
4 种薯蓣属植物GBSS 基因的电子克隆与生物信息学分析
2022-08-11 14:02陈知龙郑佳豪吴坤龙张壹
浙江农业科学 2022年8期
陈知龙 ,郑佳豪 ,吴坤龙 ,张壹
(1.文成县现代农业与康养产业研究院,浙江 文成 325300;2.文成县科学技术局,浙江 文成 325300)
薯蓣属 (Dioscorea) 是被子植物门单子叶植物纲薯蓣目薯蓣科下的一个属,目前该属已知植物有600 种以上,多为缠绕藤本,有地下块状或根状块茎是该属的主要特征[1]。薯蓣属植物中许多种类具有重要的经济价值,如温带地区普遍栽种的薯蓣(D.opposita),热带和亚热带地区广泛栽种的参薯(D.alata)、甜薯 (D.esculenta) 可以供食用和药用;盾叶薯蓣 (D.zingibernsis)、穿龙薯蓣(D.nipponica)、黄山药 (D.panthaica) 等植物的根状茎中含有薯蓣皂苷元,是合成避孕药及生产甾体激素类药物的重要原料;薯莨 (D.cirrhosa)的块茎富含缩合性鞣质和酚类化合物,常用于止血、治疗溃疡、抗菌消炎,也可用于制革等。
淀粉是薯蓣属植物块茎的重要组成成分,山药块茎中的淀粉可占干物质的50%~80%[2]。淀粉有直链淀粉和支链淀粉两种形态,在淀粉合成过程中,颗粒结合淀粉合成酶 (granule-bound starch synthase,GBSS) 是控制直链淀粉合成的关键蛋白质,负责催化生成线性多聚葡萄糖分子[3]。该蛋白由GBSS基因来编码,该基因也称糯性基因(Waxy),或蜡质基因。GBSS基因广泛存在于大麦、小麦、水稻、玉米、番茄、马铃薯等作物中,且已被克隆。研究[4]已证明Waxy基因突变导致胚乳中直链淀粉和支链淀粉含量及比例发生变化,是禾谷类作物糯性形成的根本原因。对于薯蓣属植物GBSS基因的研究还鲜有报道,仅吴志刚等[5]克隆了参薯种糯性栽培品系-文成糯米山药的GBSS基因序列,并进行了初步的基因结构特征及系统进化分析。本研究主要通过生物信息学分析,比较分析参薯、几内亚白山药、灌丛薯蓣和盾叶薯蓣4 种薯蓣属植物GBSS基因结构,蛋白质结构及系统发育特点。研究结果将为进一步阐述薯蓣属植物GBSS基因及其编码蛋白调控淀粉合成的分子机制提供科学依据。
1 材料与方法
1.1 电子克隆
以文成糯米山药GBSS基因序列为探针,BLAST 检索美国国立生物技术信息中心 (National center for biotechnology information,NCBI,https://www.ncbi.nlm.nih.gov/) 中的参薯[D.alata(taxid:55 571)]、几内亚白山药[D.rotundata(taxid:55 577)]、灌丛薯蓣[D.dumetorum(taxid:167 584)]、盾叶薯蓣[D.zingibernsis(axid:325984)] 基因组数据库,同源比对获得4 种薯蓣属植物GBSS基因同源序列。
1.2 基因结构与GBSS 蛋白结构分析
以NCBI 数据库中已注释的玉米、番茄、马铃薯等植物GBSS基因结构为参考,使用同源比对方法分析5 种薯蓣属植物的GBSS基因结构。利用mVISTA (http://genome.lbl.gov/vista/index.shtml)程序比较分析5 种薯蓣属植物GBSS基因组序列同源 性。利用 Interpro 数据库 (http://www.ebi.ac.uk/interpro/) 对GBSS 蛋白进行蛋白质家族和结构功能域分析。
1.3 GBSS 蛋白系统进化分析
从 NCBI 数据库检索获取小麦 (Triticum aestivum,AAL05405)、大麦 (Hordeumvulgare,AAM74048)、节节麦 (Aegilopstauschii,XP _020188467)、水稻 (Oryzasativa,XP_015644489)、小米 (Setariaitalica,NP_001267750)、玉米 (Zea mays,NP _001358937)、高粱 (Sorghumbicolor,XP_002436418)、铁皮石斛 (Dendrobiumcatenatum,XP _ 028549213)、油棕 (Elaeisguineensis,XP _010940833)、芭蕉(Musaacuminata,XP_009415991)、茶树 (Camelliasinensis,XP_028108084)、拟南芥(Arabidopsisthaliana,NP _ 174566)、番茄(Solanumlycopersicum,NP _001311458)、马铃薯(Solanumtuberosum,NP _ 001274918)、烟草(Nicotianatabacum,NP _ 001311577)、辣椒(Capsicumannuum,XP _ 016564900)、牵牛花(Ipomoeatriloba,XP _ 031124794)、羽扇豆(Lupinusangustifolius,XP _019431273)、百脉根(Lotusjaponicus,ACB30384)、苹果 (Malus domestica,NP_001280924)、油桃 (Prunuspersica,XP_007211283) 和木薯 (Manihotesculenta,XP_021603997) 22 种被子植物的GBSS 蛋白序列。利用ClustalX 2.0 软件进行蛋白质的多重序列比对,MEGA7 软件邻接法构建系统发育树,并对生成的系统发育树进行 Bootstrap 校正,循环次数为1 000。
2 结果与分析
2.1 GBSS 基因结构分析
以文成糯米山药GBSS基因序列为探针,通过同源比对方法获得参薯、几内亚白山药、灌丛薯蓣和盾叶薯蓣GBSS基因的基因组核苷酸序列。参照玉米、番茄、马铃薯等已详细注释的GBSS基因结构,同源比对分析获得5 种薯蓣属植物的GBSS基因结构 (表1)。从表中我们可以看出5 种薯蓣属植物的GBSS基因结构高度相似,都包含13 个可编码蛋白质的外显子和12 个内含子,其中第2~12外显子的大小在所列物种间高度保守。
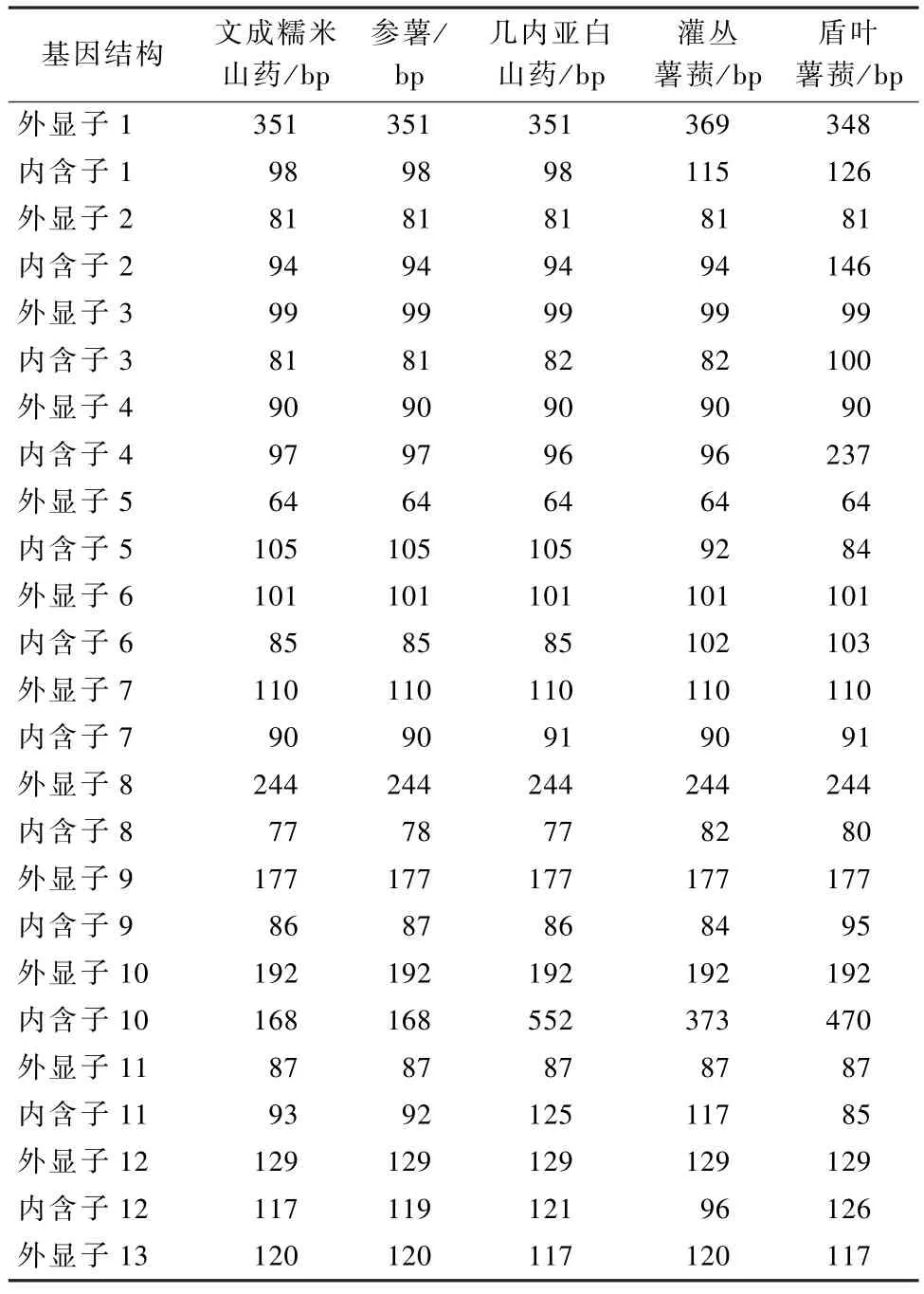
表1 薯蓣属植物GBSS 基因结构分析
以文成糯米山药GBSS基因组核苷酸序列为参照,比较分析参薯、几内亚白山药、灌丛薯蓣和盾叶薯蓣与文成糯米山药GBSS基因组核苷酸序列的同源性差异 (图1)。从图中可以发现,不管是外显子序列还是内含子序列,文成糯米山药的GBSS基因序列均与参薯高度同源,这也一定程度说明了文成糯米山药是参薯种的一个地方栽培品系。盾叶薯蓣的GBSS基因序列与文成糯米山药保守性稍差,但其外显子序列的保守性明显高于内含子。

图1 薯蓣属植物GBSS 基因组序列同源性比较分析
2.2 GBSS 蛋白结构分析
参薯、几内亚白山药、灌丛薯蓣和盾叶薯蓣GBSS基因的编码区核苷酸长度依次为1 845、1 842、1 863 和1 839 bp,分别编 码614、613、620 和612 个氨基酸。4 种薯蓣属植物GBSS基因的编码区核苷酸序列与文成糯米山药的同源性分别为99.3%、97.4%、90.5% 和85.4%,其 编码的GBSS 蛋白氨基酸序列与文成糯米山药的同源性分别为98.7%、95.3%、86.3%和81.9%。
GBSS 蛋白属于淀粉合成酶类蛋白质,除N 端含有一段70 多个氨基酸组成的质体转运肽外,还包含一个淀粉合成酶催化结构域和一个糖基转移酶结构域 (图2)。选取玉米、小米为单子叶代表植物,拟南芥、马铃薯为双子叶代表植物,与5 种薯蓣属植物GBSS 蛋白进行氨基酸序列同源性比对分析。从图3 中可以看出,GBSS 蛋白N 端质体转运肽的保守性较差,但淀粉合成酶催化结构域和糖基转移酶结构域高度保守,这说明GBSS基因的淀粉合成功能在植物进化过程中高度保守。

图2 GBSS 蛋白结构域分析

图3 GBSS 蛋白氨基酸序列分析
2.3 GBSS 蛋白系统进化分析
为了分析不同薯蓣属植物的GBSS 蛋白之间,以及与其他植物GBSS 蛋白间的进化关系,我们选取5 种薯蓣属植物以及玉米、小米、拟南芥、马铃薯等22 种其他植物的GBSS 蛋白进行同源比对,构建系统进化树 (图4)。从进化角度上看,不同植物的GBSS 蛋白在进化过程中可分为单子叶植物和双子叶植物两个分支。文成糯米山药GBSS 蛋白在所列薯蓣属植物中与参薯GBSS 蛋白的亲缘关系最近,与盾叶薯蓣GBSS 蛋白亲缘关系最远。薯蓣属植物GBSS 蛋白与属于天门冬目植物的铁皮石斛GBSS 蛋白的进化距离较近,与油棕目、姜目、禾本目植物的GBSS 蛋白都属于单子叶植物进化分支。
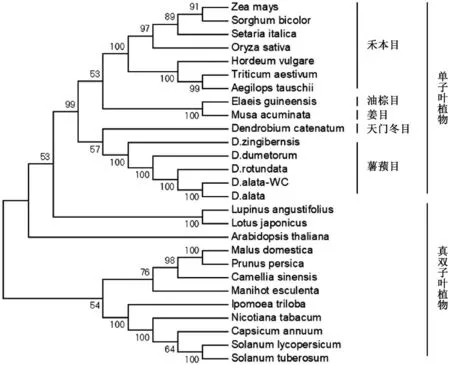
图4 多种植物GBSS 蛋白的系统进化分析
3 讨论
淀粉是高等植物光合产物主要的储存形式,可分为直链淀粉和支链淀粉。直链淀粉和支链淀粉的相对含量可以影响淀粉的口感、黏性、糊化性以及回生性等特性[6]。植物中有多种酶参与淀粉的合成,其中颗粒结合淀粉合成酶 (GBSS) 被认为在直链淀粉的延长中扮演着重要角色[3],它催化形成α-1,4-糖苷键,将葡萄糖单元连接在α-1,4-葡聚糖的非还原端,延长α-1,4-葡聚糖链。
GBSS 蛋白不能正常表达或活性下降是禾谷类作物糯性形成的根本原因。研究已证明,糯稻主要是由GBSSI基因第1 内含子5′端G/T 突变和第2外显子23 bp 碱基片段插入,引起前体mRNA 不能正常剪切,导致GBSSI表达量减少所形成的[7]。糯玉米的两种缺失突变体Wx-D7 和Wx-D10 分别是由于GBSSI基因第7 外显子和第10 外显子内碱基缺失造成GBSSI 蛋白功能丧失形成的[8-9]。糯性大麦GBSSI基因启动子区397 bp 碱基缺失则造成了基因启动子活性比非糯性大麦弱[10]。控制小麦GBSS 蛋白表达的Wx-A1、Wx-B1、Wx-D1 3 个等位基因发生突变都会影响小麦的糯性,其中缺少3 个Wx 亚基就会得到糯小麦[11]。
利用现代生物技术突变GBSS基因或降低GBSS 蛋白表达也可以使作物获得糯性特点。利用RNA 沉默策略转化小麦,降低GBSS 蛋白表达可以显著下降小麦籽粒胚乳中直链淀粉含量[12]。利用CRISPR/Cas9 基因编辑技术对水稻Waxy基因的外显子区核苷酸序列实现定点突变,可以将直链淀粉含量从亲本的15.5%降到2.0%~2.6%,接近或达到糯稻水平[13]。马铃薯GBSS4 个等位基因经CRISPR/Cas9 基因编辑均产生突变的突变体则完全缺少了直链淀粉的合成[14]。淀粉也是薯蓣属植物地下块茎的重要组成成分,因此,深入解析薯蓣属植物GBSS基因的结构和蛋白结构特点,可为今后调控薯蓣属植物直链和支链淀粉合成,以满足食品、医药、工业等行业对不同性质淀粉的需要提供科学依据。
猜你喜欢
基层中医药(2022年2期)2022-07-22
中南药学(2022年2期)2022-03-30
作文周刊·小学二年级版(2022年12期)2022-03-19
包装工程(2022年1期)2022-01-26
中国农业科学(2021年7期)2021-04-21
天然产物研究与开发(2020年2期)2020-04-27
农产品加工(2020年3期)2020-03-11
大众文艺(2019年16期)2019-08-24
科技知识动漫(2017年8期)2017-08-09
科技知识动漫(2017年5期)2017-05-11
