Ca–Glut when used as a surface treatment. They further assessed the healing system’s impact on the flexural strength of the mortar when applied as surface treatment or when incorporated as a self-healing system.They reported no adverse impacts on flexural strength due to individual system components when incorporated into mortars,even noting a slight improvement in the case of Ca–L addition in an earlier study[36]. After 28 d, the flexural strength of surface-treated mortars was higher than that of mortars with healing agents. Among the samples with incorporated healing agents, the flexural strength of those containing Ca–Glut was two times greater than that of the control samples; the samples containing spores and Ca–L did not perform well.The higher flexural strength recovery of samples with Ca–Glut was attributed to the higher CaCO3conversion of Ca–Glut by the bacteria.This resulted in a larger and denser transition zone between the biologically deposited mineral layer and cement matrix(Fig.3[65]),forming a strong bond between the deposition layer and cement matrix. The flexural strength results were confirmed using grid nanoindentation,and the hardness and modulus of the mortar,the outer precipitates,and the transition zone of different specimens were measured. The highest values of modulus and hardness were observed in the transition zone of the sample with Ca–Glut, and the lowest measurements were observed in the transition zone of the Ca–L sample.

Fig. 3. Scanning electron micrographs of transition zone in (a) Ca–L and (b) Ca–Glut specimens. Reproduced from Ref. [65] with permission of Elsevier Ltd., © 2014.
Sierra-Beltran et al.[63]impregnated LWA with Ca–L and yeast extract, and B. cohnii spores were added to a strain-hardening cement-based composite(SHCC).The presence of bacteria resulted in improved flexural strength compared with that of the control specimen. Further, the healing agents only slightly improved the mechanical properties of the SHCC with an insignificant amount of CaCO3precipitation compared to that of the control samples. The inadequate nutrition levels was noted as a possible explanation for the lack of distinguishable CaCO3precipitates in the bacterial samples and suggested further studies. There is a dearth of reported literature on this topic; however, this study can serve as a baseline for evaluating future healing systems for cement variants.
We had previously incorporated calcium alginate-immobilized B. pseudofirmus spores and nutrient broth into cement paste and cement mortar and found initial flexural strength losses of 38.96%and 58.50%for the paste and mortar,respectively[20].This is expected due to the detrimental impact of superabsorbent polymers (SAPs) on the cement matrix due to excessive water uptake,leading to void formation in the matrix [121]. The analysis of artificially cracked samples after 56 d of wet–dry incubation revealed 32.5% and 39.6% regain levels in the flexural strengths of cement paste and mortar samples, respectively. Compared with those of abiotic control samples, bacterial presence improved the regain levels in the flexural strength of cement paste and mortar by 17.1% and 10.3%, respectively. Although the initial loss in strength was extremely high for the 56 d self-healing action to compensate,the results indicated that calcium alginate was a promising carrier capable of supporting the healing agents up to 56 d. The results further showed that the presence of sand in the mortar accelerated the regain in flexural strength,with 69%occurring in the first 28 d of incubation.In contrast,the regain in the flexural strength of the cement paste specimen was mainly observed in the latter 28 d(67%); this can be attributed to sand particles acting as additional nucleation sites of CaCO3precipitation. This highlights the potential of calcium alginate capsules for protecting the healing agents over extended periods. Longer incubation periods are necessary to better understand how such a system can be improved for extended self-healing performance and assessed in future research.
Gao et al. [60] coated calcium alginate capsules containing B.pseudofirmus with chitosan and recorded a 3.78%increase in flexural strength compared with that of the capsule-free control specimen; only 1.2%–1.5% (with capsule/with cement) of the capsules were included, which decreased the expected strength reduction.Furthermore, the reduced swelling due to the pH-responsive nature of chitosan and high drying temperature (65 °C) can decrease the swelling and water uptake of capsules. Although the internal curing potential of alginate capsules is severely diminished, the authors have illustrated how hydrogels can be used as bacterial carriers in concrete without initial loss in strength. Future studies on SAP-encapsulated biological concrete self-healing can investigate the impact of MICP-driven self-healing on the drying shrinkage and autogenous shrinkage of cement-based specimens because recent findings suggest that a possible increase in mass loss and shrinkage may be exacerbated by SAP addition[122,123].
4.2. Physical properties and durability
The impact of non-ureolytic biological self-healing systems on concrete durability has primarily been assessed by investigating the water permeability of structures. Non-ureolytic concrete selfhealing systems perform this function through pore and void blocking as well as crack healing by CaCO3precipitation. Consequently,concrete durability against chemical and physical deterioration is enhanced.
4.2.1. Porosity
Although fluids can enter concrete via several transport mechanisms, the most significant parameter for concrete degradation is the extent to which the cement matrix’s interconnected pore system is further exposed to the environment through microcracks[124]. Therefore, limiting concrete exposure to corrosions by reducing concrete porosity has been a strategy to achieve more durable construction materials [125,126].
Mondal and Ghosh [62] incorporated 103, 105, and 107cells∙mL-1of B. subtilis spores into concrete and reported lower water absorption rates of 13%,23%,and 27%,respectively;they also reported that the water penetration depths compared with those of corresponding abiotic control samples were reduced (Fig. 4).However, as mentioned in Section 4.1.1, the highest compressive strength improvement was observed in specimens with 105cells∙mL-1, suggesting a discrepancy between compressive strength and porosity measurements. The discrepancy was attributed to the higher surface CaCO3precipitation with higher bacterial concentration. This reduced the entry of water and oxygen into the cement matrix from an early stage that could have interfered with the cement hydration and bacterial activity inside the matrix. The difference observed between improvements in porosity and compressive strength due to different bacteria levels can have practical importance for applications in different environments or under different load-induced pressures.
Luo and Qian[120]compared the self-healing capacity of a system composed of non-ureolytic bacterial spores and Ca–L,Ca–N,or Ca–F; they reported that the pore size distribution was associated with the calcium substrate used. The control specimen primarily had pores in the range of 10–100 nm,whereas the addition of selfhealing agents shifted the pore size distribution to larger pores in the range 100–1000 nm.
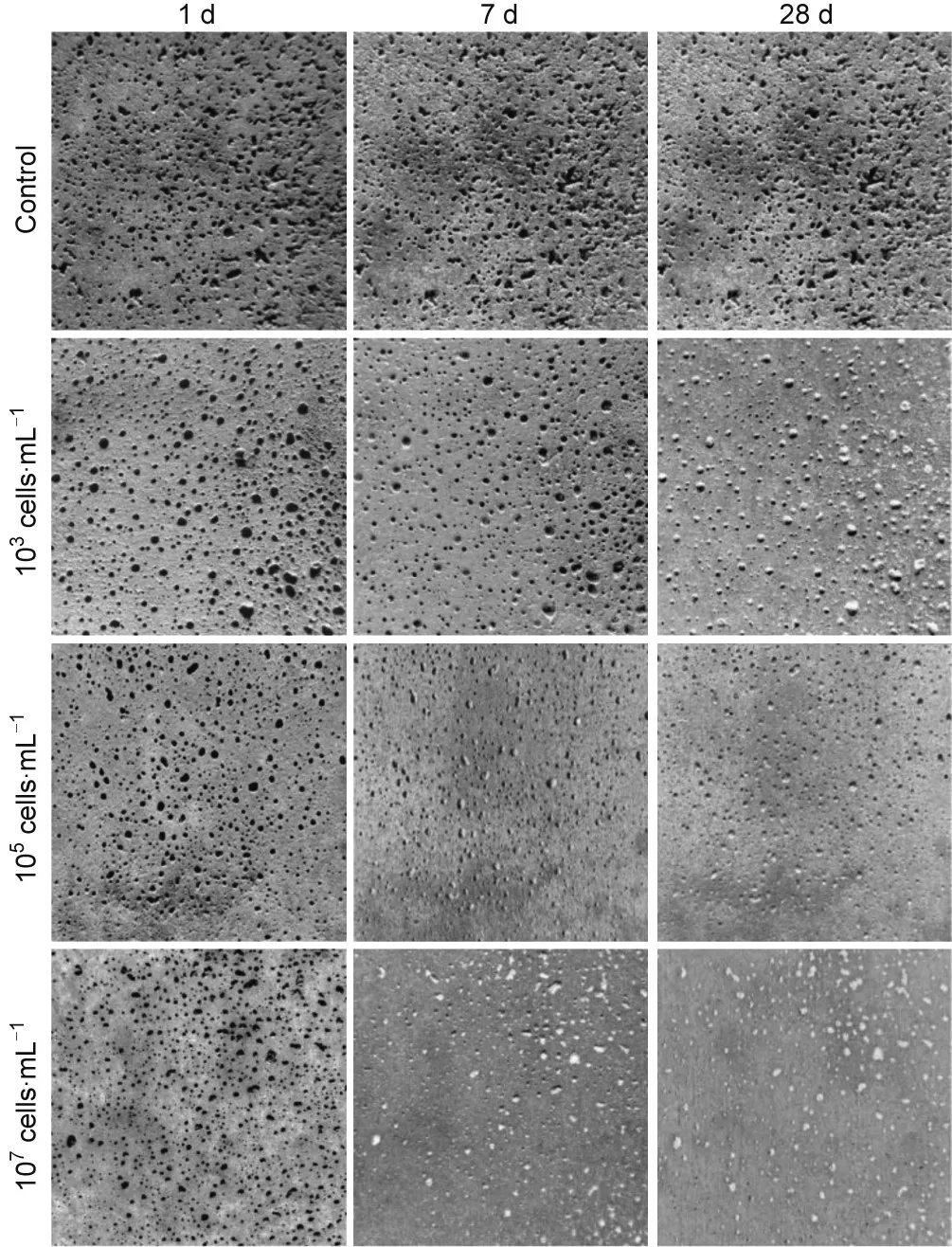
Fig. 4. Surface porosity changes over 28 d with different bacterial concentrations.Each image represents 50 cm×50 cm section. Reproduced from Ref. [62] with permission of Elsevier Ltd., © 2018.
In addition to non-ureolytic bacteria incorporation into concrete structures, these bacteria have also been investigated in terms of surface treatment. Xu et al. [64] reported a reduction of approximately 50% in the water absorption capacity of cement mortars surface-coated with B. cohnii after two weeks of incubation in nutrient media supplied with Ca–L or Ca–Glut. The reduction was attributed to pore blocking by bacterial CaCO3precipitation. The choice of calcium source did not significantly impact the pore blocking action of this surface-coating approach.However,Ca–Glut resulted in a significantly thicker CaCO3precipitation layer (260–360 μm) compared with the precipitation layer resulting from Ca–L (100–140 μm). It was observed that the morphology of CaCO3precipitates was not the same as that of well-crystallized calcite when Ca–L and CaCl2were supplied; it was also not the same as that of vaterite when Ca–Glut was supplied.This variation in crystallinity could be attributed to different organic matters, such as lactate or glutamate, which changed the crystal growth rate along different planes.
Additionally, MICP by cyanobacteria Synechococcus PCC8806 was investigated in a concrete powder solution (pH 11.7) [48];Synechococcus PCC8806 cells remained viable at pH 11.7 after 24 h. Calcium chloride (50 mmol∙L-1) was added as a calcium source to evaluate the effect of cyanobacteria on the concrete environment where Ca2+concentration was evaluated as 4 mmol∙L-1; however, it was not added to a control setup. It was reported that Synechococcus PCC8806 did not impact the carbonation of concrete samples and could not precipitate CaCO3using Ca(OH)2, but it utilized 38% of the available Ca2+after 24 h when calcium chloride was added. The calcification behavior of cyanobacteria was also investigated by adding the cyanobacteria to concrete cubes and immersing the cubes in a calcium chloride solution for 45 d. A precipitated surface layer (200–270 μm thick)on the treated cubes was observed,whereas the abiotic control samples had patches of precipitates with a maximum thickness of 115 μm. The bio-treated concrete cubes absorbed significantly less water (3 g∙cm-2) than the control concrete cubes; however, ureolytic MICP reportedly caused a more drastic water absorption loss (1 g∙cm-2) [19]. The initial incorporation of cyanobacteria as concrete reinforcement yielded promising results. The potential of cyanobacteria in concrete remediation may be better assessed by longer experimental duration, adherence to standard field methodology,and evaluation of widely used mechanical parameters, such as compressive strength. No information regarding the effect of light on Synechococcus-mediated MICP was obtained.A potential area for future research is the possible exposure of this photosynthetic bacteria to varying light levels.Understanding the adaptability of cyanobacteria to perform biomineralization on concrete surfaces can considerably benefit the field CCS because many cyanobacteria species can perform biomineralization under various ecological settings and extreme environmental conditions with the potential to sequester carbon from waste, such as organic waste streams [127].
4.2.2. Crack healing
Crack healing by non-ureolytic bacterial concrete self-healing systems can forestall microcracks from propagating and coalescing into larger cracks,thus preventing the entry of deleterious environmental elements into the structure.The efficiency of non-ureolytic biological concrete crack healing has been assessed by monitoring the crack width over time, the visual and chemical analyses of healing agents, and the changes in water flow and absorption through the cracks.
Wiktor and Jonkers [66] impregnated ECPs with Ca–L, yeast extract, and 1×105cells∙mL-1of B. alkalinitrilicus spores and observed the healing of up to 0.46 mm cracks after 100 d of curing and up to 0.18 mm crack healing in control samples attributed to the autogenous self-healing of cement-based structures.Compared with previous findings [36], the self-healing agent’s immobilization significantly improved the system’s performance.This conclusion was based on the system’s improved functionality, with selfhealing occurring at desirable rates throughout 100 d, whereas the initial study only reported 7 d of functionality.
Other protective carriers have also been investigated in conjunction with non-ureolytic bacteria and have resulted in promising crack healing. Khaliq and Ehsan [61] incorporated 2.8×108cells∙mL-1of B. subtilis spores and Ca–L into the mortar, either directly or immobilized in LWA or GNP. Crack healing was measured on samples that were pre-cracked 3–28 d after preparation.In samples pre-cracked after 3 and 7 d, the highest crack healing was observed after 28 d in mortar specimens containing GNPs with both values slightly exceeding 0.8 mm, followed by samples containing LWA with crack healing of approximately 0.6 mm.Samples containing directly added healing agents exhibited approximately 0.35 mm of healing, exhibiting 0.2 mm more crack closures than the control samples. However, in samples pre-cracked after 14 and 28 d, most crack healing was observed after 28 d in the specimen containing LWA with 0.59 and 0.52 mm healing,respectively.The samples containing GNP and pre-cracked at 14 and 28 d exhibited approximately 0.41 and 0.38 mm healing, respectively. The samples that directly received healing agents and pre-cracked at 14 and 28 d showed 0.21 and 0.15 mm of healing, respectively. A trend is observed between the specimen age when cracked and healing efficiency; the younger specimens pre-cracked at 3 and 7 d showed higher crack healing for all treatment groups than the healing observed in specimens cracked at 14 and 28 d. However, crack healing was more distinct in the specimen containing GNP.The results suggest that LWA was better than GNP in protecting the bacterial spores. The reduction in the survival of B. subtilis with the specimen age is the factor that lowers the system’s functionality. Another study found that the optimal B. subtilis concentration for crack healing when directly added was 107cells∙mL-1[62].Khaliq and Ehsan[61]suggest that GNP particles cannot protect the bacteria from the dense concrete microstructure, which can crush the spores. One possible explanation given by the authors is the susceptibility of GNP to multi-axial loading, which can increase the exposure of healing agents to the cement environment. The technique employed by Khaliq and Ehsan [61] to compare the self-healing efficiency of specimens pre-cracked at different ages clearly illustrated the difference in protective capability between LWA and GNP.The broader adaptation of this technique can aid researchers in comparing the protective potential of different carriers and possibly benefit the research field by providing more distinct contrasts among different systems in shorter incubation times. The visual analysis of the microstructure of different specimens by Khaliq and Ehsan[61]showed more abundant CaCO3crystal formation in samples with higher crack closure rates in agreement with previous reports on bio-concrete crack healing[36,128].
Xu and Yao[65]investigated the crack healing in concrete containing bacterial spores, Ca–L, or Ca–Glut or cured in a solution containing healing agents. The samples with healing agents in the curing solution experienced higher spore germination and required fewer nutrients. In contrast, the spore germination in the specimen containing embedded healing agents was lower and required more nutrients.The self-healing systems successfully healed 0.4 mm-wide cracks. The authors reported the better performance of Ca–Glut than that of Ca–L in terms of crack healing efficiency and regained post-healing flexural strength.They attributed this finding to the higher CaCO3precipitation rates with Ca–Glut, denser crystal structure (especially in the transition zone),and micrometer-long CaCO3layer connecting the cement surface to the main body of CaCO3precipitates(Fig.3[65]).These findings led to the conclusion that Ca–Glut in MICP-based concrete selfhealing systems were applicable.
Tziviloglou et al. [119] investigated the effect of a healing system composed of Bacillus spores, Ca–L, and yeast extract immobilized in LWA. The field-emission scanning electron microscopy(FE-SEM) analysis of the cracks showed larger CaCO3precipitates in the bacterial specimens than those in the control samples after 28 d,with the imprint of bacterial cells distinguishable on the crystal surfaces.Such imprints are a well-documented feature of bacterial biomineralization [129,130]. The observation of the crack surface after 56 d of incubation led to the same conclusion that crystal growth occurred in both bacterial and control specimens;however, smoother crystalline surfaces were observed in the bacterial specimen. The reasons for the disappearance of bacterial imprints is interesting. Moreover, several questions can be raised:Can increasing the distance between the mineral layer and healing agents reduce the bacterial influence on crystal growth? Did biological self-healing cease after 56 d, or was the rate of autogenous self-healing faster than the autonomous self-healing after 56 d?The answers to such questions can aid researchers in improving the prediction of the healing system’s performance in field applications.
Vermeer et al. [97] immobilized B. cohnii-related species in refined wastewater-sourced PHA flakes (0.5–1.0 mm) along with yeast extract and incorporated the non-ureolytic self-healing agent into the mortar specimens.The mortars were cracked after 28 d of curing and incubated under humid (> 95% relative humidity (RH))conditions at room temperature for 56 d;the crack size was in the range 0.46–0.48 mm. A positive control specimen was fabricated using the self-healing system based on the lactic derivative described by Mors and Jonkers [26,131]. After 56 d of incubation,the authors observed virtually complete crack healing in the positive control specimen and mortars containing the PHA-based nonureolytic self-healing system.In contrast,the crack in the negative control specimen was only partially healed. The environmental scanning electron microscopy (ESEM) observation of precipitates showed larger CaCO3crystals on the crack surface of specimens containing self-healing agents. The precipitates observed in the bacterial samples contained rod-shaped imprints of bacterial cells similar to those reported by Tziviloglou et al. [119], who only observed the imprints after 28 d; the imprints disappeared after 56 d. This difference may have arisen from the various selfhealing efficiencies of systems or may have resulted from the different curing conditions employed.
To overcome the gap between the typical laboratory conditions used in biological self-healing concrete research and environments that host a considerable portion of cement-based structures, a study [113] assessed the self-healing potential of calcium alginate-immobilized Bacillus halmapalus alongside yeast extract and mineral precursors for low-temperature marine environment applications.The extension of the application of non-ureolytic bacterial concrete self-healing systems to the marine environment is an important step, considering the susceptibility of reinforced structures to chloride and sulfate attacks. The results of the crack-healing assessment did not present significant variation in the extent of crack closure between the control and bacterial mortars. In the bacterial mortars, limited precipitation was observed around the crack edges, similar to those observed in the control samples. In bacterial mortars, the precipitates were observed to bridge the crack in certain spots. The energy-dispersive spectroscopy elemental analysis of precipitates revealed the presence of Mg-based and Ca-based precipitates around the crack and on the capsule surface, respectively. It was noted that the swelling capacity of hydrogel alginate capsules (up to 3 mm) [114] might contribute to the blocking of larger cracks; the authors reported a reduction in the 28 d compressive strength. This phenomenon has been attributed to the softness of alginate capsules relative to the cement matrix and the formation of pores in the cement matrix around hydrogel capsules. The latter was assumed to be a result of the excessive water uptake from the matrix to the hydrogel, possibly during the secondary cement hydration stage [121].
Similarly, our research team previously investigated the crack healing potential of B. pseudofirmus and nutrient broth encapsulated in calcium alginate capsules by incubating the cement paste and mortar specimens under wet–dry conditions for 56 d [20].Artificial cracks with a width of up to 350 μm were introduced.All cracks were eventually sealed in biological and abiotic specimens; those in the biological specimens were fully sealed earlier(28 d), whereas the cracks in the control specimens were sealed after 56 d. The visualization of the cement matrix using FE-SEM revealed larger and more numerous calcite crystals in the biological specimens, especially in the mortar specimens, than in the paste samples. Although the accelerated healing of cracks sufficiently small for autogenous healing is interesting, the system’s self-healing potential must also be studied with wider cracks.The wet–dry incubation regimen may have interesting effects on the samples;these can include the alternating O2and CO2concentrations and the accentuation of self-healing that results from biological and autogenous concrete self-healing. However, this technique with constant water renewal can result in Ca(OH)2loss,lowering the self-healing efficiency [132]. Gao et al. [60], who induced millimeter-wide cracks in concrete specimens with chitosan-coated calcium alginate capsules containing B. pseudofirmus spores, reported the complete healing of a 1 mm wide crack after 28 d.
Zhang et al. [133] investigated the potential of mixed bacterial cultures for concrete self-healing.They tested AE,facultative aerobic(FA),and AN consortia,which were selected based on their ability to precipitate CaCO3with Ca–L as the primary nutrient source.To assess the effect of these three consortia on concrete,ECPs were impregnated with each consortium and appropriate nutrients: for AE, 8 g∙L-1of Ca–L and 1 g∙L-1of yeast extract; for FA and AN,4 g∙L-1of Ca–L, 4 g∙L-1of Ca–N, and 1 g∙L-1of yeast extract. The ECP containing each consortium was added to separate concrete samples,whereas the control sample did not contain added bacteria.Cracks ranging from 0.1 to 0.9 mm were produced in the samples and re-examined after 28 d of curing. The fastest healing rate was reported by the AE consortium(0.36 mm),followed by the AN consortium(0.33 mm) and FA consortium(0.28 mm). The authors suggested that this observation might not reflect the FA consortium’s true potential because the environment might not have been sufficiently harsh for optimal CaCO3precipitation. However,the results demand more in-depth research on the application of bacterial consortia.Moreover,the incubation under realistic conditions may highlight the consortia’s hypothetical advantage in resisting environmental fluctuations compared with pure-culture self-healing treatments. The researchers identified the predominant genus present in each consortium and found that Citrobacter and Aeromonas were enriched in the AE consortium,whereas Pseudomonas and Azotobacter have been enriched in the FA and AN consortia.Most non-ureolytic MICP research applications for concrete reinforcement and repair have focused on the oxidation of organic salts. Denitrification and photosynthesis have also been investigated for potential concrete remediation.Ers¸an et al.[74]produced self-protected granules containing denitrifying bacteria and organic matter; Ca–F, Ca–N, and dried granules were added to the cement mixture. A healing rate of approximately 90% of 0.5 mm cracks in 28 d by CaCO3precipitation was observed. The autogenous healing of 200–250 μm cracks occurred in the control specimen with no granules. Samples containing granules had lower porosity than the control cement samples.The authors found that only approximately 15% of nitrate ions dispersed in the cement matrix were available to bacteria. Further, the control specimen containing Ca–F and Ca–N experienced more autogenous healing than without.This is expected because Ca–F and Ca–N are common concrete additives that improve mechanical properties and act as antifreeze agents.These findings indicate the application potential of denitrifying bacteria. Ers¸an et al. [25] continued by immobilizing the denitrifying bacteria in ECP and granulated activated carbon (GAC) particles. The authors reported that the maximum widths of fully healed cracks are approximately 370 and 480 μm after 28 and 56 d of incubation, respectively. The treatment involving immobilized D. nitroreducens outperformed the treatment by P. aeruginosa, achieving significant crack healing in 14 d compared with the latter’s 28 d. The crack micrographs are shown in Fig. 5 [25].
4.2.3. Crack water tightness
The assessment of water flow through fresh and healed cracks provides additional information regarding the efficiency of nonureolytic biological concrete crack healing, such as healing in the invisible crack depths and the resistance of healing products to water flow.Tziviloglou et al.[119]investigated the effect of a healing system composed of Bacillus spores, Ca–L, and yeast extract immobilized in LWA. Crack healing was assessed based on alterations to the water flow through cracks or regained water tightness(RWT) with mortar samples treated under two different curing conditions: ①wet curing and ②wet–dry cycles. No considerable difference was observed in the RWT of mortar samples with a healing system and that of the control specimens when the samples were cured under wet conditions for 56 d.The control and bacterial samples obtained approximately 80% and 90% RWT gains, respectively. However, the wet–dry incubation resulted in considerably different RWT results. The control specimens did not exhibit RWT gains and even experienced increased water flow through the cracks,whereas the bacterial specimens gained 98%RWT after 56 d. The authors explained that the RWT achieved by nonureolytic self-healing agents during wet–dry cycles was due to the higher O2levels in the cracks, possibly promoting bacterial action.
Vermeer et al. [97] compared the water flow rate through the cracks of the mortar samples before and after the self-healing cycle; they observed flow rate reductions of 55%, 81%, and 88%for the negative control, positive control, and PHA specimens,respectively.These observations are comparably similar to reports regarding the water flow through healed cracked concrete specimens,which cannot achieve a total crack seal and may be apparent from the visual inspection of fully healed cracks [113,119,134].
Similarly, Mors and Jonkers [26], who incorporated lactic acidbased flakes containing non-ureolytic bacterial spores into the mortar, compared the water flow rates among mortars incubated in water baths or humid environments (> 95% RH). The mortars cracked after 28 d of curing. It was observed that the control and bacterial mortars containing cracks < 0.2 mm in width were fully sealed after incubation in a water bath or under humid conditions;however, only the bacterial samples regained the seal after the cracks were reopened.The corresponding crack widths were elucidated based on the new water flow rate, and the incubation was repeated. It was observed that after the samples were dried, the bacterial samples incubated in the water bath maintained 95%water tightness, whereas that of the bacterial specimen incubated under humid conditions was 65%. A crack healing threshold of 0.7 mm for the self-healing system was observed. Furthermore,the cyclic WTR efficiency of the healing system was reportedly hindered after the fourth cycle.The difference between the flow resistance of precipitates formed in the water bath and those crystallized under humid conditions must be investigated further.The X-ray diffraction analysis of precipitates may reveal the structural variations resulting in the reported 30% difference in flow rates after the mortars were dried. The analysis of the attachment between precipitates and different phases of the mortar through nanoindentation examination may also be beneficial in the investigation [65,135].
Palin et al. [113] assessed the water permeability of mortars containing SAP-encapsulated non-ureolytic self-healing agents.The mortar samples were cured for 28 d and cracked and incubated at 8°C in artificial seawater.The alginate-based system was previously assessed in vitro for applicability to a cold marine environment [114]. The permeation of water through mortar cracks and the crack healing ability of mortars were measured. After 56 d of incubation,the authors reported a 95%reduction in the water permeability of bacteria samples with 0.4 mm wide cracks and a 93%reduction in bacterial samples containing 0.6 mm wide cracks.The reduction in the permeability of bacterial mortars was approximately 30%higher than that recorded for control mortars.Further,it was reported that the specimens containing beads had larger variations in permeability before and after drying than the control mortars.

Fig.5. Photomicrographs showing evolution of cracks after 28 and 56 d of water immersion:(a)reference mortar;(b)abiotic control;(c)mortar containing D.nitroreducens loaded in GAC particles; and (d) mortar containing P. aeruginosa loaded in GAC particles (given values represent average width of shown crack ± standard deviation).Reproduced from Ref. [25] with permission of Elsevier Ltd., © 2016.
Ers¸an et al. [25] continued by immobilizing the denitrifying bacteria in ECP and GAC particles.Cement samples containing biological healing agents recovered 85% more water tightness in the cracks than the abiotic controls after 56 d of incubation. The RWT assessment highlighted an important difference between the non-ureolytic autonomous self-healing and autogenous selfhealing of abiotic samples. Autogenous self-healing had merely sealed the crack mouth,leaving the inner crack surface exposed to water flow.In contrast,the non-ureolytic biological healing agents resulted in calcite and aragonite precipitation along the inner crack walls, considerably reducing the water flow through the cracks.The abiotic samples contained abundant hydration products, such as ettringite and C–S–H. The results emphasize the importance of CO32-production by bacterial metabolism for the healing of deep cracks in which low CO2concentrations may limit autogenous self-healing [136]. This property can aid in protecting the steel reinforcements and the structure’s core from degradation. The findings highlight the potential of AN oxidation of organic carbon sources (e.g., Ca–F) by nitrate reduction in bio-concrete research.Certain non-ureolytic bacterial concrete self-healing systems are summarized in Table 1 [25,61–66,120,133,137,138].
5. Genetic modification of non-ureolytic bacteria for selfhealing concrete
Genetic manipulation techniques can expand the functional arsenal of microbes by providing the underlying code for a previously absent cellular machinery. Sarkar et al. [139] targeted a silica-leeching protein previously reported to enhance the mechanical properties of concrete [140]. The authors transformed an Escherichia coli (E. coli) bacterial strain with the silica-leaching gene that expresses a bioremediase-like protein.The incorporation of the genetically transformed E. coli into mortar improved the compressive strength by 30%;a silica-containing mineral identified as gehlentine was observed in the matrix. As E. coli does not form spores, only vegetative cells that survived a week were added to the mortar. The authors then transferred B. subtilis with the same gene to examine the silica-leaching protein’s function with spore-forming bacteria [141]. The incorporation of the modified B. subtilis in mortars improved the mechanical strength and crack healing up to 28 d(Fig.6[141]).This improvement was attributed to the enhanced micropore filling, increased matrix compactness,and stronger ITZ.
The above projects demonstrate the potential of genetic engineering for biological self-healing concrete. Considering biomineralization,cells could be equipped with proteins that can precipitate different minerals(as previously described)or more complex mineral forms similar to those observed in multicellular organisms(e.g.,teeth and bones)formed through biologically controlled mineralization (BCM). Mineral formation through BCM is strictly controlled by various proteins, which may be used in the construction sector in the future. Furthermore, entire metabolic pathways can be inserted, or existing pathways can be modified for efficiency and rate.Such prospects rely on proteomics research,which involves the assignment of a specific role to a protein; this may be a function that other researchers can subsequently use.
6. Feasibility of non-ureolytic bacterial concrete self-healing
The non-ureolytic self-healing systems described in Sections 3 and 4 have improved the physical and mechanical propertiesof concrete/cement/mortar samples through MICP. These systems include agents that can achieve self-healing under varying oxygen levels using a host of precursors and a diverse set of bacterial species. Here, the feasibility of implementing nonureolytic bacterial concrete self-healing systems in terms of sustainability, the attempts to use the technology outside of the laboratory, and the remaining challenges and future directions are discussed.
Table 1 Non-ureolytic bacterial concrete self-healing systems.
6.1. Sustainability
Although concrete itself has a relatively low environmental impact, the constantly increasing demand for its use has resulted in an ever-growing global problem. Concrete self-healing systems are intrinsically sustainable. However, all facets of sustainability must be considered to ensure that biological self-healing concrete applications can be upgraded from the laboratory to global applications. As further discussed, the three main pillars of sustainability—environmental, economic, and social sustainabilities—are interconnected when the goal is the widespread use and the social acceptance of biological self-healing concrete.
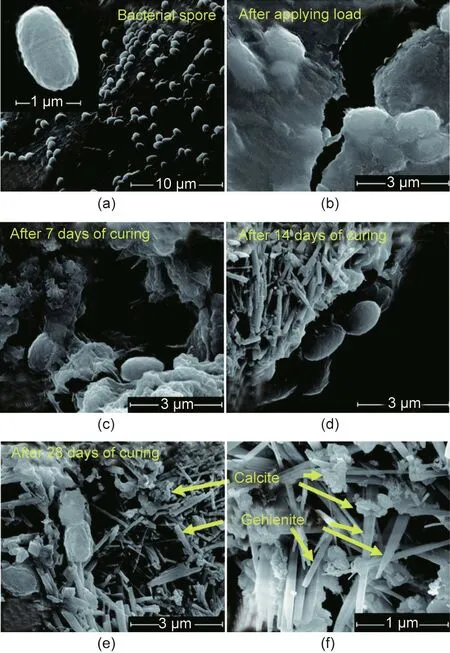
Fig. 6. FE-SEM images of progress of gehlenite formation inside mortar with transformed B. subtilis on different days. (a) Dormant bacterial spore; (b) bacterial spores next to a load-induced crack;(c)the cement matrix after 7 d of curing;(d)the cement matrix after 14 d of curing; (e, f) the cement matrix after 28 d of curing.Reproduced fromRef.[141]with permissionof The RoyalSocietyofChemistry,©2015.
Non-ureolytic concrete self-healing systems expand the sustainable nature of concrete self-healing products by employing various cost-reducingand waste-reducing measures.Increasing the sustainability of concrete through better autogenous self-healing has been demonstrated by incorporating industrial and agricultural wastes,which either act as cement replacement and/or enhance the physical and mechanical properties of concrete and extends the service life of structures. Biological concrete self-healing systems serve similar purposes but can function over extended periods and undergo various cycles.The implementation of a controlled-release mechanism by nanomaterials,such as nanoclays,multicore capsules,or multichannel fibers,could be a future solution to extend the service life of non-ureolytic bacterial concrete self-healing systems [142–146]. Pozzolanic or expansive nanoclays, such as halloysite nanotubes,montmorillonite,and kaolinite,have been extensively studied as sustainable carriers with a controlled-release property.Moreover, they have shown compatibility with cement structures by improving the mechanical properties of concrete and its resilience to alkali–silica reaction and crack formation [143,147–153].Clay particles used in biological self-healing concrete systems may benefit from a protective layer where the pozzolanic reaction of cement with clay materials may be disadvantageous or where additional protection is required for biological healing agents[111,154].
Furthermore, non-ureolytic biological self-healing systems can also incorporate various waste products,such as bacterial nutrients,calcium sources,or carrier materials,as discussed in Section 3.1.The integration of locally sourced waste as healing agents or carriers can lower the construction industry’s CO2footprint and the cost of selfhealing products [155,156]. The prospect of simultaneous CO2sequestering and CaCO3precipitation by bacterial concrete selfhealing systems can also move the systems closer to commercialization. However, this requires an in-depth investigation to evaluate the factors that impact CaCO3precipitation. In addition to CO2sequestering through bacterial action,the use of CO2-capturing carrier materials,such as biochar,can add to the sustainable nature of concrete self-healing products[157,158].
Moreover, using local resources can result in employment gain for local labor and talent. The construction industry is anticipated to be the primary market in developing countries[159].Such fastgrowing economies can be a launchpad for education and social integration regarding novel materials and construction approaches.The production and distribution of self-healing concrete systems can be an industry that can follow the anticipated growth of the construction sector.This is possible provided that sufficient support for local industries and businesses is available in terms of investment,tax deductions from the government for environmentally friendly efforts,positive and widespread media coverage,and social acceptance of the sustainability goals. Although biological self-healing systems generally exhibit positive impacts by increasing durability and improving structural integrity, they are insufficient to gain social acceptance and economic favor.
6.1.1. Economic feasibility
The economic viability of bio self-healing systems can only be achieved after the systems have been applied and tested on a large scale using a uniform assessment methodology and studied in the long term to gain the trust of the construction industry’s decisionmakers [160,161]. These efforts can also showcase the impact of such systems on construction speed and provide a clear picture of the cost of such applications. They can further demonstrate how construction workers with limited to moderate knowledge of self-healing systems can work with the products, assess the quality of the fresh concrete mix containing self-healing system,and even identify errors or problems associated with the application of self-healing system. Large-scale pilot projects can be considerably enhanced with support from government bodies, nonprofit organizations, and other environmentally active entities.Such support can improve public perception and knowledge regarding concrete and the efforts that have been devoted toward developing a more environmentally friendly product. The largescale implementation of biological concrete self-healing systems may benefit from pairing with novel construction techniques(e.g., three-dimensional printing of concrete), which have been used to construct various structures[162].Other areas of construction, such as underground construction (which poses challenges that are different from those encountered in urban structures[163]) and cemented dam construction [164]. These structures may also be resilient against specific destructive forces, such as seismic structures engineered to withstand earthquakes [165]. A notable example is the use of a chemical concrete self-healing system for tunnel engineering [166].
To date, the long-term performance of concrete self-healing products remains a matter of speculation. Nevertheless, this can be investigated in large-scale pilot projects using concrete selfhealing systems that can be analyzed after years of environmental exposure. Non-ureolytic bacterial concrete self-healing systems have been applied to an aging Dutch parking lot[167]and a water canal in Ecuador [30]. The water canal’s reported field condition(5 °C and 2000 m above sea levels) can lead to self-healing fluctuations compared with laboratory assessments,further highlighting the importance of field studies in this area of research. Low temperatures can drastically reduce bacterial activity and the solubility of calcium salts, such as Ca–L [168]; high altitude corresponds to lower atmospheric pressure. Hence, lower concentrations of gases, such as O2and CO2, can hinder bacterial activity and retard the physical properties of concrete. Wiktor and Jonkers[167] treated the surface of a parking lot located in a coastal area and subjected it to freeze–thaw cycles with non-ureolytic healing agents. It significantly sealed heavily leaking concrete cracks and improved the freeze–thaw resistance of the concrete structure.The addition of B. pseudofirmus-containing lightweight perlite aggregate to panels placed alongside a highway in Wales, United Kingdom proved challenging compared with the panels containing abiotic healing agents in terms of producing the same quantity of healing agents required in handling and casting [169]. The spores and nutrients were immobilized on separate perlite particles, and the panel was designed with a feeding network.Overall,the bacterial panel showed a slightly lower cracking load and strength than the control samples; however, the parameters were in an acceptable range. Such studies, accompanied by reports of long-term effects, can clarify the unanswered questions regarding the longterm impact of MICP on concrete structures and the functionality of biological self-healing systems.
Assessing the impacts of non-ureolytic self-healing concrete systems under realistic conditions requires non-destructive monitoring and detection techniques that are seldom required in laboratory measurements. Non-destructive tests to monitor various civil and historic structures have been established [170–172];however, no such example has yet been presented regarding biological self-healing concrete.Although these challenges are present in the precise determination of the viability of biological healing agents under laboratory conditions, the primary purpose of this research field must be given forethought. The in situ application of biological concrete self-healing systems must be accompanied by non-destructive tests that can provide information on the structure’s health and the state of biological components. Such revolutionary techniques may emanate from novel advancements in concrete monitoring,such as machine learning and artificial intelligence [173]. In the medical context, theranostic systems have been developed to deliver therapeutic agents with the inclusion of a detection function, thus enabling simultaneous therapy and detection [174]. As biomimicry is the basis of biological selfhealing concrete, the future of this field may include systems that operate based on a similar principle. The incorporation of detectable elements,such as nano-metallic particles or components with strong fluorescent signals coupled with advanced detection methods, could be the future solution to the question of the long-term applicability of biological self-healing concrete.
The production cost of biological concrete self-healing systems has been previously identified as a challenge[87,88].In this regard,non-ureolytic self-healing systems have included various costreduction strategies.Non-ureolytic systems can rely on several different substrates (different organic calcium salts) sourced from various waste materials, as discussed in Section 3.1.2. The same waste products can also be assimilated into ureolytic bioconcrete systems as calcium sources alongside urea. Establishing a deeper understanding of the nutritional requirements of bacteria and the bioavailability of nutrients during the self-healing process have been suggested as a strategy for improving bacterial growth and can result in better self-healing performance and economic feasibility[112,175].The potential of this approach can be assessed by future research. Using mixed bacterial cultures has been proposed as a substantial cost-reducing option for concrete selfhealing products because non-sterile spore production from nonpure bacterial cultures is reportedly cheaper by more than 70% in contrast to using sterile pure spore cultures [87]. Mixed bacterial cultures can simultaneously incorporate ureolytic and nonureolytic bacteria [67,158,176]. Future research can test the efficiency and limitations of different MICP pathways as they cooccur under mixed culture conditions.
In addition to lower production costs, mixed cultures allow a single self-healing system to operate under various environmental conditions by performing a combination of metabolic processes not possible or expected from a single bacterial species, leading to improved cell survival MICP performance [67,177,178]. Using mixed bacterial cultures for concrete self-healing can widen the research prospects in this field as such cultures can drastically behave differently under various conditions, shifting from a cooperative relationship to a competitive state[179].Hence,examining the mixed culture behavior in the concrete environment by studying interspecies interactions under different conditions can clarify the potential of such systems or concrete self-healing. Moreover,the immobilization of mixed cultures as separate pure cultures or as a single mixed culture may provide insight into their concrete self-healing potential. Future research can focus on factors, such as the effect of different environments on mixed culture behavior,the optimal number of species and individual population levels,the degree of contact among different species,and the immobilization techniques suitable for mixed culture concrete self-healing systems. Researchers must also devise quality control protocols for mixed cultures. The dynamics of mixed culture population can change because of several reasons,such as oxygen availability or bacterial preference to conditions(e.g.,salinity or temperature).Such conditions may result in the overgrowth of some species,whereas other bacteria might struggle to function and maintain their population levels. Based on the MICP and concrete selfhealing results obtained when mixed cultures and combined calcium sources have been used [133], future research on different bacterial combinations and calcium substrate mixtures can further determine the applicability of such systems.
The environmental range of applicability of urea-dependent and urea-independent concrete self-healing systems depends on the metabolic requirements of the bacteria used. Typically, nonureolytic systems that can operate under a broad range of conditions as non-ureolytic MICP pathways (e.g., oxidation of organic salts and denitrification)are AE and AN.However,certain ureolytic bacteria can also precipitate CaCO3when deprived of oxygen[180].Overall, non-ureolytic bacterial concrete self-healing systems are seemingly more sustainable and widely applicable than ureabased bio-concrete products and thus deserve more attention from researchers.Silva et al.[87]emphasized that the added cost of bioconcrete products can be justified by the increased service life of structures and the environmentally friendly nature of such products, especially in eco-turned markets. The application of nonureolytic bacterial concrete self-healing systems is undoubtedly a step in that direction.
Another approach for cheaper production of non-ureolytic spores is the nitrification of granules composed of bacteria and organic matter [181]. The developed granules are self-protecting and rely on established concrete additives (i.e., Ca–N and Ca–F)as primary nutrient sources.These qualities further reduce the production cost of the healing agents, providing a similar degree of concrete self-healing as that of immobilized bio-concrete products[74]. The granules were reportedly cultivated for two months;however, they represent a healing agent that is approximately 95% cheaper to produce than the ureolytic system analyzed by Silva et al. [87] in terms of production cost.
6.1.2. Social awareness: Health concerns
The safety of bacterial products that could be used in residential spaces in the future must not be a matter of speculation because public trust is a crucial growth factor.Although most bacteria used in biological self-healing concrete are not disease-causing or pathogenic,specific examples are worth mentioning.B. sphaericus,a widely used ureolytic strain in bio-concrete research, can cause infections in immunocompromised individuals [182]. Such risks may hamper public trust in bio-concrete products, limiting the application to sensitive environments,such as hospitals.However,health risks can also be associated with certain non-ureolytic strains (e.g., B. subtilis) used for bio-concrete research; specific B.subtilis species can cause food poisoning [183]. The possibility of ailment does not mean that B. subtilis species cannot be used for bio-concrete research; it simply means that health hazards must be thoroughly investigated before large-scale application is implemented. Moreover, researchers must ensure that the bacterial strain used does not bear pathogenic genes. Further assurance can be obtained by considering the compatibility of biological healing agents with commercial livestock and determining whether any interactions may arise between the healing agents and specific additives used in the construction of bio-stable farms [184].
Another health-related problem is the use of bacterial consortia.Although researchers who utilize such microbial mixtures have reported the predominant bacterial species that make up the mixture, additional safety checks are necessary to determine whether any pathogenic microbes are present in the mixture to ensure product safety.
As discussed,genetically modified bacteria have shown promising results. The antibacterial resistant gene is often included to select the transferred bacteria against the non-transferred wildtype bacteria during the genetic transfer procedure. As the spread of antibacterial resistance is a severe and fatal problem, researchers should rethink this genetic manipulation strategy when bacteria are intended to be used in residential spaces; other selection strategies, such as metabolic selection, are available.
6.2. Limitations
Unfortunately, most non-ureolytic self-healing reports were conducted without protecting the healing agents with a carrier or capsule. Although such treatments can still improve physical parameters under controlled laboratory conditions, the healing agents are unnecessarily sacrificed to harsh mixing conditions and inhospitable cement environments.Consequently,these treatments barely contribute to the advancement of the field.The direct addition of spores and vegetative bacterial cells can lead to the release of organic materials that can retard cement hydration and damage the overall physical properties of the cement matrix.Similarly, this can occur when yeast extract is directly added to the cement. Moreover, most vegetative cells or spores cannot survive the mixing process, but this is not always the case. Chaurasia et al. [185] focused on the impact of nutrient-starved and wellnourished ureolytic and non-ureolytic bacteria on the cement’s microstructural properties and suggested that bacteria could be involved in pathways other than the usual MICP pathways mentioned in Section 2 that can improve the cement matrix.
Reports on the effects of environmental conditions on concrete self-healing are scarce [186]; no reports are available on such effects on biological self-healing agents.Although the bacteria used in this field are versatile and vigorous organisms, the rate and impact of their self-healing action can be significantly altered by environmental conditions. Therefore, the in-depth analysis of this topic could set specific standards for applying biological healing agents in different environments. Furthermore, the impact of global warming should be considered.
6.3. Alternative applications
The interplay between global warming, accelerating urbanization, agricultural deforestation, and other unsustainable agricultural practices, among other factors, are responsible for the increase in desert areas.Two major sustainable development goals are counteracting desertification and land degradation [187,188].Ureolytic and non-ureolytic MICP-capable bacteria have been used as sand and soil solidifying agents[189–191];hence,their use is an environmentally friendly means to overcome the abovementioned growing ecological threat. The main challenge in using a bacterial solidification strategy is the considerable quantity of healing agents required, especially the calcium source. The investigation of the potential of concrete self-healing agents with bacterial species—such as B.subtilis,which can grow and perform MICP at neutral pH values—could reduce the required quantity of solidifying agents and increase the solidification efficiency by localizing the solidifying agents.
Releasing NH3to aquatic environments is dangerous because it can hinder the application of ureolytic self-healing concrete systems to marine environments [192] and structures for ecological purposes, such as artificial reef construction. Using self-healing concrete systems instead of typical concrete structures can be advantageous to artificial reefs because they can accelerate CaCO3deposition, which has been shown to improve coral longevity[193,194]. Moreover, the presence of CaCO3on the surface of artificial reefs is generally recommended [195,196]. Considering the range of organisms that can potentially benefit artificial reefs(e.g., photosynthetic cyanobacteria or electroactive bacteria), the prospects of using non-ureolytic bio-concrete systems for such applications are broad.
7. Conclusions
Non-ureolytic bacterial concrete self-healing agents have been proven as viable alternatives to ureolytic systems. Moreover, they allow the implementation of cost-reducing strategies (e.g., incorporating waste materials and using mixed bacterial cultures) to potentially commercialize biological concrete self-healing systems.Non-ureolytic MICP pathways remove the environmental burden posed by ureolytic MICP by offering environmentally friendly options that can be active under various conditions using various substrates. Non-ureolytic bacterial mixtures can perform simultaneous metabolic activities,which are not possible by pure bacterial cultures; moreover, they can tolerate drastic environmental changes. Mixed-culture bio-concrete systems can also include a mixture of calcium substrates,which could be sourced from industrial waste. The concrete self-healing potential of non-ureolytic pathways, such as photosynthesis, is generally untapped but presents positive prospects. Field studies are scarce, and most selfhealing analyses have been conducted without considering the impact of fluctuating field conditions. The long-term performance of biological concrete self-healing systems remains speculative.Future advances in assessing the long-term performance of nonureolytic biological concrete self-healing systems and their largescale application could be combined with advances in monitoring and evaluation methods.These methods are anticipated to provide information regarding the health of the structure and the state of biological healing agents. Overall, non-ureolytic bacterial concrete self-healing systems represent the future of bio-concrete research.They can be potentially affordable, sustainable, and applicable to various environments and uses.
Acknowledgments
We extend our gratitude to Monash University for funding this project and to our colleagues Ms. Sheik Ambarine Banon Auckloo,Ms. Khanisya Palaniandy, and Ms. Febrianne Sukiato for support and proofreading.
Authors’ contribution
Mohammad Fahimizadeh, Pooria Pasbakhsh, and Lee Sui Mae conceptualized and designed the study. Mohammad Fahimizadeh did the literature review and wrote the main draft of the paper,while Pooria Pasbakhsh reviewed the main draft and restructured the paper. Mohammad Fahimizadeh, Pooria Pasbakhsh, Lee Sui Mae, R.K. Singh Raman, and Joash Ban Lee Tan drafted and improved the revised paper. All authors critically revised the manuscript and gave final approval for the version to be published.All authors agree to be accountable for all aspects of the work in ensuring that questions related to any part of the work are appropriately investigated and resolved.
Compliance with ethics guidelines
Mohammad Fahimizadeh,Pooria Pasbakhsh,Lee Sui Mae,Joash Ban Lee Tan, and R.K. Singh Raman declare that they have no conflict of interest or financial conflicts to disclose.

- Engineering的其它文章
- Electric Air Taxis Create Megadeal Buzz
- Tissue Engineering and Regulatory Science
- Factors Predicting Progression to Severe COVID-19: A Competing Risk Survival Analysis of 1753 Patients in Community Isolation in Wuhan,China
- A Vaccine Based on the Receptor-Binding Domain of the Spike Protein Expressed in Glycoengineered Pichia pastoris Targeting SARS-CoV-2 Stimulates Neutralizing and Protective Antibody Responses
- Fabrication and Applications of Multi-Fluidic Electrospinning Multi-Structure Hollow and Core–Shell Nanofibers
- Past and Future Changes in Climate and Water Resources in the Lancang–Mekong River Basin: Current Understanding and Future Research Directions