
春小麦开花期光合参数对土壤水分和温度变化的响应
2022-08-10 05:45雷俊张健赵福年齐月张秀云李强尚军林
生态环境学报 2022年6期
雷俊 ,张健,赵福年,齐月,张秀云,李强,尚军林
1.中国气象局兰州干旱气象研究所/甘肃省干旱气候变化与减灾重点实验室/中国气象局干旱气候变化与减灾重点开放实验室,甘肃 兰州 730020;2.定西市气象局,甘肃 定西 743000;3.定西市农业科学研究院,甘肃 定西 743000;4.兰州资源环境职业技术大学,甘肃 兰州 730021
小麦是中国的三大粮食作物之一,小麦的丰歉对全国粮食安全有着举足轻重的作用。中国小麦总产量约占世界粮食作物总产量的25%以上,而干旱半干旱区小麦的生产量占全国生产总量的 50%以上,占有十分重要的地位(张凯等,2015)。春小麦的生长发育不仅受自身遗传物质的控制,同时也受温度、湿度、光照、土壤、二氧化碳浓度等诸多环境因子的影响。
随着工业化的发展和人类活动的影响,近百年来全球气候变暖已毋庸置疑,1880—2012年,全球平均地表温度升高了 0.85 ℃,气候变暖的趋势特征几乎在全球各地均可以观测到(IPCC,2013)。气候变暖引起全球降水格局的改变,从而对作物根系土壤水分供给将产生重要的影响。同时,温度作为作物生长发育的关键气象因子之一,其升高在加速作物同化作用的同时,也使得异化作用增强,影响生物量的积累和产量的形成。目前,有关气候变暖对农业发展、农业生态的影响已成为社会各界关注的热点(赵鸿等,2016;姚玉璧等,2018)。
光合作用是植物利用光能固定大气中CO2合成有机物的生物过程(梁星云等,2017),是作物生长发育和产量形成的生理基础,同时也是全球碳循环及其他物质循环的重要环节(孙彩霞等,2010)。光合作用产生的物质通过转化能产生90%—95%的干物质,光合能力的强弱决定了作物产量和品质的高低(董祥开等,2008)。研究表明,植物光合生理过程与自身因素如叶片结构和生理机能等密切相关,又受光强、气温、CO2浓度、湿度、土壤含水量等外界环境因子的影响(Sofo et al.,2009;张振文等,2010),并随环境条件的改变呈现出一定的变化规律。光合参数是反映植物对逆境生理过程响应的主要指标,有助于阐明植物对环境变化的生理适应性(靳欣等,2011;苏华等,2012)。
已有研究表明,春小麦产量和品质与气象条件密切相关,气温变化对春小麦不孕小穗数、穗粒重和成穗率有显著影响,不同生育期对气温变化的敏感性存在差异(姚玉璧等,2011;齐月等,2019)。温度升高1.0—2.5 ℃,春小麦三叶期净光合速率下降,拔节期、开花期净光合速率一致升高(肖国举等,2011);增温0.5 ℃,苗期-拔节期叶面积、叶绿素含量、净光合速率、水分利用效率一致上升,而增温1.0—2.0 ℃变化则相反,表现为一致下降趋势。张凯等(2015)研究表明,增温对春小麦株高和穗分配系数的影响表现为负效应,在相同的水分条件下,增温幅度越大,产量降幅越高(王鹤龄等,2015)。任鹏等(2010)研究表明,春小麦花后高温,旗叶净光合速率下降,产量下降。马莉等(2018)研究表明,灌溉量增加,春小麦根际可利用的有效水分增多,叶片光合能力呈现增加趋势,且灌浆前期大于后期。
目前,国内外学者对气象条件对春小麦光合生理特性、生产力和产量等的影响进行了大量的研究(Asseng et al.,2011;Stupko et al.,2011;Dimina et al.,2018;姚玉璧等,2011;齐月等,2019);而黄土高原半干旱区作为中国典型雨养农业区,也是气候变化的敏感地带,有关土壤水分和温度变化对春小麦光合生理特性影响的定量研究较少。本文拟通过控制试验,揭示春小麦开花期旗叶光合生理参数对水分以及温度变化的响应特征,以初步掌握作物在不同环境因子控制背景下特定因子变动引起的光合特性的变化规律,为黄土高原半干旱区春小麦水分的高效利用提供理论依据。
1 材料与方法
1.1 研究区概况
本试验在中国气象局兰州干旱气象研究所定西干旱气象与生态环境试验站内进行,该站位于甘肃省定西市安定区西川农业园区(104°37′E,35°35′N),海拔 1920 m,年平均气温 6.3 ℃,年日照时数2500 h。该区年降水量386 mm,且分布不均匀,主要集中在7—9月。供试土壤为黄绵土,土质疏松,易耕作,但土壤贫瘠,持水能力弱,在蒸发较大时,土壤易受干旱威胁。
1.2 试验设计与方法
1.2.1 试验设计
试验春小麦品种为“定西40号”,该品种抗旱抗病性强,幼苗半匍匐,叶色灰绿,根系发达,苗期生长旺盛,增产潜力大,水分利用效率高,为当地常种品种。春小麦在2019年3月20日播种,于同年7月24日收获,全生育期126 d。试验布设8个小区,每个小区面积为1 m×2 m,中间用深度为2 m的水泥板隔开,防止不同小区间水分横向输送。试验设置两个不同的水分处理,每个处理4个重复,处理A为对照,在春小麦全生育期保证各小区土壤水分保持田间持水量的60%—65%。处理B为控水处理,在开花期各小区土壤水分保持田间持水量的40%—45%,乳熟期恢复灌水,保证土壤水分与对照相同,保持田间持水量的60%—65%。试验期间应用EM50自动土壤水分监测系统对0—50 cm深度土壤水分实时监测,并及时灌水。应用Li-6400XT光合作用观测系统测定时进行温度控制,分别控制气室温度为25、26以及27 ℃。
1.2.2 测定项目和方法
在春小麦播后第 90、91天(控水后第 20、21天)进行光合观测,此时春小麦为开花期,营养生长和生殖生长并进,需水量大,对水分变化敏感。利用美国Li-COR公司生产的Li-6400XT光合作用观测系统测定春小麦旗叶净光合速率(Net photosynthetic rate,Pn)、气孔导度(Conductance to H2O,Gs)、胞间CO2浓度(Intercellular CO2concentration,Ci)、蒸腾速率(Transpiration rate,Tr)等气体交换参数;气孔限制值(Ls)的计算公式为:

式中:
Ci——胞间CO2浓度;
Ca——大气CO2浓度。
叶片水分利用效率(Water use efficiency,WUE,用Ewue表示)用净光合速率和蒸腾速率的比值来表示(王振华等,2015)。

式中:
Pn——净光合速率;
Tr——为蒸腾速率。
光响应曲线的测定应用Li-6400-02B红蓝光源提供不同强度光合有效辐射(Photosynthetically active radiation,PAR,单位:μmol·m-2·s-1),分别在0、20、40、70、100、150、200、300、400、600、900、1200、1500、1800、2100、2400 μmol·m-2·s-116个光点下测定开花期叶片净光合速率,控制叶室CO2摩尔分数 (390±1) μmol·mol-1,叶室温度分别为25、26及27 ℃(对照处理对应A-25、A-26及A-27;控水处理 (B) 对应 B-25、B-26及B-27),叶片在每个温度下,用 1200 μmol·m-2·s-1的 PAR 适应30—40 min,待仪器读数稳定后,进入自动测量程序,随PAR梯度由高到低进行测量,重复测定3次。
1.2.3 统计分析方法
光响应曲线采用非直角双曲线模型(Nonrectangular hyperbola equation)(Farquhar et al.,1982)进行拟合,其表达式为:
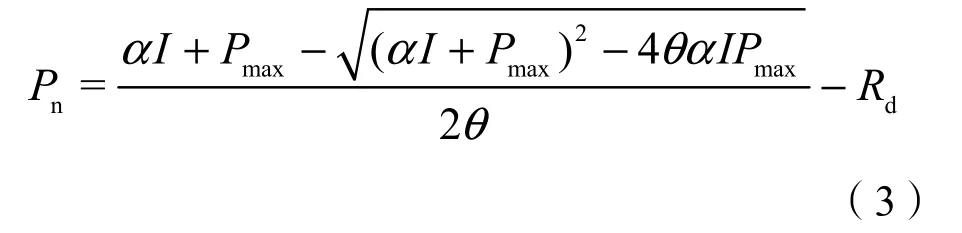
式中:
Pn——叶片净光合速率(μmol·m-2·s-1);
I——光合有效辐射(μmol·m-2·s-1);
θ——曲线的凸度(0<θ≤1),凸度越大,曲线的弯曲程度越大。在模拟光响应曲线时,凸度(θ)、最大净光合速率(Pmax)、表观量子效率(α)及暗呼吸速率(Rd)初始值分别设为0.5、30、0.05和2。其中参数θ、Pmax、α的范围分别设置为:θ≤1,Pmax≤50,α≤0.125,Rd不设定(刘宇锋等,2005)。
采用直线方程拟合弱光强条件下(≤200 μmol·m-2·s-1)的光响应数据得到表观量子效率(Apparent quantum efficiency,AQE,用Eaqe表示)、光补偿点(LCP)及光饱和点(LSP),通过求解直线方程式(4)得到(叶子飘等,2010):

当Pn=0时,计算得I值为LCP
当Pn=Amax时,计算得I为LSP
图形处理采用Origin 8.1软件,采用SPSS 16.0软件进行单因素方差分析(One-way ANOVA,SSR)、差异显著性检验(α=0.05)和光响应曲线的拟合,数据以平均值±标准差(Mean±SD)表示。
2 结果与分析
2.1 水分与温度对光合参数的影响
不同处理春小麦光响应曲线如图1所示。在一定光合有效辐射(PAR)范围内,不同处理春小麦净光合速率随PAR的增大而增大;对照处理(A)在 PAR 大于 700 μmol·m-2·s-1时,光合速率增速放缓;控水处理(B)在 PAR 大于 500 μmol·m-2·s-1时净光合速率增速放缓,光响应曲线趋于平缓,达到光饱和,A处理叶片达到光饱和的PAR阈值较B处理增大,增幅约为40%(表1)。在26 ℃和27 ℃条件下,B处理春小麦光合速率随PAR增大有减小趋势,光合作用表现出明显的光抑制。A处理叶片在PAR高值区所对应的光合速率明显大于B处理,表明水分不足对春小麦光合速率有较大的影响。

表1 不同处理春小麦开花期叶片光响应的特征参数Table 1 Characteristic parameters of the photosynthesis at the flowering stage of spring wheat among different treatments
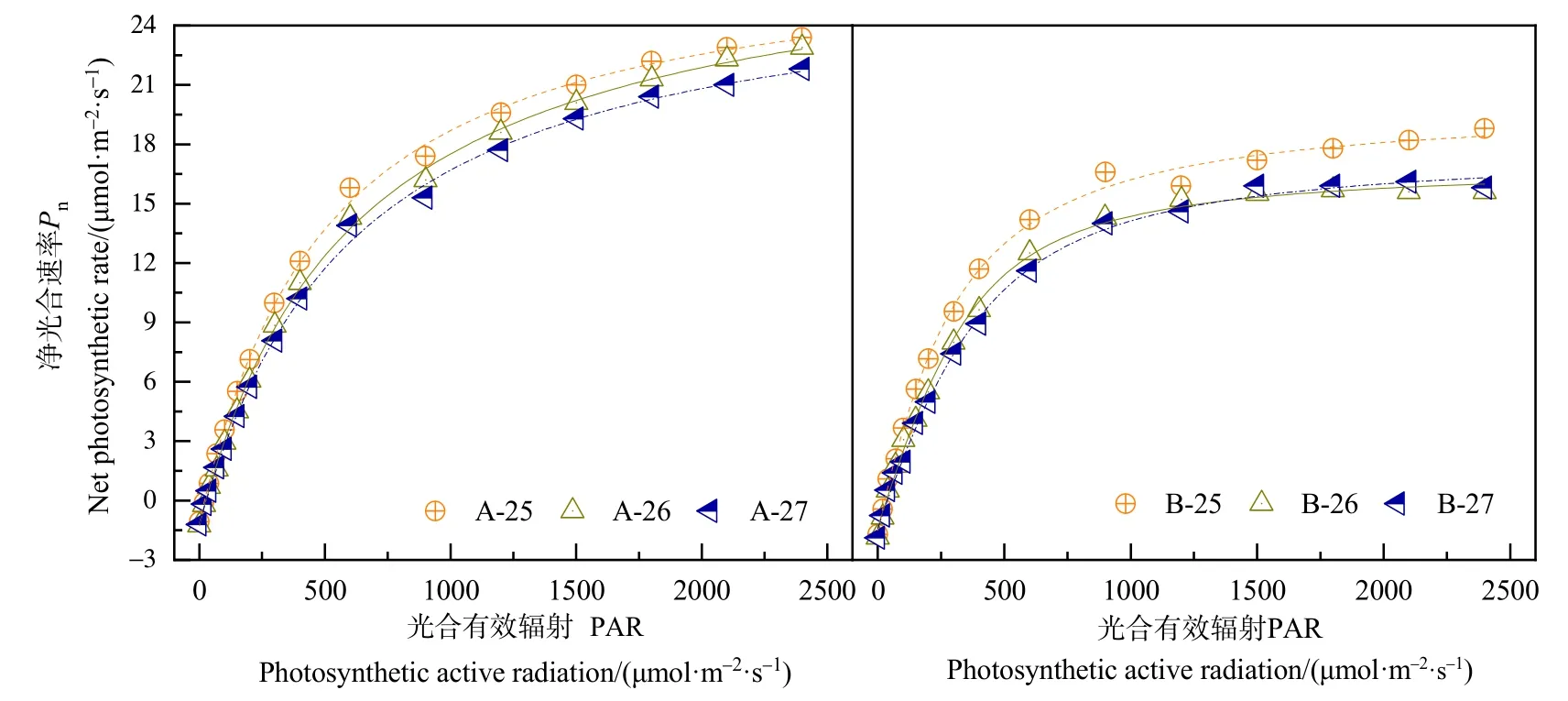
图1 不同处理春小麦开花期叶片光响应曲线比较Figure 1 Comparison of the light response curves at the flowering stage of spring wheat among different treatments
当 PAR 大于 500 μmol·m-2·s-1时,A 处理各个温度梯度之间净光合速率均差异明显,通过0.05显著性水平检验,在B处理春小麦各光强下,26 ℃和27 ℃处理之间差异较小,25 ℃条件下春小麦净光合速率与其他两个温度处理差异显著(P<0.05)。这可能是由于供水充足时,春小麦对温度的变化更敏感,通过其自身调节,及时响应PAR的变化而引起光合速率的变动。
比较两个处理不同温度下叶片光响应参数(表1)。A处理叶片表观量子效率(AQE)较B处理升高,平均增幅为28%,且温度越低,AQE越大。A处理叶片在25 ℃条件下较26 ℃、27 ℃分别增高16%和26%,A-25与A-27差异通过0.05水平显著性检验。B处理叶片在25 ℃条件下较26 ℃、27 ℃分别增高 13%和 19%,处理间未通过 0.05水平显著性检验。表明水分不足,温度升高,春小麦利用弱光的能力降低。A处理平均最大净光合速率(Pmax)较 B 处理升高 10.041 μmol·m-2·s-1,增幅为53%,可见水分对Pmax的影响较大。在水分充足时,Pmax随温度的变化不明显,处理间差异未通过显著性水平检验(P>0.05);水分不足,温度升高,Pmax明显降低。A处理春小麦光补偿点(LCP)在各个温度下均小于B处理;水分充足,27 ℃温度条件下LCP较25 ℃显著升高(P<0.05),增幅为15%;水分不足,随着叶片温度的升高,LCP明显增大,27 ℃、26 ℃温度条件下LCP显著高于25 ℃温度条件(P<0.05)。A处理叶片光饱和点(LSP)较B处理升高51%,且在各个温度条件下LSP较B处理差异显著(P<0.05),表明水分供给不足使春小麦利用弱光与强光的能力均降低。两个处理春小麦光补偿点均随温度的升高而逐渐增大,表明温度对春小麦利用弱光能力有一定的影响,温度越高,利用弱光能力越差。A、B处理春小麦光饱和点随温度的升高均有增大趋势,温度越高,光饱和点越高,表明较高的温度使春小麦利用强光的能力提高。A处理春小麦暗呼吸速率(Rd)在各个温度梯度下均高于B处理,水分供给充足,温度升高,Rd增大,水分供给不足,Rd随温度升高而降低。
2.2 水分与温度对春小麦气体交换参数的影响
2.2.1 水分和温度对蒸腾速率的影响
春小麦蒸腾速率(Tr)的光响应如图2a、b所示。在光合有效辐射(PAR)小于 500 μmol·m-2·s-1时,A、B处理春小麦Tr的增加幅度均较大,随着PAR的增大,A处理Tr持续快速增大;而B处理春小麦在26 ℃和27 ℃条件下,Tr变化较为平缓,表现为相对稳定的趋势。随着PAR的增大,A处理春小麦Tr在各个温度梯度下均大于B处理,表明水分供给对春小麦Tr有重要影响。比较不同温度条件下两个处理Tr的光响应过程,供水充足时,温度的变化对Tr影响较小,3个温度条件下大部PAR所对应的Tr之间差异不明显(P>0.05);在PAR大于1700 μmol·m-2·s-1时,25 ℃温度条件下Tr明显小于 26 ℃和27 ℃,但未通过0.05水平显著性检验。B处理春小麦在 PAR 大于 500 μmol·m-2·s-1后,随着 PAR的增大,25 ℃处理Tr不断增大,而26 ℃和27 ℃温度条件下Tr变化平缓,25 ℃温度条件下Tr明显大于26 ℃和27 ℃。
2.2.2 水分与温度对气孔导度(Gs)的影响
春小麦开花期气孔导度(Gs)的光响应如图2c、d所示。不同处理Gs随PAR的增加变化规律不完全一致,A处理Gs在3个温度梯度下均先快速增大,而后增速放缓,总体表现为持续增大的趋势。B处理在3个温度梯度下Gs先迅速增大,达到最大值,后又持续减小。A处理春小麦Gs以26 ℃最大,27 ℃最小,25 ℃和26 ℃差异不明显(P>0.05),27 ℃处理明显小于其他两个温度处理,通过 0.05水平显著性检验。B处理春小麦Gs在 25 ℃时最大,26 ℃和 27 ℃温度条件下,PAR大于 500 μmol·m-2·s-1,Gs差异不明显(P<0.05)。
2.2.3 水分与温度对胞间CO2浓度(Ci)的影响
春小麦开花期胞间CO2浓度(Ci)的光响应如图2e、f所示。不同处理春小麦Ci随光合有效辐射(PAR)的变化规律大致相同,均表现为在PAR小于 500 μmol·m-2·s-1时,Ci随 PAR 的增大而迅速减小;随着PAR的增大,Ci变幅减缓。B处理春小麦在PAR高值区,随着PAR的增加,Ci有微弱增加趋势。3 个温度梯度下,PAR 大于 500 μmol·m-2·s-1,A处理Ci的变化趋势表现为A-26>A-25>A-27,其中 A-25和 A-27之间差异较小;B处理春小麦在PAR 大于 800 μmol·m-2·s-1后,25 ℃条件下Ci明显大于其他两个温度处理,通过 0.01水平显著性检验;26 ℃和27 ℃处理之间差异不明显(P>0.05)。
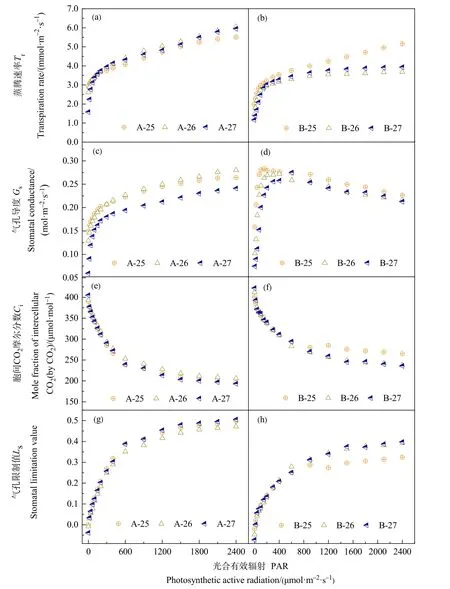
图2 不同处理春小麦开花期气体交换参数对光合有效辐射的响应Figure 2 Response of gas exchange parameters to photosynthetic active radiation of spring wheat at flowering stage under different treatments
2.2.4 水分与温度对气孔限制值(Ls)的影响
图2g、h为春小麦气孔限制值(Ls)对光强的响应。在低光强下,不同处理春小麦Ls均随PAR的增加而迅速增大,而后随着PAR的不断增大,Ls呈缓慢的增加趋势。随着PAR不断增加,B处理春小麦在PAR高值区,Ls有微弱的降低趋势,其中25 ℃条件下降低趋势最明显。比较不同温度梯度下春小麦Ls,结果表明,25 ℃条件下春小麦在PAR高值区Ls明显小于其他两个温度处理,而26 ℃和27 ℃之间则差异不大。A处理春小麦在PAR较弱时,各温度梯度间Ls差异不大;随着PAR的增大,26 ℃条件下Ls显著小于其他两个温度处理(P<0.05)。Ls随PAR的变化规律与Ci随PAR的变化规律相对应,且呈相反的变化趋势。
2.3 水分与温度对水分利用效率(WUE)的影响
不同处理叶片水分利用效率(WUE)随PAR的变化如图3所示。在弱光区,WUE随PAR的增大而迅速增大;随着PAR的进一步增大,不同处理春小麦在不同温度条件下 WUE的变化有所差异,A处理在 25 ℃条件下 WUE在 PAR高值区变幅缓慢,其值趋于平稳,而 26 ℃和 27 ℃条件下,在PAR 达到 1700 μmol·m-2·s-1前后 WUE 达到最大值,此后逐渐降低。比较A处理不同温度梯度WUE的变化,结果表明,25 ℃条件下 WUE最高,而26 ℃和27 ℃之间差异不明显。在光合有效辐射高值区,B处理在3个温度梯度下WUE均先达到最大值,而后逐渐下降。其中,25 ℃处理WUE明显小于其他两个温度处理,而26 ℃处理WUE最高。比较两个水分处理春小麦WUE的大小,结果表明,在25 ℃条件下,随着PAR的增大,A处理WUE大于B处理;而26 ℃和27 ℃条件下,在PAR高值区,B处理叶片WUE较A处理增大。
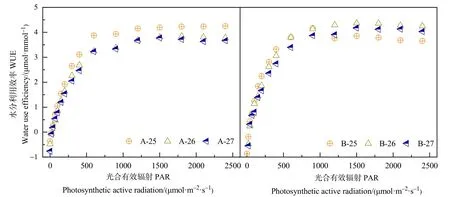
图3 不同处理春小麦开花期水分利用效率对光合有效辐射的响应Figure 3 Response of water use efficiency to photosynthetic active radiation of spring wheat at flowering stage under different treatments
3 结论与讨论
3.1 讨论
作物光合作用既受外界因子的影响,也受植物体内部结构和生理状况的调节(Silvia et al.,2004;张凯等,2021)。水分因子是作物生长发育的最大限制因素(郑盛华等,2006)。光照是光合作用的能源,在弱光下,光照是光合速率最重要的限制因子(喇燕菲等,2010;李伟等,2012)。温度通过影响作物的生化反应速率、调控蒸腾失水和气孔的开闭而影响光合生理过程(陈凤丽等,2013)。
本研究结果表明,水分供给不足,随着光合有效辐射增大,旗叶光合速率明显下降,最大光合速率减小,与王淑英等(2013)的研究结果一致。光饱和点与光补偿点是表征叶片对强光和弱光的利用能力大小的重要参数,代表了作物的需光特性和需光量(Joseph et al.,2005;叶子飘等,2010)。本研究结果表明,水分供给不足,春小麦光饱和点下降,光补偿点上升,光能利用能力下降,这可能与水分胁迫易导致光合产物运输受阻,在叶片中积累对光合作用形成一定的反馈抑制有关(王振夏等,2012)。水分供给不足,随着光照强度的增加,春小麦气孔导度在达到最大值后,又逐渐下降,胞间CO2浓度变化幅度较弱光时小,甚至有增加趋势,表明春小麦气孔限制因素向非气孔限制因素转变(Farquhar et al.,1982)。气孔导度逐渐下降,光合速率减小,蒸腾速率降低,这与大豆(郭数进等,2015)、马铃薯(王婷等,2010)等作物在土壤水分供给不足时的表现类似。降低幅度因作物品种的差异而不同。蒸腾速率的降低幅度较光合速率大,导致水分供给不足的处理的水分利用效率较供水充足处理大。
小麦属于C3作物,喜温凉气候,对高温胁迫的响应敏感(任鹏等,2010)。水分供给充足时,随着光照强度增加,较高的温度(27 ℃)由于提高了叶片气孔下腔的饱和水汽压,蒸腾作用增大,引起保卫细胞失水加剧,气孔收缩,气孔导度明显下降,胞间CO2浓度降低,气孔限制值Ls增大,CO2供应受阻,光合速率下降,与李娜等(2019)在增温对宁夏北部春小麦光合生理参数影响的研究结论一致。下降幅度因试验处理、气象生态因子以及春小麦品种的差异而不同。任鹏等(2010)研究表明,春小麦花后高温,旗叶净光合速率下降,与本研究结论一致。刘春溪等(2018)基于开放式主动增温系统(Free air temperature increasing,FATI),对水稻光合生理参数对增温的响应特征的研究中也得出类似结论,但降幅与本研究存在差异,主要是由于作物种类的不同而造成的。温度升高,植物光补偿点往往升高(Zhao et al.,2009)。本研究中,两种不同土壤水分条件下,温度升高,春小麦光补偿点升高,与已有研究结论一致(徐兴利等,2012)。表观量子效率和光补偿点的变化表明,较低温度下(25 ℃)春小麦利用弱光的能力增强。从净光合速率的最大值来看,26 ℃条件下春小麦最大光合速率较其他两个温度梯度大,而暗呼吸速率在 26 ℃温度时最大,表明 26 ℃条件下春小麦细胞活性更强。水分供给不足时,随着光强的增加,25 ℃条件下春小麦更易达到较高的光合速率;在光合有效辐射高值区,25 ℃条件下春小麦气孔导度明显大于其26 ℃和27 ℃。水分供给不足时,胞间CO2浓度和气孔限制值的波动表明,25 ℃条件下春小麦光合作用的变化受非气孔限制因素影响较大。从光合作用参数的模拟结果分析可知,温度越高,水分供给不足,春小麦利用强光和弱光的能力均下降,而最大净光合速率和暗呼吸也以 25 ℃最大,这表明在水分供给不足时,较低的环境温度更有利于春小麦光合作用,而过高的温度与水分胁迫叠加会造成光合过程受阻,从而可能影响生长发育以及产量的形成。
此外,从温度对不同水分条件下春小麦蒸腾作用的影响来看,随着光合有效辐射的增加,水分供给充足,温度升高利于蒸腾作用的进行。水分供给不足时,温度升高抑制了春小麦蒸腾作用,这与作物的自我保护相关。温度越高,在光合有效辐射高值区,气孔导度下降越快,气孔的部分关闭导致蒸腾速率不断减小。
3.2 结论
(1)水分条件对春小麦光合生理参数有重要影响。水分供给充足时,春小麦达到光饱和的光量子通量密度、表观量子效率增加,增幅分别为40%和28%;随着温度的升高,表观量子效率降低,最大净光合速率、光饱和点在水分供应充足时增大;光补偿点、暗呼吸速率在水分供应充足时较水分供应不足时下降;温度升高,叶片光补偿点增大。
(2)蒸腾速率在水分供应充足时较水分供给不足时明显增大。水分供给充足时,蒸腾速率随光合有效辐射的增加而增大,且对温度变化响应的敏感性下降。水分供给不足时,随着温度的升高,蒸腾速率的增加幅度下降;水分供给不足时叶片在各温度梯度下气孔导度随着光强的增大而下降,而光强增大,胞间CO2浓度下降,气孔限制值在水分供给充足时增大。
(3)在一定的光强范围内,水分利用效率随光强的增大而增大。水分供给充足时,春小麦水分利用效率在较低温度条件(25 ℃)下最大;水分供应不足时,在较低温度条件下(25 ℃),春小麦水分利用效率在光合有效辐射高值区快速下降。
猜你喜欢
今日农业(2021年3期)2021-12-05
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
农业与技术(2020年12期)2020-07-04
中国农业科技导报(2020年12期)2020-03-15
农民致富之友(2019年26期)2019-10-11
农学学报(2019年1期)2019-02-21
农民致富之友(2017年22期)2017-12-08
新课程·下旬(2017年7期)2017-08-14
湖北农业科学(2014年13期)2014-08-28
