
Wheat genotypes tolerant to heat at seedling stage tend to be also tolerant at adult stage: The possibility of early selection for heat tolerance breeding
2022-08-16 09:25:26LuLuHuiLiuYuWuGuijunYan
The Crop Journal 2022年4期
Lu Lu, Hui Liu, Yu Wu, Guijun Yan
a UWA School of Agriculture and Environment, The University of Western Australia, Perth, WA 6009, Australia
b The UWA Institute of Agriculture, The University of Western Australia, Perth, WA 6009, Australia
c Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, Sichuan, China
Keywords:Heat tolerance Seedling stage Adult stage Linear regression Genetic mechanism
A B S T R A C T Wheat production is seriously influenced by extreme hot weather,which has attracted increasing attention. It is important to compare wheat responses to heat at seedling and reproductive stages, to explore the potential relationship between the performances at different growing stages and the possibility of early selection to accelerate heat tolerance breeding.In this study,forty wheat genotypes were screened under heat stress at both seedling and adult stages.It was found that root lengths at seedling stage were severely reduced by heat stress with significant variations among wheat genotypes.Heat-tolerant genotypes at seedling stage showed less root length decrease than susceptible ones.Wheat genotypes tolerant at seedling stage showed higher yield at adult stage after heat treatment. The performances of wheat genotypes screened under heat stress at seedling and adult stages were ranked by seedling damage index and adult damage index.A significant positive relationship was found between heat tolerance at seedling stage and at adult stage (r=0.6930), suggesting a similar tolerant/susceptible mechanism at different plant growth stages and the possibility of early selection at seedling stage for breeding heat tolerance.Extremely tolerant and susceptible genotypes with consistent performances at seedling and adult stages were genetically compared and associated SNP markers and linked candidate genes were identified.
1. Introduction
Wheat production is seriously influenced by extreme hot weather [1-7]. Heat stress can affect the crop at different stages,including seedling stage [8] and reproductive stage [9-11]. Under the changing climate, it is important to accelerate breeding of wheat cultivars with more tolerance to high temperature stress.
A challenge for study of heat tolerance in wheat is the identification of reliable selection criteria and efficient screening methods to detect heat-tolerant plants [12]. Due to the difficulty of screening wheat genotypes under heat stress in various spatial environments, no consistent selection criterion for heat tolerance has been established. Instead, some indirect selection criteria were suggested.
For seedling stage, phenotypic screening can be done for traits such as plant fresh weight,dry weight,carbon isotope discrimination, canopy temperature and spectral reflectance indices [13] or chlorophyll content,chlorophyll fluorescence parameters and their heat tolerance index [14] for selection of tolerant genotypes. Root was reported as a reliable predictor of plant responses to drought[15] as it contacts directly with soil or hydro environment for water and nutrition absorption under stress. Our preliminary observation(unpublished data)at seedling stage screening showed that 1) root traits, especially root length,were more influenced by heat stress than shoot traits;and 2)among root traits,fresh weight and dry weight generally showed relatively small variations among different genotypes at seedling stage.
There are many options for phenotypic screening of heat tolerance at adult stage such as canopy temperature, leaf senescence,night respiration, chlorophyll fluorescence and cell membrane thermo-stability [16,17], especially the traits already been proved having strong correlations with grain yield in wheat, such as loss of chlorophyll content during grain filling, thylakoid membrane damage and plasma membrane damage [18]. SPAD (Soil Plant Analysis Development) meter is a simple, quick and non-destructive method to measure chlorophyll content, with advantages for rapid screening.However,final yield is still the best measurement for heat tolerance at adult stage. The post-anthesis reproduction and grain-filling stages are the most heat sensitive periods where yield is the most impacted by heat stress in wheat[9-11], so it was often selected to apply the stress treatment,and the tolerance during this stage is extensively studied[10,18-28].
It has been suggested that early vegetative stage such as seedling stage of wheat have greater stress avoidance and resilience[29], and seedling establishment may have large impacts on crop yield and stand [30]. Moreover, it is easy and convenient for mass phenotyping and high-throughput characterisation during seedling stage,so early generation screening and selection has attracted considerable interests [12,31-34]. Some reports indicated that some particular quantitative trait loci (QTL) at seedling stage and reproductive stage were collocated on the same chromosomes[14],moreover,some well-known heat-induced genes at reproductive stage were found significantly enhanced in acetylation levels in wheat seedlings[35].These discoveries made it necessary to further study the relationship of heat tolerance performances at different stages of wheat.
Alsamadany [36] screened 499 wheat genotypes at seedling stage in order to investigate the genetic variation at early growth stage in wheat. Genotypes were evaluated for heat tolerance by subjecting them to heat stress(35°C)and non-stress(25°C)conditions. The results provided materials with known performances at seedling stage. On the other hand, Collins et al. [24,37] screened a wide range of wheat germplasm to identify genotypes with stable and high yield under heat stress for breeding. Various genotypes were identified in greenhouse/growth chamber experiments and in irrigated field trials in southern New South Wales(NSW)in Australia where delayed sowing was used to impose heat stress. This outcome provided a list of wheat genotypes with known performances for heat tolerance at adult stage.
Few comparison and association of different stages in wheat for heat tolerance performances have been reported. Our hypotheses are: 1) a relationship may exist for heat tolerance at early stage and adult stage; and 2) there is possibility of early selection in wheat breeding for heat tolerance due to this relationship. In this study, experiments were correspondingly designed according to these hypotheses using selected wheat materials from the two studies [36,37]. On one hand, genotypes with extreme performances at seedling stage [36] were selected by targeted selection for screening of adult performances under heat stress;on the other hand, representative genotypes with known adult performances[37] were selected to test their seedling-stage performances. The objectives of this study are to 1)examine if extreme tolerant genotypes can show similar performances at adult stage; 2) find the relationship/trend of heat-tolerant performances at different stages in wheat; and 3) identify SNP markers and linked genes associated with performances under heat. The general approach for investigation based on the developmental stages of bread wheat [38] was shown in Fig. 1.
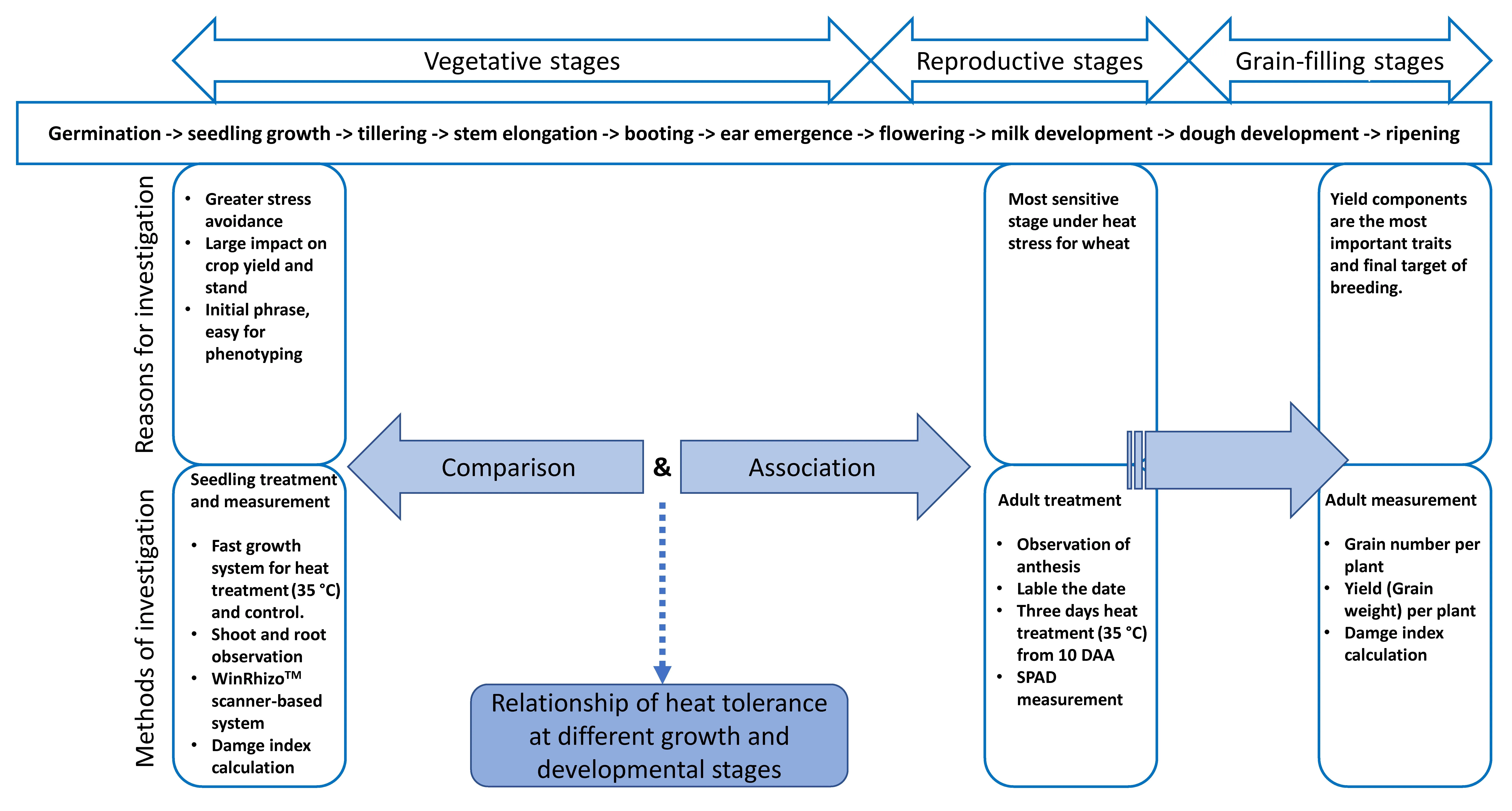
Fig.1. Flowchart showing the methodologies used for investigation at different growth stages.Seedling stage was studied for early tolerance under heat stress.Post-anthesis period (from flowering till harvest was studies for adult stage heat tolerance. By comparison and association of performances of various genotypes, relationship of heat tolerance at different stages was revealed. DAA means days after anthesis.
2. Materials and methods
2.1. Germplasm used for screening
2.1.1. Genotypes for seedling screening with known performances at adult stage
Twenty-four wheat cultivars (Table S1) were selected for seedling stage heat-tolerance(HT)screening.Their HT performances at adult stage with accessible data related to yield traits were reported [24,37]. They were selected as the representatives from 137 genotypes in greenhouse/growth chamber experiments and in irrigated field trials.
2.1.2. Genotypes for adult stage screening with known seedling stage performance
Sixteen wheat genotypes(Table S2)including eight extreme tolerant genotypes and eight extreme susceptible genotypes were selected for adult stage HT screening to investigate phenotype relationship between seedling stage and adult stage [36]. They were identified as extreme genotypes for HT at seedling stage in controlled environment room (CER) from a previous screening of 499 genotypes for their seedling heat tolerance performances [36].They represented the two tails in a normal distribution in the population of the 499 genotypes.

Table 1SNP markers and genes identified common for two stages.
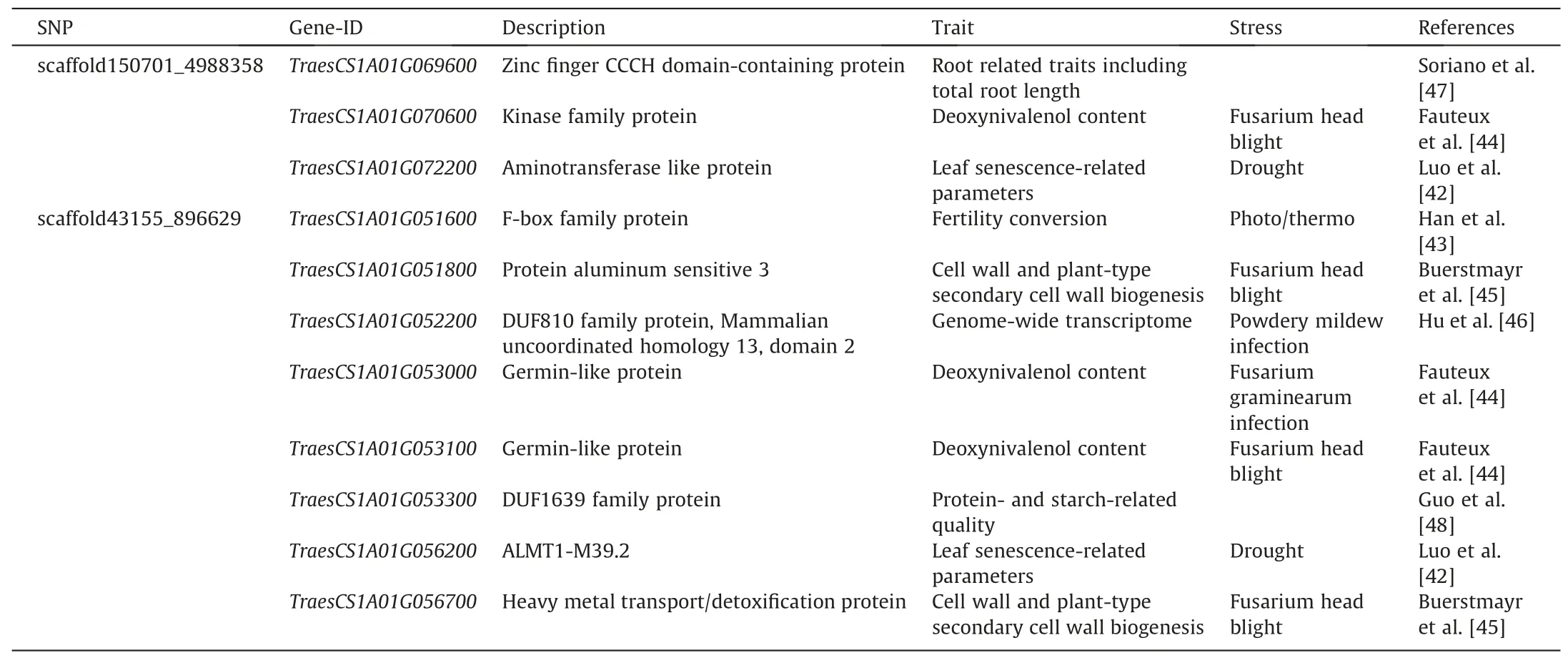
Table 2Candidate genes that have been reported previously.
2.2. Experimental set-up and treatment application
2.2.1. Seedling stage screening
A hydroponic growth system [36], which enabled fast evaluation of a large number of genotypes, was developed using a large plastic box(58.5 cm×42.0 cm×33.0 cm dimension)with parallel layers of vertically organized boards across the width of the box with wheat seeds in between (Fig. S1). 35°C was used for stress treatment [36], and the optimum temperature for wheat growth(25°C) [32,39] was used as the standard control temperature.
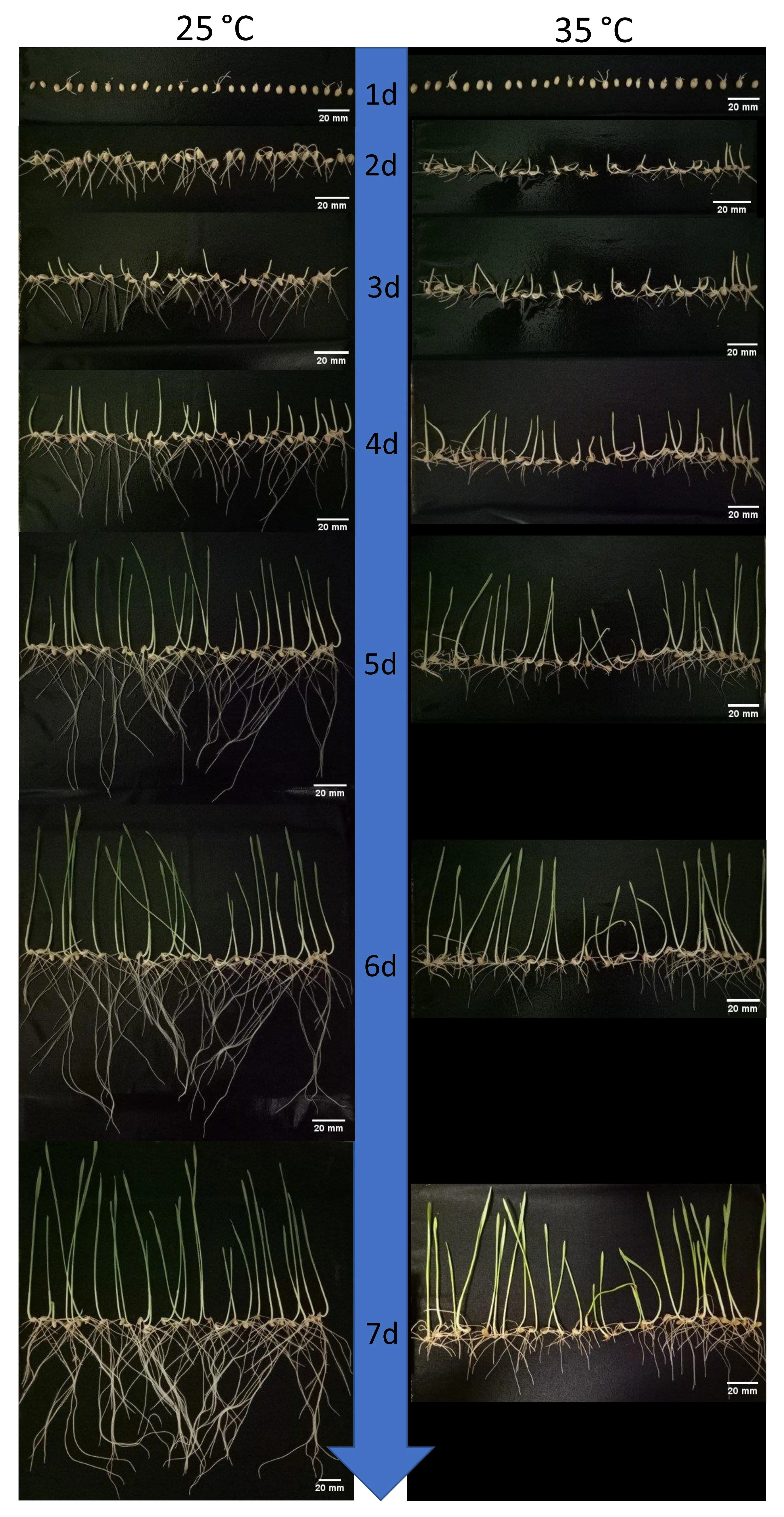
Fig. 2. Comparison of seedling-stage growth under non-stress (25°C) and heat stress (35°C) for seven days in cultivars ‘‘Ventura” etc. Compared to shoots, the roots were impacted adversely much more severely by heat stress.Roots and shoots can grow to similar length under non-stress after seven days,however root lengths reduced considerably under heat stress condition.Seed 1,2,3(from left)=Ventura;seed 4,5,6=Livingston;seed 7,8,9=Sokoll;seed 10,11,12=Espada;seed 13,14,15=Sentinel; seed 16, 17, 18=Zebu; seed 19, 20, 21=Dakota; seed 22, 23,24=Clearfield JNZ and seed 25, 26, 27=Guardian. Only 9 genotypes are shown here, similar results were found in other genotypes. 1 d=day 1 ... 7 d=day 7 as shown in the blue bar between the two photo columns.
Ten seeds for each genotype were soaked overnight in distilled water at 25°C. Six germinated seeds were randomly selected and placed in the growth holder system, pre-moistened with distilled water. Three replications were used for each environment of control and heat treatment. The experiment was conducted at two environments: one in a controlled environment room (CER) with 14-hour photoperiod (200 μmol m-2s-1), air relative humidity of 70.0%-75.0% and constant temperature of 25±1°C; and the other in similar conditions but at a temperature of 35±1°C. Distilled water matching the temperature was added to each container as needed throughout the evaluation period, to avoid water-deficit stress. The assembled units in the growth system were opened daily and photos were taken to record changes. One week later all the plants were taken out for measurements. WinRhizo scanner-based system (v.2007, Regent Instruments Inc., Sainte-Foy,Quebec,Canada)were used for measuring root traits including total root length,root surface area,root diameter and root volume.
2.2.2. Adult stage screening
The seeds of all genotypes were germinated in Petri dishes with water,before sowing one plant per pot(8 cm×8 cm×16 cm)containing soil media (5: 2: 3 compost: peat: sand, pH 6.0) [19]. For each line, six pots (three replicates each for the control and the heat-stress treatment) were grown in a naturally lit glasshouse at The University of Western Australia,Crawley,Western Australia(31°59′S, 115°49′E). The plants were fertilized fortnightly with‘Diamond Red’ (Campbells Fertilisers Australasia Pty Ltd, Laverton North,Australia)from four weeks after sowing until the end of the grain-filling period. The experiment was arranged in a completely randomized block design.
Anthesis date, as Zadoks’ growth scale Z60 [38], of each plant was recorded by tagging each plant on the wheat head where the first anther appeared. The time point to start the heat treatment, treatment temperature, and other settings were as per previous studies [18,23]. Specifically, on the 10th day after anthesis(DAA), three pots among six were moved into a growth chamber set to 37/27°C (day/night), 14-h photoperiod, and 420 mmol m-2s-1light intensity for 3-day heat treatment. Enough water was given to the plants to ensure there was no drought stress.The pots were returned to the glasshouse after heat treatment.
2.3. Scanning of samples
2.3.1. Seedling stage
Roots were scanned by WinRhizo scanner-based system(v.2007, Regent Instruments Inc.), which provided accurate and unbiased root trait values than manual measurements as shown in previous studies [15]. Root traits such as total root length, root surface area, root diameter and root volume were calculated by the software. Other seedling traits such as shoot length, seedling length and fresh and dry weight were also measured.
2.3.2. Adult stage
Chlorophyll contents (CC) of flag leaf on main stem were measured by a handheld portable chlorophyll meter (SPAD-502Plus;Konica Minolta,Osaka,Japan)before and after the treatment.Grain number per plant (GN) and grain weight per plant (Yield) were measured after harvest.
2.4. Statistical analysis
Screening data analysis was carried out based on measurements under heat stress (HS) and non-stress (NS) conditions, as below.
1) Seedling DI = (Root length under NS-Root length under HS)/Root length under NS
2) CC DI (Chlorophyll content damage index) = (CC at 10 DAA-CC at 13DAA)/CC at 10 DAA
3) GN (Grain number per pot) DI = (GN under NS-GN under HS)/GN under NS
4) Adult DI=Yield (grain weight per pot) DI = (Yield under NS-Yield under HS)/Yield under NS
Statistical analyses were undertaken using non-parametric test(Mann-WhitneyUtest)to compare phenotypic variations and confirm significant differences between NS and HS treatments. The relationship between heat-tolerant performances at different growth stages was analysed by simple linear regression.
2.5. Genotyping comparison
The most tolerant and most susceptible genotypes were selected and divided, as two extreme groups - extreme tolerant group with consistent tolerant performances from seedling stage to adult stage and extreme susceptible group for their susceptible performances at the two stages.Genotypes in the two groups were subjected to Infinium Wheat Barley 40K v1.0 BeadChip (https://www.illumina.com/) for SNP polymorphic alleles between the two groups.Particularly,SNP clustering and genotype calling were performed using GenomeStudio 2.0 software(Illumina).SNPs with a call frequency <0.8 (i.e., missing data points >20%), minor allele frequency(MAF)<0.05 or heterozygous calls >0.25 were removed.The identified SNPs that genetically distinguished the tolerant group and susceptible group were worked as anchors to perform a BLAST for searching candidate gene(s) against the wheat reference genome[40].The searching range was between SNP physical position±2 Mb, as affected genes were often up to 2 Mb from the associated SNP [41].
2.6. Expression identification
The expression information of the candidate gene(s) was searched to further validate the identified genes by the expVIP tool that available at http://www.wheat-expression.com. Two points were mainly focused when utilising the information for validation.Firstly, whether the expression of the particular gene(s) was related to heat stress; secondly, in what tissues if the gene(s)expressed at a high level.
3. Results
3.1. Results from seedling screening of genotypes with known performances at adult stage
3.1.1. Roots responded to heat stress more obviously than shoots at seedling stage by continuous observation
Under non-stress condition, roots of wheat grew faster than shoots in the first few days and had similar length as shoots at the end of screening on the 7th day(Fig.2).Under heat stress,however, the roots were observed to be more severely influenced by heat than other organs in all the cultivars. Though the number of primary roots remained unchanged, the root lengths reduced considerably compared to the controls.Moreover, the growth of roots was much slower than shoots under high temperature from germination to the end of the 7th day for most genotypes. The final lengths of roots were much shorter than shoots. Therefore, root traits were focused on and further studied.
3.1.2. Significant differences of root traits under heat and non-heat stress conditions
Among the four root traits including total root length,root surface area,root diameter and root volume,the root length decreased the most under heat stress on an overall basis compared to control(Fig. S2). The decreased length ranged from 3.87 cm (Kennedy) to 40.44 cm (Zebu). Nineteen cultivars decreased their total root lengths significantly while five cultivars showed no significant differences. The second most impacted trait by heat stress was root surface area, 21 cultivars decreased their root surface areas significantly under heat stress and three cultivars showed no significant differences. The root surface area decreased with a range from 1.05 cm2(Westonia)to 5.93 cm2(Halberd).Root diameter was relatively less impacted by heat stress compared to the first two traits.Under heat stress condition,one cultivar‘‘Yitpi”had a significantly increased root diameter of 0.19 cm, whereas five cultivars including ‘‘Halberd”, ‘‘Gladius”, ‘‘Kennedy”, ‘‘Sunleg” and ‘‘Clearified JNZ” had significantly decreased root diameters and other 18 cultivars showed no significant differences. For the last trait, the root volume, four cultivars, namely ‘‘Frame”, ‘‘Espada”, ‘‘Gile” and‘‘Tamarin Rock”,showed no significant changes whereas 20 others had significantly decreased root volumes under heat stress.
3.1.3. Rank cultivars for their seedling DI
3.2. Results from adult stage screening of genotypes with known performances at seedling stage
No significant changes of chlorophyll content after heat stress treatment were found for five cultivars, namely ‘‘Perenjori”,‘‘Mitre”, ‘‘Nobre”, ‘‘Ananda” and ‘‘Brazil”. While for the other 11 genotypes, the chlorophyll content decreased significantly after treatment (Fig. S4). Two genotypes ‘‘Isareal A” and ‘‘Cascades”showed increased GN) with shrinking grain size, One genotype‘‘Brazil 32”showed significantly decrease of GN,there were no significant changes on GN for other genotypes. Yield of four cultivars decreased under heat stress treatment, including ‘‘Brazil 32”,‘‘Blanco Sin Barbillas”, ‘‘Cascades” and ‘‘Stilletto”, no significant changes were found for other cultivars.
3.3. Traits relationships at the two stages
3.3.1. Root length and yield were selected as representative traits to investigate correlation of HT between the two growth stages

Fig.3. Relationship of heat tolerance at seedling stage and adult stage.Forty genotypes were plotted by blue dots.The x-axis represented seedling DI and y-axis represented adult DI. The linear trend line was shown by dotted blue line.
As total root length was the most reduced trait under heat stress with large variation among various cultivars,it was selected as a representative trait to calculate DI to rank the cultivars for HT at seedling stage.Meanwhile,due to the importance of yield,it was selected as the representative trait to calculate DI to score cultivars for HT performances at adult stage. The relationship of HT at the two growth stages was established based on these two traits.
3.3.2. Relationship of heat tolerance at seedling stage and adult stage
All 40 tested wheat cultivars were plotted together for their HT DI both at seedling and adult stages, to investigate relationship of HT performances at two different stages (Fig. 3).
A general linear relationship was found based on the scatter plot with anr=0.6930 (P<0.01), showing that the seedling HT performances had a positive relationship with performances at adult stage. The more tolerant a cultivar was at seedling stage,the more likely it would be tolerant at adult stage. For example,among the extreme tolerant cultivars of the top 8 identified and selected at seedling stage, ‘‘Perenjori”, ‘‘Mitre”, ‘‘W156” and ‘‘Tevere”turned out to be also tolerant at adult stage for yield.The other four cultivars including ‘‘Ananda”, ‘‘India 227”, ‘‘Arrino” and‘‘Nobre” at least showed medium tolerance at adult stage. It suggested the possibility of selecting at early seedling stage for the breeding of adult stage tolerance.
3.4. Molecular basis of the relationship of HT at seedling stage and adult stage
3.4.1. SNP markers identified for cultivars tolerant at both seedling stage and adult stage
Extremely tolerant group including ‘‘Perenjori”, ‘‘Mitre” and‘‘W156” (Fig. S4) and extreme susceptible group including ‘‘Maravilla”, ‘‘Brazil 32” and ‘‘Blanco Sin Barbillas” were compared for SNP polymorphism between them and two SNPs were identified(Table 1). Finally, a total of 82 high-confidence genes were found.
The Prince, far more attracted by the thoughts of possessing the Princess than her promised dower, set forth39 in his quest after taking leave of the King and Queen, the latter giving him a miniature of her daughter which she was in the habit of wearing
3.4.2. Previously reported genes among the candidate genes
Among the total 82 candidate genes,eleven previously reported genes were focused on and summarized (Table 2) for their information, such as their annotation description, related traits and stress in the publications. The function annotation revealed that they all encoding specific proteins with particular molecular functions, which were probably played key roles during the process of wheat growth under heat stress from seedling stage to adult stage.GenesTraesCS1A01G072200andTraesCS1A01G056200have been previously found related to leaf senescence under drought stress[42],TraesCS1A01G051600related to fertility conversion under photo/thermo stress [43],TraesCS1A01G070600,TraesCS1A01G051800,TraesCS1A01G053000,TraesCS1A01G053100andTraesCS1A01G056700related to deoxynivalenol content or cell wall and plant-type secondary cell wall biogenesis afterfusarium graminearuminfection [44,45],TraesCS1A01G051800related to powdery mildew infection [46]. Two genesTraesCS1A01G069600andTraesCS1A01G053300were previously identified by others not under particular biotic or abiotic stress, however,TraesCS1A01G069600was related to root traits including total root length,andTraesCS1A01G053300was related to protein and starch quality,which are important to final yield [47,48].
3.4.3. Expression validation
By the expression information (Fig. S5), it was shown that the expressions of the 11 genes were all related to heat stress. The wheat shoots and leaves were the issues where these genes expressed at a high level.
4. Discussion
Traits conferring HT must be explored thoroughly to maximise germplasm exploitation[49].There was no previous report on relationship about heat tolerant performances at different growth stages in wheat. This study extended the existing research outcomes at individual stages and further investigated how the identified seedling tolerant/susceptible germplasm respond to heat stress at the reproductive stage, and how the reported reproductive-stage tolerant/susceptible germplasm respond to heat at the seedling stage. It could provide an important guidance for breeding practice.
One question this study answered is that ‘‘what” the relationship of heat-tolerance at different stages is, which is important for breeding selection and prediction. In this study, a general positive relationship of heat tolerance at seedling stage and at adult stage was established by DI that was calculated from seedling root length and final yield, suggesting tolerant genotypes at seedling stage tend to be tolerant at adult stage. Among eight extreme seedling-tolerant cultivars ‘‘Perenjori”, ‘‘Tevere”, ‘‘Arrino”, ‘‘India 227”, ‘‘W156”, ‘‘Nobre”, ‘‘Ananda”, ‘‘Mitre”, four of them including‘‘Perenjori”,‘‘Tevere”,‘‘W156”,‘‘Mitre”showed premier heat tolerance at adult stage(lowest adult DI in Fig.3)than other genotypes in the collection, the other four cultivars namely ‘‘Arrino”, ‘‘India 227”, ‘‘Nobre”, ‘‘Ananda” that performed well at seedling stage with at least a medium rank (with medium adult DI in Fig. 3) for adult performance, with still smaller adult DI (better performance at adult stage) than check (control) cultivar ‘‘Mace”. The National Variety Trials(NVT)program by GRDC(Grains Research&Development Corporation) is the largest independent co-ordinated trial network in Australia. According to statistics from NVT [50], wheat cultivar ‘‘Mace” is the most widely adopted variety in Western Australia (local production). It has been considered a benchmark variety for yield since its release in 2008.Therefore,when releasing and managing any variety, it will normally be compared with‘‘Mace” for consideration of the yield and other important agronomical traits such as varieties maturity, biotic and abiotic tolerance. In this study, ‘‘Mace” was also tested and we suggested it can also be regarded as a check (control) or benchmark for good or bad heat-tolerance performance as it has a medium seedling DI and a medium adult DI (middle position in Fig. 3).
By putting all the tested cultivars into a two-way chart (one way stands for seedling stage and one way stands for adult stage),performances of various genotype under heat stress at different growth stages showed a trend of positive association.In the future,if more genotypes from a broader origin and source could be involved into this chart to make it more accurate, then the yield performance of a germplasm under heat stress could be predicted by curve fitting. Prediction accuracy for wheat breeding within a large panel was reported to be substantially higher than observed in smaller populations[51]. If early selection at seedling stage can be carried out based on a standard curve established by a large quantities of wheat varieties, then seedling performance of a particular wheat genotype can be used to select genotypes for checking adult stage performance in greenhouse or phytotron, and further tested in field for production to validate real performance of heat tolerance.
The implication of above‘‘early selection”strategy is that it will accelerate the breeding programme for HT by reducing workload and saving time for selection process. Conventional crop breeding are time taken, requiring at least 5-6 generations before assessments, often up to 11-13 years between making crosses to new cultivar release [52]. If extreme tolerant genotypes could be selected at seedling stage, first, workload of extensive selection will be reduced by focusing on extreme genotypes with superior seedling performances. Meanwhile, breeders do not have to wait until harvest to identify HT performance of each generation, thus a series of steps in breeding process such as parent selection,cross and selection at F3-F4, will speed up. Of course, final yield performance has to be validated on F5and later generations for adult selection. With the fast generation technology [53] and seedling selection of HT, time taken for the production of new cultivars could be easily halved to 5-6 years.
Another question this study answered is why the positive relationship exists between seedling tolerance and adult stages for heat tolerance in wheat. It was reported that some well-known heat-induced genes at reproductive stage such asTaHSF1,TaHSF4,TaMBF1c,TaHSP17.4, andTaHSP101were found significantly enhanced in acetylation levels in wheat seedlings[35].The results from this study found the similar trend. From the result of this study, we speculate that heat tolerance at seedling stage probably related to the ability of the varieties to establish healthy root system which can maintain at adult stage. Roots are vital for plants because roots directly contact with the soil environment and are responsible for taking up nutrients and water [15]. Furthermore,we also speculate that genotypes showed consistent good performances under heat stress at both stages may be because the highly expressed genes are similar at the two stages. A common mechanism of HT at seedling stage and adult stage was proposed,involving enzyme system, signal transduction, and degradation of toxic or harmful products under heat stress condition.
Genes encoding heat shock proteins(HSPs)were so far the most studied genes related to molecular responses under heat stress.Most of HSPs function as molecular chaperones in maintaining homeostasis of protein folding and acquisition of thermostolerance [54-56]. The identified candidate genes distinguishing extreme heat tolerant genotypes and susceptible genotypes in this study were all encoding specific proteins with particular molecular functions, such as enzyme protein or structure protein etc., and protein homeostasis under heat stress including protein quality control, effective elimination of toxic proteins, and translational regulation was reported especially important for rice thermostolerance [57], so it was deduced that the tolerant genotypes performed better from seedling to adult stage than the susceptible genotypes mainly due to their advantages in: 1) Enzyme system and signal transduction. Signal transduction is the key part in the pathway of heat tolerance and kinases are particularly prominent in signal transduction and co-ordination of complex functions[58]. The identified geneTraesCS1A01G070600encoded kinasefamily protein,which may arouse the downstream pathway of heat tolerance. Dynamic and effective kinase/ enzyme system lead to unhindered signal transduction and the differences in their efficiency of activities may lead to tolerance or susceptibility under heat stress; 2) Protein structure retention. Full activation of proteins requires multiple-site phosphorylation on their conserved region [59]. The identified genes related to Zinc finger domaincontaining protein,F-box family protein etc.may helped the tolerant genotypes superior in structure retention for conserved and functional region; and 3) Clearing toxic or harmful products.TraesCS1A01G051800encoded aluminium sensitive protein andTraesCS1A01G056700encoded heavy metal transport/detoxification protein.TraesCS1A01G056200was annotated as ALMT1-M39.2. An ALMT1 (Al-activated malate transporter) gene cluster was once reported controlling aluminum tolerance of rye [60]. It was deduced that the tolerant wheat genotypes were more sensitive and efficient in clearing waste that produced in cell under heat stress condition through better function of detoxification.
The better a cultivar performed in these aspects,the more tolerance under heat stress this cultivar might show.Above mechanism was proposed to explain why some cultivars could be tolerant from seedling stage to adult stage,and why it is possible to select tolerant genotypes from early stage on a genetic aspect. In summary,the main reasons might be that tolerant genotypes generally have a more effective enzyme system and unhindered signal transduction, and more capability to degrade toxic or harmful products.
This study tested the seedling-adult relationship for heat tolerance mainly on Australian spring cultivars, a wider range of cultivars from other countries need to be tested further to find a more explicit trend. Further studies are necessary to check the expression patterns and potential natural genetic variations of these target genes in different wheat accessions,especially in the extremely tolerant and susceptible genotypes. Moreover, the SNPs identified in this study can be converted to KASP (Kompetitive Allele-Specific PCR) markers or PCR based markers for validation in a wide range of nature populations and cross populations to make them more practically useful in wheat breeding.
5. Conclusions
A total of 40 wheat genotypes were evaluated for their heattolerant performances at seedling and adult growth stages. Heattolerant genotypes at seedling stage showed less root length decrease under heat stress, and genotypes tolerant at adult stage showed high yield under heat stress. A significant linear positive relationship was found between HT performances at seedling stage and at adult stage.Two SNP markers and 11 candidate genes were identified associated with HT from seedling stage to adult stage.Expression patterns showed that the expressions of the 11 genes were all related to heat stress. A genetic mechanism explaining the superior tolerance was postulated.The result of this study suggested the similar controlling mechanism of HT may exist at different growth stages,and the possibility to select HT at seedling stage for the breeding of heat tolerance in wheat.
CRediT authorship contribution statement
Lu Lu:Conceptualization, Data curation, Formal analysis, Writing - original draft.Hui Liu:Funding acquisition, Methodology,Resources, Software, Formal analysis, Supervision, Writing -review&editing.Yu Wu:Supervision.Guijun Yan:Funding acquisition,Methodology,Formal analysis,Resources,Supervision,Writing - review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This research was funded by the Global Innovation Linkages Project (GIL53853) from the Australian Department of Industry,Science, Energy and Resources. The authors thank Dr. Daniel Mullan at InterGrain Pty Ltd for his kind help in genotyping using Infinium Wheat Barley 40K v1.0 BeadChip.
Appendix A. Supplementary data
Supplementary data for this article can be found online at https://doi.org/10.1016/j.cj.2022.01.005.

- The Crop Journal的其它文章
- Brief Guide for Authors
- Revisiting the role of delta-1-pyrroline-5-carboxylate synthetase in drought-tolerant crop breeding
- A new gain-of-function OsGS2/GRF4 allele generated by CRISPR/Cas9 genome editing increases rice grain size and yield
- Influence of seven levels of chemical/biostimulator protection on amino acid profile and yield traits in wheat
- Corrigendum to ‘‘De novo design of future rapeseed crops: Challenges and opportunities” [Crop J. 10 (2022) 587-596]
- Improving the resistance of the rice PTGMS line Feng39S by pyramiding blast, bacterial blight, and brown planthopper resistance genes