
A Vaccine Based on the Receptor-Binding Domain of the Spike Protein Expressed in Glycoengineered Pichia pastoris Targeting SARS-CoV-2 Stimulates Neutralizing and Protective Antibody Responses
2022-08-17 01:51BoLiuYingYinYuxioLiuTintinWngPengSunYngqinOuXinGongXuchenHouJunZhngHonggungRenShiqingLuoQinKeYongmingYoJunjieXuJunWu
Engineering 2022年6期
Bo Liu, Ying Yin, Yuxio Liu, Tintin Wng, Peng Sun, Yngqin Ou, Xin Gong,Xuchen Hou, Jun Zhng, Honggung Ren, Shiqing Luo,e, Qin Ke,e, Yongming Yo*,Junjie Xu,*, Jun Wu,*
a Department of Microorganism Engineering, Beijing Institute of Biotechnology, Beijing 100071, China
b Medical Innovation Research Division & Fourth Medical Center of the Chinese PLA General Hospital, Beijing 100853, China
c Department of Neurosurgery, First Medical Center of the Chinese PLA General Hospital, Beijing 100853, China
d Shenzhen Taihe Biotechnology Co. Ltd., Shenzhen 518001, China
e Institute of Physical Science and Information Technology, Anhui University, Hefei 230000, China
Keywords:Coronavirus SARS-CoV-2 Vaccine Yeast Receptor-binding domain (RBD)
ABSTRACT In 2020 and 2021, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), a novel coronavirus,caused a global pandemic. Vaccines are expected to reduce the pressure of prevention and control, and have become the most effective strategy to solve the pandemic crisis. SARS-CoV-2 infects the host by binding to the cellular receptor angiotensin converting enzyme 2(ACE2)via the receptor-binding domain(RBD)of the surface spike(S)glycoprotein.In this study,a candidate vaccine based on a RBD recombinant subunit was prepared by means of a novel glycoengineered yeast Pichia pastoris expression system with characteristics of glycosylation modification similar to those of mammalian cells. The candidate vaccine effectively stimulated mice to produce high-titer anti-RBD specific antibody. Furthermore, the specific antibody titer and virus-neutralizing antibody (NAb) titer induced by the vaccine were increased significantly by the combination of the double adjuvants Al(OH)3 and CpG. Our results showed that the virus-NAb lasted for more than six months in mice. To summarize, we have obtained a SARS-CoV-2 vaccine based on the RBD of the S glycoprotein expressed in glycoengineered Pichia pastoris,which stimulates neutralizing and protective antibody responses. A technical route for fucose-free complex-type N-glycosylation modified recombinant subunit vaccine preparation has been established.
1. Introduction
The global pandemic caused by the severe acute respiratory syndrome coronavirus 2(SARS-CoV-2)has resulted in the infection of more than 99 million people and the deaths of 2.13 million people worldwide until 25 January 2021 [1]. SARS-CoV-2 binds to the host angiotensin converting enzyme 2 (ACE2) receptor via the receptor-binding domain (RBD) region of the spike (S) protein[2]. A change in the conformation of the S protein triggers membrane fusion,permitting virus invasion into cells.Studies have shown that RBD-specific antibodies can bind to either the upstream or downstream conformation of the RBD or to structures other than the ACE2 binding site, thereby blocking the binding of the RBD to ACE2 [3]; this makes the RBD an ideal antigen for neo-coronavirus vaccines[4,5].In vaccine design,therefore,inducing the immune system to produce antibodies against the RBD region in the S glycoprotein may be effective in preventing virus infection.
The RBD contains two glycosylation sites,N331 and N343[6].At present, vaccines directed to the S protein or the RBD protein as antigens are produced by insect cells or mammalian cells [4,5].The production cycles of these two vaccine platforms are relatively long and have a relatively high cost. Yeast is an ideal proteinpreparation platform,as it is characterized by fast growth,low production cost,and rapid production[7].Yeast also has the ability of protein post-translational modifications, such as N-glycosylation[8]. However, N-glycosylation in yeast leads to hypermannosylation, which will cause protein epitope masking and high-level structural changes, thus affecting the immunogenicity and protective effect of the vaccine. Glycosylation engineering has led to the production of glycoengineered Pichia pastoris yeast,which performs glycosylation similar to that of mammalian cells while retaining the advantages of fast growth, low cost, and rapid reaction [9,10]. The H7N9 hemagglutinin glycoprotein vaccine expressed by glycoengineered yeast has been shown to have good immunogenicity and protective effects [11].
Over the course of transmission, the virus continues to mutate,and these mutations confer various advantages. For example, the D614G mutation of the S glycoprotein increases the infectivity of the virus, and the N493K mutation in the RBD domain enhances the affinity of the RBD with the host ACE2 receptor by adding a pair of salt bridges [12]. The N493K mutation also reduces the binding capacity of the RBD with several monoclonal antibodies(including REGN10933, REGN10987, LY-CoV555, and s309, which are in clinical trials),thus enabling immune escape.An effective and safe vaccine is the most promising means of preventing the spread of the virus. The glycoengineered yeast expression system will be chosen to develop vaccines based on mutants of the virus S glycoproteins, as it is an industrial-scale system that can produce the candidate vaccines.
In this study,the RBD glycoprotein of SARS-CoV-2 was expressed in glycoengineered yeast,and the administration of the RBD assisted by the aluminum hydroxide(Al(OH)3) and CpG adjuvants induced the production of virus-neutralizing antibody(NAb)in mice.
Adjuvants are parts of vaccines that are used to enhance the magnitude, breadth, and durability of the immune response. At present, aluminum adjuvants are the most widely used adjuvants.Their wide application benefits from their effectiveness in a variety of vaccines, better safety (with only slight side effects), and lower price. Two types of aluminum adjuvants are commonly used in licensed vaccines: aluminum hydroxide adjuvant (AH) and aluminum phosphate adjuvant (AP) [13]. Although aluminum adjuvants have been used for many years, their immuneenhancement mechanism has not been fully explained. At present, the most recognized immune-enhancement mechanism of aluminum adjuvants is the‘‘depot effect”that involves the slow release of antigens from the site of immunization. Aluminum adjuvants tend to stimulate a T helper cell 2 (Th2) immune response. CpG oligodeoxynucleotides (ODNs) are a new type of vaccine adjuvant. They are ligands for Toll-like receptor 9 (TLR9),and can induce a T helper cell 1 (Th1) immune response. Three major classes of stimulatory CpG ODNs have been identified based on their structural characteristics and their activity on human peripheral blood mononuclear cells (PBMCs)—particularly on B cells and plasmacytoid dendritic cells (pDCs) [14]. CpG1018 is a CpG-B class oligonucleotide adjuvant that is currently being evaluated in clinical trials as a potential vaccine adjuvant for coronavirus disease 2019 (COVID-19) vaccines. The CpG-based hepatitis B virus (HBV) vaccine Heplisav-B, which is an improved HBV vaccine licensed for use in adults(age>18 years),offers a simplified two-dose regimen compared with other HBV vaccines[15].
2. Material and methods
2.1. Yeast strain, bacterial strain, plasmids, and materials
The glycoengineered Pichia pastoris was reconstructed according to a previously reported method [16], which has also been described in our previous work [11]. The bacterial strain was cultured in Luria–Bertani (LB) medium containing 1.5% agar. The Escherichia coli DH5α (TakaRa Biotech, China) used for cloning was cultured at 37°C. Yeast extract, agar, and tryptone were purchased from OXOID (UK); a yeast nitrogen base without amino acids was obtained from Difco (USA); fetal bovine serum (FBS)was obtained from Beijing Qianzhao Xinye Biology Science and Technology Co., Ltd. (China); and T4 DNA ligase, restriction endonuclease, DNA ladder, Q5 DNA polymerase, and a molecular weight protein marker were obtained from New England Biolabs(USA). Anti-SARS-CoV RBD antibody and horseradish peroxidase(HRP)-goat anti-rabbit immunoglobulin G (IgG) antibody were obtained from Sino Biological (China). The DNA extraction kit and yeast genomic DNA kit were purchased from Sigma (USA).
2.2. Cloning and expression of the SARS-CoV-2 RBD
The SARS-CoV-2 RBD gene (GenBank accession number MN908947.3) was codon-optimized and synthesized by Tsingke(China). The RBD gene was cloned into the XhoI and NotI sites in the pPICZαA vector(Invitrogen,USA)to yield pPICZαA-RBD,which is driven by the AOX I promoter. The plasmid pPICZαA-RBD linearized with BglII was transformed into glycoengineered yeast by means of electroporation, and the resulting yeast was designated as Glycoeng-yeast/RBD. The Glycoeng-yeast/RBD clones were screened by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot using the anti-SARS-CoV RBD antibody and the HRP goat anti-rabbit IgG antibody(dilution ratio of 1:2500).
2.3. Yeast fermentation and the RBD purification procedure
Fermentations in 50 L bioreactors were conducted according to the methods described earlier [17]. In brief, the seed culture contained 200 mL of seed culture in yeast extract–peptone–dextrose(YPD) media. The fermentation culture contained 10 g yeast extract, 20 g peptone, 40 g glycerol, 4×10–3g biotin, 4.47 g Na2HPO4, 8.22 g NaH2PO4, and 13.4 g yeast nitrogen base (YNB)per liter of culture medium. The fermentations were carried out using the following conditions: The pH was controlled to 6.4 with NH4OH and the temperature was set at 25°C. After a 10 min starvation phase, the methanol feed (100% methanol with 12 mL∙L–1Pichia trace minerals (PTM1) salts) was initiated.Methanol induction was continued for 45–50 h.
The fermentation supernatant was recovered by centrifugation at 8000 revolutions per minute(rpm)for 25 min and then collected for RBD purification.The samples were sequentially subjected to a multimodal weak cation exchange (Capto MMC; GE Healthcare,USA), hydrophobic chromatography (Phenyl Sepharose High Performance; GE Healthcare, USA), strong anion exchange (Source 30Q; GE Healthcare, USA), strong cation exchange (Source 30S;GE Healthcare, USA), and size-exclusion chromatography (SEC;Superdex-200; GE Healthcare, USA) to finally obtain high-purity RBD protein.
2.4. Characterization of SARS-CoV-2 RBD
The purified recombinant RBD was analyzed using 12% SDSPAGE and treated with peptide-N-asparagine amidase (PNGase) F at 37°C. The purity of the RBD was analyzed by means of SEC–high-performance liquid chromatography (SEC–HPLC) and reverse-phase-high-performance liquid chromatography (RPHPLC). Absorbance values were recorded at 280 nm (Agilent 1260 HPLC).The recombinant RBD was analyzed using C8 RP-HPLC with a gradient of 0–100% mobile phase B over 40 min. Mobile phase A was composed of 0.1% formic acid (FA), and mobile phase B was composed of 95% acetonitrile and 0.1% FA.
The binding capacity of the RBD produced from yeast cells and that produced from mammalian cells to human ACE2 was measured via enzyme-linked immunosorbent assay (ELISA). The solution of ACE2 was placed in a flat-bottomed 96-well plate at a final concentration of 10 μg∙mL-1, coated with buffer(50 mmol∙L–1carbonate, pH 9.6), and left at 4°C overnight. The plate was washed four times with a blocking solution of phosphate-buffered saline containing 0.1% Tween-20 (poly(butylene succinate-co-butylene terephthalate) (PBST)). Next, blocking solution in PBST containing 5% skim milk was added to the plate,which was then incubated for 1 h at 37°C. Serially diluted recombinant RBD was added,incubated at 37°C for 1 h,and then washed four times. The anti-SARS-CoV-2 RBD antibody was diluted by 1:2000 and added to the wells. After being incubated at 37°C for 1 h, the plates were washed four times. The goat anti-rabbit IgG horseradish peroxidase (HRP) binding antibody was diluted 1:2000 and added to the wells. After being incubated at 37°C for 1 h, the plates were washed four times and then developed with 3,3′,5,5′-tetramethylbenzidine for 15 min. 2 mol∙L–1H2SO4stop solution at 50 μL per well was added to stop the reaction.
2.5. Glycoform analysis experiment
The DNA sequencer-assisted–fluorophore-assisted carbohydrate electrophoresis (DSA–FACE) method described previously[15]was used to detect oligosaccharide chains on the RBD.In brief,the glycoprotein was treated by glucosidase, and the sugar chains were marked with the fluorescent marker 8-aminopyrene-1,3,6-trisulfonic acid (APTS). The samples were separated under an ABI 3100 DNA sequencer (USA).
2.6. Mice and vaccinations
BALB/c mice were obtained from Beijing Weitong Lihua Laboratory Animal Technology Co.,Ltd.and raised at the Animal Center of the Beijing Institute of Biotechnology.All animal experiments were approved by the Institutional Animal Care and Use Committee of the Beijing Institute of Biotechnology and conducted in accordance with the institutional guidelines of the committee.The experimental animal welfare ethics number is IACUC-DWZX-2020-039.
Female BALB/c mice aged 6–8 weeks were randomly divided into three groups(n=10)as the first batch of animal experiments:the 10 μg RBD/100 μg aluminum hydroxide(CRODA,Denmark)gel group; the 10 μg RBD/100 μg aluminum hydroxide gel/25 μg CpG group; and the phosphate-buffered saline (PBS) control group.Mice were immunized with 100 μL (intramuscular injection) in their hind leg muscles on days 0, 14, and 28.
To determine the optimal immune dose of the RBD, 6–8 weekold BALB/c female mice were randomly divided into four groups(n=5) as the second batch of animal experiments: the 10 μg RBD/50 μg aluminum hydroxide gel/25 μg CpG group; the 5 μg RBD/50 μg aluminum hydroxide gel/25 μg CpG group; the 2.5 μg RBD/50 μg aluminum hydroxide gel/25 μg CpG group; and the PBS control group. Mice were immunized with 100 μL (intramuscular injection) in their hind leg muscles on days 0 and 14. To determine the best immunization strategy, we immunized the BALB/c mice at different times (n=5) as the third batch of animal experiments: two immunizations at days 0 and 14, respectively;two immunizations at days 0 and 21,respectively;two immunizations at days 0 and 28, respectively; and three immunizations at days 0, 14, and 28, respectively. The mice were immunized with 100 μL of 10 μg RBD/50 μg aluminum hydroxide gel/25 μg CpG(intramuscular injection) in their hind leg muscles.
2.7. Histology and immunohistochemistry
The five animals of each group in the first batch of animal experiments were decapitated four weeks after the last vaccine injection. The organs, including the hearts, lungs, livers, and spleens, were collected and fixed with formaldehyde, embedded in paraffin, and cut into slices. These sections were stained with hematoxylin and eosin,observed with a light microscope,and analyzed for histology.
2.8. Analysis of RBD-specific serum antibodies
The five animals of each group in the first batch of animal experiments were bled on days 14,28,42,98,154,and 182(weeks 2,4,6,14,22,and 26).The animals of the second and third batches of animal experiments were bled two weeks after the first and last vaccination. After the blood had clotted for 1–2 h at room temperature, the samples were centrifuged at 5000 rpm at 4°C for 15 min. The upper serum was collected and stored at –20°C.The recombinant RBD was coated onto a flat-bottom 96-well plate at a final concentration of 2 μg∙mL-1in coating buffer(50 mmol∙L–1carbonate, pH 9.6), and then left at 4°C overnight. The plate was washed four times with PBST. Next, blocking solution in PBST containing 5% skim milk was added to the plate, which was then incubated for 1 h at 37°C. Serially diluted mouse serum was then added. The plate was incubated at 37°C for 1 h, and then washed four times. The goat anti-mouse IgG HRP binding antibody was diluted to 1:4000 and added to the wells. After being incubated at 37°C for 1 h, the plates were washed four times and developed with 3,3′,5,5′-tetramethylbenzidine for 15 min. 2 mol∙L–1H2SO4stop solution at 50 μL per well was then added to stop the reaction.The absorbance was detected on a 450 nm microplate reader.
2.9. SARS-CoV-2 neutralization assay
A micro-neutralization assay was used to detect the neutralization activities of the immunized mice serum in the biosafety level 3(BSL-3) facility [16]. In brief, serially diluted sera were incubated with 100 50% tissue culture-infectious dose (TCID50) of the SARS-CoV-2 strain (IME-BJ01) at 37°C for 2 h. Thereafter, the virus-serum complexes were transferred to a monolayer of Vero E6 cells cultured in 96-well plates.The inoculated plates were then incubated for 72 h and stained with 0.05%crystal violet for 35 min.Optical density(OD)values were measured at 570 or 630 nm after the addition of the decolorization solution. The neutralization results were calculated using the Reed-Muench method in order to estimate the serum dilution (50% effective concentration(EC50)titer)required to obtain half of the maximum neutralization of the infection. Seropositivity was defined as a titer ≥16. The initial serum dilution (1:16) was set as the confidence limit of the assay.
Neutralizing activities against the SARS-CoV-2 501Y.V2 and B.1.1.7 pseudoviruses in the immunized mice serum were tested.The EC50was defined as the serum dilution at which the relative light units (RLUs) were reduced by 50% compared with the virus control wells(virus+cells)after the subtraction of the background RLUs in the control groups with cells only.In brief,the pseudovirus was incubated with serial dilutions of the test samples (six dilutions in a three-fold step-wise manner) in duplicate for 1 h at 37°C, together with the virus control and cell control wells in sexaplicate.Freshly trypsinized cells were then added to each well.After 24 h of incubation in a 5%carbon dioxide(CO2)environment at 37°C, the luminescence was measured, as described in section on pseudovirus titration. The EC50values were calculated with nonlinear regression—that is, log (inhibitor) versus response (four parameters)—using GraphPad Prism 8 (GraphPad Software, Inc.,USA).
3. Results
3.1. Expression of SARS-CoV-2 RBD protein in glycoengineered Pichia pastoris
The RBD of the S protein was prepared using the glycoengineered Pichia pastoris expression system, which has mammalian N-glycosylation modification capacity[12].The RBD gene sequence of SARS-CoV-2 encodes a glycoprotein of 223 amino acids(residues 319–541 of the S protein) with a theoretical molecular mass of 25 kilodalton (kDa) and two N-glycosylation sites (N331 and N343). A schematic of the S protein containing the RBD gene and the expression vector for the RBD gene are shown in Figs. 1(a)and (b), respectively. The RBD gene was cloned between the XhoI and NotI sites of the pPICZαA vector to generate pPICZαARBD (SARS-CoV-2) (Fig. 1(b)). The linearized pPICZαA-RBD was transformed into glycoengineered yeast, and the resultant yeast strain was designated as Glycoeng-yeast/RBD. The results from Western blot testing revealed that all of the examined Glycoengyeast/RBD clones had approximately 30 kDa bands (Fig. 1(c)). The observed band from the SDS-PAGE analysis was heterogeneous,but the observed molecular weight (30 kDa) was higher than the theoretical molecular mass of 25 kDa, which may be because of the glycosylation of RBD.
3.2.Efficient purification of RBD from glycoengineered Pichia pastoris
We established a 4 L scale yeast fermentation process,with the whole fermentation cycle lasting for five days (Fig. 2(a)). Because RBD carries an α-factor secretion signal peptide (Fig. 1(b)), the expressed RBD was secreted into the culture supernatant. SDSPAGE analysis of the supernatant from yeast induced at different time points showed that the expression level of the RBD was high after 48 h of induction(Fig.2(b)).The RBD secreted into the supernatant was obtained by means of a cation exchange column,hydrophobic chromatography,anion chromatography,cation chromatography, and gel filtration chromatography. High-purity RBD protein was obtained with a yield of 50 mg∙L–1.SDS-PAGE analysis confirmed that the purified RBD protein ran as a single band(Fig.2(c)). The RBD protein showed single, sharp SEC–HPLC (TSK gel G2000SWXL, 5 μm, φ 7.8 mm×300 mm) and RP–HPLC (Agilent ZORBAX 300SB-C8,5 μm,φ 4.6 mm×250 mm)profiles,indicating high purity and integrity (Fig. 2(d)). The binding capacity of the RBD produced from yeast cells and mammalian cells to human ACE2 was measured by ELISA. The absorbance was detected on a microplate reader of 450 nm. The results indicated no significant difference between the binding capacity of the RBD produced from yeast cells and that produced from mammalian cells to human ACE2 (P=0.5784) (Fig. 2(e)).
3.3. Purified RBD shows complex glycosylated modifications
The apparent molecular mass of the purified RBD protein(approximately 30 kDa)was about 25%greater than the theoretical molecular mass (approximately 25 kDa). The RBD protein treated with PNGase F was consistent with theoretical mass of 25 kDa (Fig. 3(a)), suggesting that the RBD had undergone N-glycosylation modification. The RBD expressed by glycoengineered yeast had a terminally biantennary galactosylated glycan,as analyzed by DSA–FACE (Fig. 3(b)). DSA–FACE was used to characterize the RBD N-glycans by using bovine ribonuclease B as a marker,and five kinds of glycans(Man5GlcNAc2to Man9GlcNAc2,or M5–M9) were identified (Fig. 3(b-I)) [18]. The glycans of the RBD expressed in yeast were incubated with β1-4-galactosidase, β-N-acetylglucosaminidase, and α1-2,3 mannosidase,respectively,or with a mixture of β1-4-galactosidase and β-N-acetylglucosaminidase. The β1-4-galactosidase removed the terminal galactoses on the N-glycan from Gal2GlcNAc2Man3-GlcNAc2, resulting in the generation of GlcNAc2Man3GlcNAc2(Fig. 3(b-IV)). The β-N-acetylglucosaminidase further cut the N-acetylglucosamine from the N-glycans to generate Man3GlcNAc2(Fig. 3(b-V). The α1-2,3 mannosidase was used to generate Man2GlcNAc2from Man5GlcNAc2(Fig. 3(b-VI)). Over all, the yeast-expressed RBD was modified at the afucosylated mannosetype and complex glycans characterized as Man5GlcNAc2and Gal2-GlcNAc2Man3GlcNAc2(Fig. 3(b-II)). In comparison, previous work on HEK293-derived and Chinese hamsters ovary cells-derived RBD has shown that they mainly possess fucosylated diantennary and triantennary complex-type N-glycans and have a small amount of high mannose N-glycans (Man5, Man6) with or without phosphorylation [19].
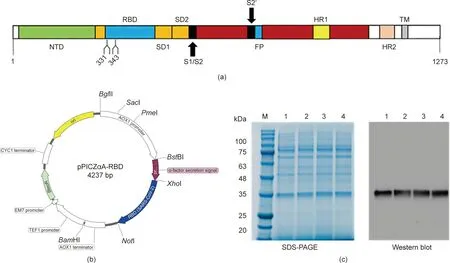
Fig.1. Characterization and expression of the RBD protein of SARS-CoV-2.(a)Schematic of the SARS-CoV-2 S protein sequence,showing the N-terminal domain(NTD),RBD,heptad repeats 1 and 2 (HR1 and HR2), transmembrane domain (TM), subdomains 1 and 2 (SD1 and SD2), fusion peptide (FP), and transmembrane region (TM). Nglycosylation sites are marked with the residue numbers(331 and 343).(b)Map of the pPICZαA-RBD plasmid.(c)SDS-PAGE and Western blot analysis of various Glycoengyeast/RBD clones, shown in lanes 1, 2, 3, and 4. M: marker; CYC1: cytochrome C1; EM7: synthetic prokaryotic promoter that drives constitutive expression of the Zeocin resistance gene in Escherichia coli; TEF1: translation elongation factor 1; AOX1: aldehyde oxidase 1; BleoR: bleomyicin-R.
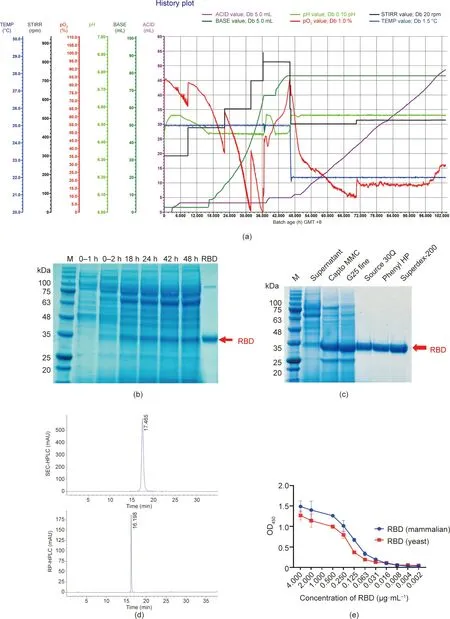
Fig.2. Fermentation of yeast35/RBD and the RBD purification procedure.(a)The complete fermentation process of the RBD protein,showing the stirring speed(STIRR;black),dissolved oxygen(pO2;red),variations of temperature(TEMP;blue),base filling amount(BASE;dark green),pH(light green),and acid filling amount(ACID;purple).(b)SDSPAGE analysis of RBD expression at different culture times. (c) SDS-PAGE of the RBD during the purification process. (d) SEC–HPLC and RP–HPLC of the purified RBD.(e) Binding capacity to human ACE2 of RBD produced from yeast cells and RBD produced from mammalian cells. Phenyl HP: phenyl sepharose high performance;G25 fine:Sephadex G25 coarse; Superdex-200: Superdex-200 prep grade; OD450: optical density at 450 nm; AU: absorbance unit.
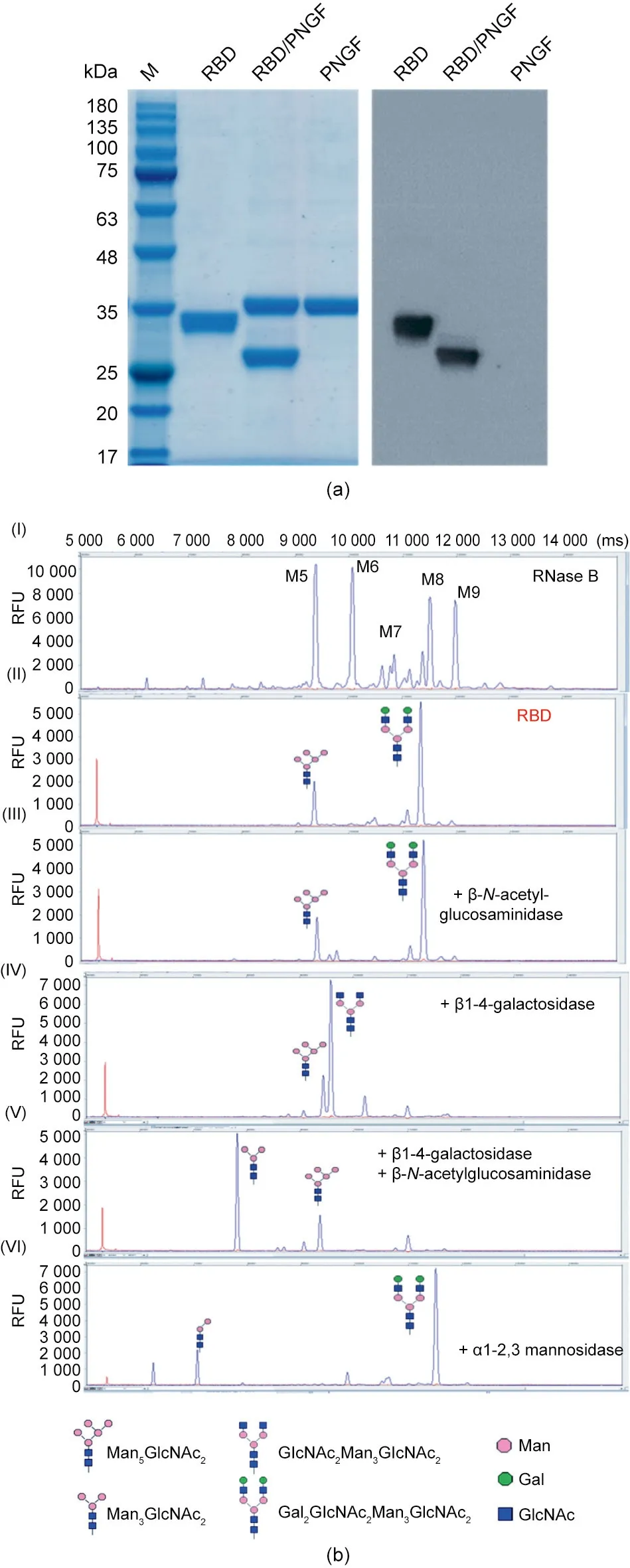
Fig. 3. Characterization of the RBD. (a) SDS-PAGE results of RBD digested with PNGase F.(b)N-glycans analysis of the RBD protein:(I)Reference glycans of bovine RNase B (Man5GlcNAc2–Man9GlcNAc2); (II) N-glycans of RBD protein; (III) glycans of RBD digested with β-N-acetylglucosaminidase;(IV)glycans of RBD digested with β1-4-galactosidase; (V) glycans of RBD digested with β-N-acetylglucosaminidase and β1-4-galactosidase; (VI) glycans of RBD digested with α1-2,3 mannosidase.PNGF: peptide-N-glycosidase F; RFU: relative fluorescence unit.
3.4. Identification of antibodies against RBD
To develop vaccines using the RBD, we selected aluminum hydroxide as the adjuvant for its safety and effectiveness. To prepare the vaccine, aluminum hydroxide was added to the RBD,resulting in precipitation of the candidate vaccine with aluminum gel.To evaluate the potential of the RBD as a vaccine,the RBD was given to BALB/c mice with CpG (CpG2006) [13,20] and aluminum hydroxide as double adjuvants. PBS was given as the control. At 14 days after the last immunization, serum samples were collected.
The RBD immunogen with the combination of CpG and aluminum hydroxide as the adjuvants markedly increased the immunogenicity of the antibody titer (Fig. 4(a)). At 14 days after the first immunization, the specific antibody titers of the RBD+Al(OH)3+CpG group and the RBD+Al(OH)3group were 7.4×104and 66.0, respectively (P<0.0001). At 14 days after the second immunization,the antibody titer of the RBD+Al(OH)3+CpG group was 6.8×106and that of the RBD+Al(OH)3group was 7.9×103. The RBD+Al(OH)3+CpG group induced a significantly higher antigen-specific IgG compared with the RBD+Al(OH)3group(P<0.0001). Because of the low antibody titer in the RBD+Al(OH)3group, a third immunization was performed. Two weeks later, the antibody titer of the RBD+Al(OH)3group was 3.4×105,but that of the RBD+Al(OH)3+CpG group(i.e.,four weeks after the second immunization) without the third immunization was approximately 4.5×106(P<0.0001). These results indicated that the RBD with both CpG and aluminum hydroxide was superior to the RBD with the aluminum hydroxide adjuvant as an immunogen.
We next tested sera from the groups for neutralizing activities against live SARS-CoV-2 virus (IME-BJ01) (Fig. 4(b)). At 14 days after the first immunization, the titers of NAb in the RBD+Al(OH)3+CpG group and the RBD+Al(OH)3group were below 1:16, which was basically the same as that in the negative control group. At 14 days after the second immunization, the NAb titer of the RBD+Al(OH)3group was still below 1:16, but that of the RBD+Al(OH)3+CpG group was 1:1000 (ranging from 430 to 2800). The RBD+Al(OH)3+CpG group elicited higher titers of NAb compared with the RBD+Al(OH)3group(P<0.05).At 14 days after the third immunization, the NAb titer was 1:105 (ranging from 33 to 210) in the RBD+Al(OH)3group; the NAb titer in the RBD+Al(OH)3+CpG group without the third immunization (four weeks after the second immunization) was 1:1259 (ranging from 310 to 1900) (P<0.05).
To determine the optimal immunity dose of RBD,we performed the experiment with different doses of the RBD antigen.At 14 days after the first immunization, the antibody titers of the 10 μg RBD group and the 5 μg RBD group were 7.9×104and 5.0×104(P=0.1268) (Fig. 4(c)). The antibody titer of the 2.5 μg RBD group was 5.0×104, which was not significantly different from that of the 10 μg RBD group (P=0.1268). At 14 days after the second immunization, the antibody titers of the 10 μg RBD group and the 5 μg RBD group were 3.2×106and 2.5×106, respectively(P=0.3739). The antibody titer of the 2.5 μg RBD group was 1.0×106, and less antigen-specific IgG was induced compared with the 10 μg RBD group(P<0.05).These results indicated no significant difference in the antibody titer induced by 10 μg or 5 μg of RBD antigen immunization to BALB/c mice with 50 μg Al(OH)3and 25 μg CpG per dose; however, the immunity effect using 2.5 μg RBD was poor.
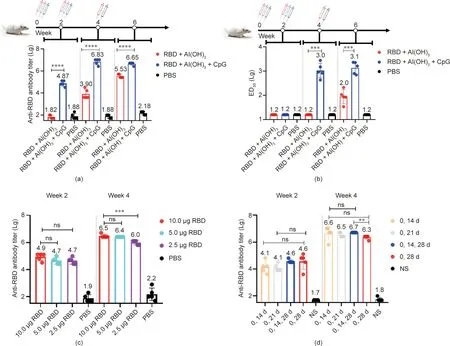
Fig.4. Immunity effects of vaccines with different adjuvant components on BALB/c mice.(a)Antibody titers after three immunizations with vaccines with different adjuvant components; no third immunization was performed in the RBD+Al(OH)3+CpG group. (b) Antibody neutralizing activity of vaccines with different adjuvant components after three immunizations;no third immunization was performed in the RBD+Al(OH)3+CpG group.(c)Antibody titers of 100 μL vaccine preparations mixed with different doses of RBD (10.0, 5.0, and 2.5 μg), 50 μg of Al(OH)3 adjuvant and 25 μg of CpG for 14 days and 28 days after the first immunization. (d) Antibody titers at 14 days after immunization with the RBD vaccine with different immunization strategies(5 μg RBD mixed with 50 μg Al(OH)3 adjuvant and 25 μg CpG mixed in 100 μL).ns:no significant difference; NS: normal saline. P-values were analyzed with t tests; ns: p > 0.05, **: p < 0.01, ***: p < 0.001, ****: p < 0.0001.
To determine the best immunization strategy, we immunized BALB/c mice at different time: two immunizations at days 0 and 14; two immunizations at days 0 and 21; two immunizations at days 0 and 28; and three immunizations at days 0, 14, and 28. At 14 days after the first immunization, no significant difference was found in the antibody titer (Fig. 4(d)). At day 14 after the second immunization, the antibody titers were 4.0×106in the‘‘0,14 d”group and 3.2×106in the‘‘0,21 d”group.The titers were 5.0×106in the ‘‘0, 14, 28 d” group at 14 days after the third immunization and 2.0×106in the‘‘0,28 d”group at 14 days after the second immunization. The ‘‘0, 14, 28 d” group induced significantly more antigen-specific IgG compared with the ‘‘0,28 d”group.No significant difference was noted between the other groups.These results indicate that the‘‘0,14 d,”‘‘0,21 d,”or‘‘0,14,28 d” schedules could be potential immunization strategies.
3.5. Safety evaluation
To evaluate the toxicity of the RBD candidate vaccine,histopathological changes of the main organs including the liver,lung, and heart tissues were examined by light microscopy. The results revealed no obvious differences between the experimental and control groups (Fig. 5).
3.6. Characterization of the persistence of RBD antibody response
To determine the persistence of the antibody titer,blood samples were collected from the tails of mice at weeks 2,4,6,14,22,and 26 after immunization. The antibody titer of the RBD+Al(OH)3+CpG group was 6.3×106at 14 days and 3.2×106at 12 weeks after the second immunization (Fig. 6(a)) (P=0.0604).The antibody titer was 6.3×105at 22 weeks after immunization and 6.3×105after 26 weeks. The antibody titer of the RBD+Al(OH)3group was 3.2×105at two weeks after the third immunization and 6.3×105at 14 weeks after the immunization(P=0.1823).The antibody titer was 1.6×105at 22 weeks after the immunization,and there was no significant difference with the antibody titer 2 weeks after the third immunization (P=0.1878). At 26 weeks after immunization,the antibody titer was 2.5×104.
Fig.5. Possible histopathological changes to organs including liver,lung,and heart tissues(stained by hematoxylin and eosin)were observed by means of light microscopy.
To determine the persistence of the NAb titer, blood was collected at weeks 2, 4, 6, 14, 22, and 26 after the last immunization.The NAb titer of the RBD+Al(OH)3+CpG group was 1:1000(ranging from 430 to 2800)at four weeks after immunization and 1:501(ranging from 250 to 1500) at 26 weeks after immunization(P=0.2218) (Fig. 6(b)). The antibody titer of the RBD+Al(OH)3group was 1:105 (ranging from 33 to 210) at two weeks after the third immunization and 1:63 (ranging from 28 to 170) after 26 weeks (P=0.5545).
At 14 days after the third immunization, the NAb titer of the RBD+Al(OH)3group against 501Y.V2 was 1:182, and that of the RBD+Al(OH)3+CpG group was 1:347. At 14 days after the third immunization, the NAb titer of the RBD+Al(OH)3group against B.1.1.7 was 1:832, and that of the RBD+Al(OH)3+CpG group was 1:1047. Thus, our data show that the vaccination determined cross-neutralization against B.1.1.7 and 501Y.V2 (Fig. 6(c)).
4. Discussion
Similar to the S proteins of other viruses,the S protein of SARSCoV-2 is coated with carbohydrate chains,which potentially mask epitopes to block the host’s immune response.The S protein monomer has 22 N-glycosylation sites[21],with two glycosylation sites,N331 and N343, in the RBD region. Correct glycosylation of antigens is critical for immunogenicity. Many vaccine technologies based on the S protein currently use mammalian cells or insect cells as the expression system in order to ensure the correct glycosylation modification. Yeast, a lower eukaryote, has the ability of hyper-mannosylation. Studies have shown that hypermannosylation modifications may mask the epitope of antigens[22]. The H7N9 influenza virus hemagglutinin (HA) subunit particle vaccine prepared by means of glycosylation-modified humanized yeast, which has mammalian N-glycosylation ability,has been shown to have good immunogenicity and protected mice from the H7N9 virus [11]. In this study, we used glycoengineered Pichia pastoris to express SARS-CoV-2 RBD. DSA–FACE analysis showed that the N-glycosyl groups in the RBD were a fucose-free complex glycosyl group and a low-mannose glycosyl group,which ensured that the RBD epitope could not be masked.
To examine the safety of the candidate RBD vaccine, we examined the potential toxicity of the RBD vaccine in mice. No adverse effects were observed,such as body weight and behavioral changes(data not shown). There were no significant pathological changes in the liver, lung, heart, and other organs on microscopic examination.
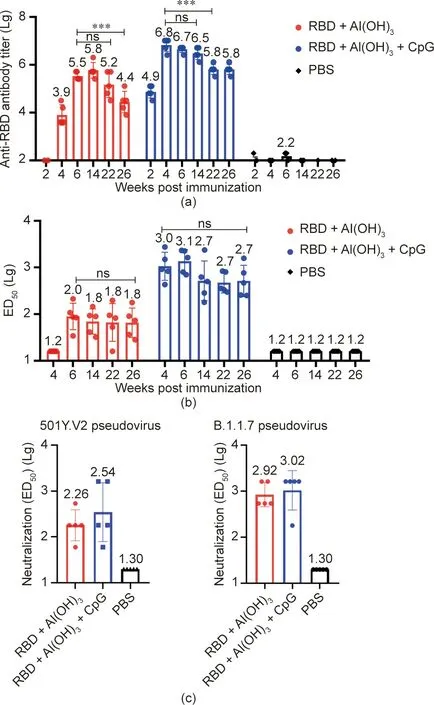
Fig. 6. Detection of RBD antibody persistence at 6 months after immunization.(a) Antibody titer persistence at 26 weeks after immunization with different adjuvant components. (b) Antibody neutralizing activity persistence at 26 weeks after immunization with different adjuvant components. (c) Neutralizing activities against the SARS-CoV-2 501Y.V2 and B.1.1.7 pseudoviruses. ED50:median effective dose. P-values were analyzed with t tests; ns: p > 0.05, ***: p < 0.001.
In our study, we found that Al(OH)3combined with CpG had a significant adjuvant effect. In mice, Al(OH)3combined with CpG induced a strong humoral and cellular immune response against the RBD. During the 26-week follow-up, the titer of virus-NAb did not significantly decrease. These results provide immunogenicity research data for a vaccine using RBD expressed by glycoengineered yeast as the immunogen.Importantly,the glycoengineered yeast production platform can quickly respond to virus mutations with an expandable production scale.Thus,the preparation of candidate vaccines can be expected to be completed with a stable production process within a few weeks. The candidate RBD vaccine presented here has the potential to contribute significantly to the control of the COVID-19 pandemic.If the SARS-CoV-2 subunit vaccine with the CpG and Al(OH)3adjuvants proves to be successful,rapid production of viral glycoproteins based on glycoengineered yeast will become an important new technology for rapid response to any emerging viral threat.
Acknowledgments
This work was supported by the National Key Research and Development Program of China (2020YFC0841400-008), the NationalScienceandTechnologyMajorProjects(2018ZX09711003-013-002), and the National Natural Science Foundation of China (81673339 and 81773619).
Compliance with ethics guidelines
Bo Liu, Ying Yin, Yuxiao Liu, Tiantian Wang, Peng Sun, Yangqin Ou, Xin Gong, Xuchen Hou, Jun Zhang, Hongguang Ren, Shiqiang Luo, Qian Ke, Yongming Yao, Junjie Xu, and Jun Wu declare that they have no conflict of interest or financial conflicts to disclose.

- Engineering的其它文章
- Electric Air Taxis Create Megadeal Buzz
- Tissue Engineering and Regulatory Science
- Factors Predicting Progression to Severe COVID-19: A Competing Risk Survival Analysis of 1753 Patients in Community Isolation in Wuhan,China
- Fabrication and Applications of Multi-Fluidic Electrospinning Multi-Structure Hollow and Core–Shell Nanofibers
- Past and Future Changes in Climate and Water Resources in the Lancang–Mekong River Basin: Current Understanding and Future Research Directions
- Mass Transfer,Gas Holdup,and Kinetic Models of Batch and Continuous Fermentation in a Novel Rectangular Dynamic Membrane Airlift Bioreactor