
蒿柳1年生雌雄株幼苗的生长与防御差异研究
2022-08-09 13:43翟飞飞毛金梅李海栋张运兴李振坚刘俊祥孙振元
西北农林科技大学学报(自然科学版) 2022年8期
翟飞飞,毛金梅,李海栋,张运兴,李振坚,刘俊祥,李 霞,孙振元
(1 河南理工大学 建筑与艺术设计学院,河南 焦作 454000;2 中国林业科学研究院 林业研究所,北京 100091;3 新疆林业科学院 经济林研究所,新疆 乌鲁木齐 830063;4 菏泽学院 农业与生物工程学院,山东 菏泽 274000)
据研究,雌雄异株植物有15 600种,占被子植物(304 419~261 750种)的5.1%~6.0%,分布于175个科987个属[1]。有关雌雄异株植物的研究主要集中在雌雄比例[2-3]、雌株差异[4-5]、雌雄鉴定[6-7]、雌雄分化[8]以及雌雄决定[9-10]等方面。雌雄株在形态、生理生化等方面都表现出差异,可能与其对资源的需求和分配不同有关[11-13]。目前关于雌雄差异的研究较为广泛,且运用了形态学、生理学、组学等手段[14-17]。
植物吸收的资源通常有3条分配途径:生殖、生长和维持(新陈代谢或防御)[13,18]。生活史权衡理论认为,植物从外界获得的资源是有限的,某一途径资源投入得多,必然会减少其他途径的资源分配量,因此不同分配途径存在某种权衡关系[19]。一般而言,雌株需要孕育种子和果实,其生殖成本较高,而雄株的生殖成本较低[20-21],雌雄生殖成本的不同将导致其在生长、防御等方面存在差异。植物生长与防御之间的平衡是植物生理生态的基本原则[22]。Cornelissen等[23]对54项研究进行了汇总分析发现,大部分植物的雄株表现出生长快、防御水平低等特性,更易被食草动物取食。但对于柳属植物而言,不同种的雌雄株生长、防御能力存在很大差异:Salixlasiolepis、S.rigida等种的雌株生长慢、防御能力强[24-25];极地柳(S.polaris)等种的雌株生长快,但雌雄株的防御能力无异[26];还有一些种的雌雄株生长无差异,但防御能力表现不一[27-31];康定柳(S.paraplesia)雌株的生长和防御能力都强于雄株,可能是因为雌株存在超额补偿机制[15]。说明植物生长与防御具有很强的种特异性,需要对具体种进行针对性研究。前人针对生长、防御的研究主要集中在一个时间点上,但雌雄株间的差异在生长季不同阶段是否表现一致值得研究。
蒿柳为杨柳科柳属植物,具扦插易繁殖、生长速度快、轮伐期短等特性,在生物质能源开发、污染土壤的植物修复等方面应用前景广阔。本研究以蒿柳1年生扦插苗为观测对象,通过对雌雄幼苗生长季不同月份生长、防御等相关指标变化动态的分析,探讨雌雄株间的差异,以揭示蒿柳雌雄株的生长与防御关系,为雌雄株间的差异研究提供理论基础,并为其在生产实践中的选择性运用提供指导。
1 材料与方法
1.1 试验材料与设计
试验所用材料取自内蒙古自治区赤峰市赛罕乌拉自然保护区(44°14′ N,118°20′ E)。在野生群体中,采用成对取样的方法采集20对蒿柳雌雄株的插穗。采集时,每对雌株与雄株的间距控制在2 m以内,以保证为同一家系,从而消除年龄效应和位置效应;对与对之间至少相距50 m,以保证为不同家系。插穗采集后,在中国林业科学研究院科研温室外试验田进行扦插,并进行常规管理。随机选取10对蒿柳雌雄无性系于6月5日、7月3日、8月4日、9月7日和10月5日进行生长指标与光合作用参数测定;同时,取功能叶片于液氮中速冻,低温保存用于光合色素含量、酶活性、防御物质含量等指标测定。试验采用完全随机区组设计,共3个区组,每区组含10对蒿柳雌雄株,每对雌雄株各扦插1株。在指标测定时,区组间及区组内雌雄株均单独取样测定。
1.2 测定指标与方法
1.2.1 生长指标 植株地径采用游标卡尺测定基部以上1 cm处直径;植株高度采用卷尺测定植株基部到顶稍的长度。
1.2.2 光合作用参数 随机选取蒿柳雌雄幼苗的中上部叶片,于上午10:00―12:00用便携式光合测定仪Li-Cor 6400进行气体交换参数测定,参数包括净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr)。每株选2枚叶片,红蓝光源光强设为1 000 μmol/(m2·s)。叶室内叶片长度为3 cm,叶室两端及中间叶宽用直尺测定,叶片面积按2个梯形计算。将实际叶片面积输入Li-Cor 6400的输出文件,自动重新计算各参数。
1.2.3 光合色素含量 取蒿柳雌雄幼苗中上部叶片,去主脉后剪成0.3 cm×0.3 cm大小。称0.200 g叶片,用体积分数80%丙酮溶液黑暗浸提48 h后,测定663,645和470 nm处的吸光度,计算叶绿素a(chlorophyll a,Chl a)、叶绿素b(chlorophyll b,Chl b)和类胡萝卜素(carotenoid,Car)的含量。
1.2.4 多酚和单宁含量 多酚含量测定采用Folin-酚法,以邻苯二酚溶液作为标准品[32];缩合单宁含量测定采用香草醛-盐酸法,以儿茶素溶液作为标准品[32]。
1.2.5 酶活性 超氧化物歧化酶(superoxide dismutase,SOD)活性采用氮蓝四唑光还原法[33]测定;过氧化物酶(peroxidase,POD)活性采用愈创木酚法[32]测定;多酚氧化酶(polyphenol oxidase,PPO)活性采用分光光度法[32]测定;苯丙氨酸解氨酶(phenylalnine ammonia lyase,PAL)活性采用L-苯丙氨酸法[33]测定。
1.3 数据分析
采用SPSS 18.0软件和Excel 2003进行数据分析,数据均为3次重复的“平均值±标准误”,蒿柳雌雄株间的差异采用单因素方差分析,图表绘制在Origin 8.5中完成。
2 结果与分析
2.1 蒿柳1年生雌雄幼苗生长的差异
蒿柳1年生雌雄幼苗间株高和地径的差异见图1。

图柱上标不同小写字母表示雌雄株间有显著差异(P<0.05)。下同
由图1可以看出,在6-10月,蒿柳雌雄株的株高和地径均呈升高趋势。与前一测定时期相比,蒿柳雌雄株的株高增长率分别为59.84%,39.73%,17.27%,1.60%和92.92%,32.07%,24.49%,2.66%,10月雌雄株的株高生长减缓;雌雄株的地径增长率分别为36.43%,19.22%,17.27%,8.27%和37.08%,32.07%,18.08%,16.02%,10月雌雄株仍保持较高的径向生长速率。6月,蒿柳雌雄株间株高和地径均无显著差异(P>0.05);7月,雄株株高显著高于雌株(P<0.05),但雌雄株地径无显著差异(P>0.05);8-10月,蒿柳雄株株高、地径均显著高于雌株(P<0.05)。
2.2 蒿柳1年生雌雄幼苗光合作用的差异
由图2可知,6-10月蒿柳雌雄株的Pn和Gs呈先降低后升高趋势,7月最低;Ci变化较平缓;Tr在6-9月变化较平缓,10月较低。6月,蒿柳雌雄株间Pn无显著差异(P>0.05);7-10月,雄株Pn显著高于雌株(P<0.05)。6-9月,蒿柳雌雄株间Gs无显著差异(P>0.05);10月,雄株Gs显著高于雌株(P<0.05)。6-10月,蒿柳雌雄株间Ci均无显著差异(P>0.05)。6月,蒿柳雌雄株间Tr无显著差异(P>0.05);而7-10月,雄株Tr显著高于雌株(P<0.05)。

图2 蒿柳1年生雌雄幼苗间光合作用参数的差异
2.3 蒿柳1年生雌雄幼苗光合色素含量的差异
从图3可以看出,6-10月,蒿柳光合色素含量呈先降低后升高的趋势,7月最低。6月,蒿柳雌雄株间Chl a、Chl b和Car含量均无显著差异(P>0.05);7-10月,3种色素含量呈上升趋势,且均表现为雄株显著高于雌株(P<0.05)。

图3 蒿柳1年生雌雄幼苗间光合色素含量的差异
2.4 蒿柳1年生雌雄幼苗防御物质含量的差异
蒿柳雌雄株的防御物质主要是多酚和缩合单宁。从图4可以看出,6月蒿柳多酚和缩合单宁含量较高;7-10月,两者含量较低。在整个生长季(6-10月),蒿柳雌雄株间多酚和缩合单宁含量均无显著差异(P>0.05)。
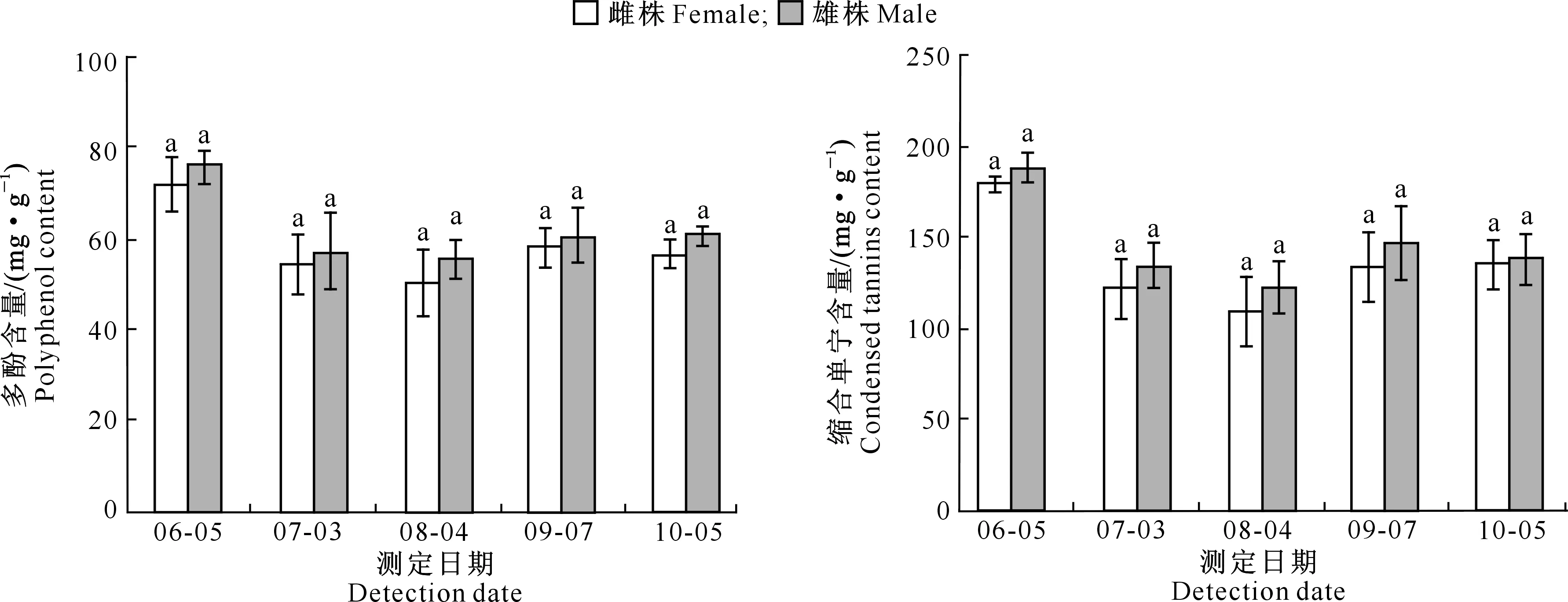
图4 蒿柳1年生雌雄幼苗间多酚和缩合单宁含量的差异
2.5 蒿柳1年生雌雄幼苗防御酶活性的差异
由图5可知,6-10月,蒿柳雌雄株SOD、POD、PPO和PAL活性变化趋势不尽相同,但4种酶活性均在10月时达到最高。除6月雌雄株间SOD和PAL活性无显著差异(P>0.05)外,其他各时期SOD、POD、PPO和PAL活性均表现为雄株显著高于雌株(P<0.05)。

图5 蒿柳1年生雌雄幼苗间防御酶活性的差异
3 讨 论
3.1 蒿柳1年生雌雄幼苗的生长差异
本试验过程中对供试材料都进行了正常养护管理,但试验正逢炎热夏季,蒿柳也遭受了一定程度的高温、干旱等逆境胁迫。本研究发现,蒿柳1年生雌雄幼苗生长、防御指标在不同月份差异较大,但是雌雄株间差异在7-9月较稳定,因此可选此时间段进行差异研究。
蒿柳雌雄株的株高在6月、地径在6-7月无显著差异,试验期间其他阶段的株高和地径均表现为雄株显著高于雌株,这与前人对东北红豆杉(Taxuscuspidata)[34]、S.rigida[25]、S.lasiolepis[24]等植物的研究结果一致。但极地柳[26]、东北鼠李(Rhamnusschneideri)[35]等植物雌雄株间生长差异不大,可能是由于雌株个体存在对自身生殖耗费的补偿机制[35]。
本研究中,6月蒿柳雌雄株间Pn无显著差异,7-10月雄株显著高于雌株。雄株的Pn高于雌株,表明雄株具有更强的光合作用,生长能力更强,与雌雄株的生长差异表现一致,在火炬树(Rhustyphina)等植物上的研究结果[36]也表现出相同规律。蒿柳雌雄株间Gs和Ci整体无显著差异,说明雌雄株的光合作用差异由非气孔限制因素引起。在光合作用过程中,光合色素起光能吸收、传递和转化的作用[37],其中叶绿素(Chl)是主要的光能吸收、转化色素,其含量高低直接反映植物的生长状况和光合能力;类胡萝卜素(Car)不仅是光合色素,还是一种重要的抗氧化剂,可吸收剩余能量、淬灭活性氧,防止膜脂过氧化[37-38]。6月蒿柳雌雄株间Chl a、Chl b和Car均无显著差异,7-10月3种光合色素含量均表现为雄株显著高于雌株,与光合作用变化趋势一致,说明光合色素含量是造成雌雄株光合差异的原因之一。
3.2 蒿柳1年生雌雄幼苗的化学防御差异
自然界中,植物经常遭受生物或非生物逆境的危害,并因此形成了机械防御、化学防御等多种防御机制[39]。其中,植物次生代谢物在化学防御中起主导作用,酚类、单宁等主要通过影响昆虫的取食和代谢而发挥防御作用[39-40]。水分胁迫下,滇杨(Populusyunnanensis)雌株的单宁、总酚、黄酮等防御物质含量显著低于雄株;而青杨(P.cathayana)的总酚含量无性别差异,雌株的缩合单宁含量高于雄株[41-42]。黑叶柳(S.myrsinifolia)和康定柳分别在增温和低土壤肥力条件下,雌株的单宁和酚类等物质含量高于雄株[15,30-31]。本研究结果显示,蒿柳雌雄幼苗的多酚和缩合单宁等防御物质含量均无显著差异,这与在极地柳[26]上的研究结果一致。

3.3 蒿柳1年生雌雄幼苗生长与防御的关系
根据资源可用性假说(resource availability hypothesis),内在生长速率决定防御水平,在恶劣环境下生长慢的植物用于防御的资源较多,而在适宜环境中生长快的植株防御投入较少[4,50]。如欧洲山杨(Populustremula)雌株生长较慢、生物量低,但雌株的黄酮、单宁等物质含量高,防御能力较强[4,51]。依据生活史权衡理论,一般雌株的生殖投资大于雄株,其用于营养生长或防御的资源较少,防御能力较差[35,52]。前人对滇杨1年生扦插苗在干旱条件下进行研究发现,雌株生长缓慢、防御能力较弱,表明繁殖影响了植物的生长与防御且在生活史中起优先作用,这可能是植物在繁殖与生长或防御间存在权衡,而不仅是生长与防御间的权衡[53]。本研究可以用生活史权衡理论来解释,即在有胁迫因子存在的自然环境中,蒿柳雌雄幼苗防御物质含量无显著差异,但雌株防御酶活性较低,生长相对缓慢,繁殖可能影响了雌株的生长与防御,蒿柳在繁殖与生长、防御间存在进化的权衡。前期在蒿柳天然分布种群的调研中发现,河岸带生境下性别比例偏雌性,而在远离河岸带的干旱生境中偏雄性。本研究中,蒿柳雌株生长缓慢,防御酶活性相对较低,干旱等胁迫因子可能降低了其适合度,可以为解释干旱生境中蒿柳偏雄性现象提供新视角。同时,逆境条件下雄株生长快且可解决飞絮问题,因此在实际应用中可优先选用。
4 结 论
对蒿柳1年生雌雄幼苗在6-10月的生长与防御指标进行动态分析发现,7-9月雌雄差异相对稳定,可选此阶段进行雌雄差异研究。整体来看,蒿柳雄株的株高和地径显著高于雌株,可能得益于雄株较高的净光合速率,而光合色素含量在蒿柳光合作用中发挥着重要作用。蒿柳雌雄株的多酚和单宁含量无显著差异,但雄株SOD、POD、PPO和PAL活性基本显著高于雌株,说明雄株在清除活性氧、抗病虫害方面的能力可能更强,但酶活性与防御能力的相关性有待进一步研究。本研究结果显示,在有胁迫因素存在的自然环境中,蒿柳雌雄幼苗防御物质含量无显著差异,但雌株防御酶活性较低,生长相对缓慢,繁殖可能影响了雌株的生长与防御,该物种在繁殖与生长、防御间存在进化的权衡。蒿柳雄株生长相对较快,可解决飞絮问题,在实际应用中可优先选用。
猜你喜欢
中外葡萄与葡萄酒(2022年4期)2022-07-26
动物营养学报(2022年5期)2022-06-10
酿酒科技(2022年5期)2022-05-24
今日农业(2021年10期)2021-11-27
今日农业(2021年9期)2021-11-26
诗潮(2021年1期)2021-03-15
儿童故事画报·自然探秘(2019年10期)2019-01-14
现代农业研究(2017年10期)2018-01-02
中老年健康(2016年1期)2016-03-07
Coco薇(2015年12期)2015-12-10
