
南亚热带滨海沙地3种林分类型凋落物的养分释放动态
2022-08-09 13:46曹雨虹张艺帆谢伟东
西北农林科技大学学报(自然科学版) 2022年8期
刘 易,曹雨虹,张艺帆,谢伟东
(广西大学 林学院,广西 南宁 530004)
凋落物是连接生态系统生物地球化学循环中植物-凋落物-土壤连续体的纽带,是维持土壤生物生存的物质和能量来源[1],凋落物腐解则是养分归还林地和地力恢复的主要途径[2]。同时,凋落物也是森林生态系统重要的碳库,在全球CO2变化过程中发挥复杂的源汇效应,凋落物分解对温室效应的贡献及全球变暖对凋落物分解动态的影响已成为新的研究热点[3]。植物吸收的养分中,90%以上的氮磷和60%以上的矿质元素都来自植物归还土壤的养分再循环[4]。因此,凋落物分解是生物地球化学循环过程的关键环节,在维持生态系统的结构和功能中占据着重要地位[5]。森林凋落物量、养分归还量及凋落物分解速率,主要受立地条件、林分类型、林龄、林分密度、凋落物组分和凋落物基质等多重因素影响[6-7]。目前有关森林凋落物的研究集中在不同森林生态系统和林分类型的年凋落物量及其组成、季节动态变化、养分分解过程、营养元素计量和凋落物动态影响因子等方面[8],但上述研究大多是针对单一树种凋落物进行的,而仅依据单一凋落物分解研究结果来反映自然界中混合凋落物的实际分解特征具有局限性。据统计,我国有关凋落物分解的研究主要集中在森林生态系统,而对湿地生态系统(包括沼泽、河流、湖泊、滨海等)凋落物分解过程的研究较少[5]。
滨海海陆交错过渡带生态系统具有典型的生态“边缘效应”和极强的生态脆弱性[9]。滨海地区土壤较为贫瘠,海岸沙地林分具有较高的养分转移率,林木较难直接从沙地获取养分,因此滨海沙地在植被恢复、地力固持、水土保持、水源涵养等方面的生物地球化学循环和生态服务功能受到广泛关注。由于滨海的经济开发造成生态系统退化、沙化严重,近年来我国滨海地区逐步引种木麻黄(Casuarinaequisetifolia)、巨尾桉(Eucalyptusurophylla×E.grandis)等树种对海防林进行更新改造[10],其中对木麻黄[11]、厚荚相思(Acaciacrassicarpa)[12]、吊丝竹(Dendrocalamusminor)[13]等人工纯林的养分转移机制也有相关研究。这些树种在海岸带抵御台风方面起到了较好的作用,但其存在群落结构单一、凋落物分解速度慢、水土固持和地力恢复效果欠佳等缺点。滨海地区以乡土树种为优势种的天然植被类型,具有更强的适应性和防风固沙能力,但我国对乡土树种的研究较晚,对其凋落物的动态变化研究也较少[14]。温远光等[15]研究发现,珍贵乡土树种红锥和望天树与桉树混交林的生物量比纯林提高13.97%~14.33%,而碳储量比纯林提高13.93%~14.89%。因此,继续深入开展海岸沙地乡土树种天然林养分吸收和转移的研究,对于揭示树种间的养分转移特性及滨海沙地的植被恢复具有重要意义[16]。
本试验对滨海沙地的高山榕(Ficusaltissima)/假苹婆(Sterculialanceolata)天然林、红鳞蒲桃(Syzygiumhancei)天然林、马占相思(Acaciamangium)人工林纯林的凋落物养分含量和养分归还量变化动态进行研究,剖析滨海特殊生境下天然林和人工林的养分循环过程及维持地力的能力,以期为滨海天然林保护、植被恢复和防护林建设树种的选择与配置提供理论依据。
1 材料与方法
1.1 研究地概况
研究地位于广西防城港东兴市北仑河口红树林保护区海岸带(107°53′~108°15′E,21°31′~21°44′N)。东兴市地处广西壮族自治区南部,属于南亚热带季风性海洋气候,全年气候温和,雨热同期,春夏多雨,盛行台风,秋冬干旱;年平均气温23.2 ℃,年日照时数在1 500 h以上,年平均降雨量达2 822.9 mm。该区域为滨海平原,地势平坦,海拔一般在10 m以下。土壤类型为砂质土,土壤易沙化,水分、养分流失严重。
高山榕+假苹婆天然次生林(记为高山榕/假苹婆,下同)乔木层平均高16.3 m,平均胸径22.7 cm,郁闭度0.8,植被总盖度92%,主要乔木树种有高山榕、假苹婆、红鳞蒲桃、鸭脚木(Scheffleraoctophylla)、肉实树(Sarcospermalaurinum)、密花树(Rapaneaneriifolia)等;灌木层盖度95%,主要种类有细枝龙血树(Dracaenagracilis)、罗伞树(Ar-disiaquinquegona)、南山花(Prismatomerisconnata)、九节(P.rubra)、龙船花(Ixorachinensis)等;草本层盖度10%,主要种类有山姜(Alpiniachinensis)、沿阶草(Ophiopogonbodinieri)、鸭跖草(Commelinacommunis)等。
红鳞蒲桃天然林(记为红鳞蒲桃,下同)乔木层平均高15.6 m,平均胸径26.2 cm,郁闭度0.7,植被总盖度86%,主要乔木树种有红鳞蒲桃、黑嘴蒲桃(S.bullockii)、岭南山竹子(Garciniaoblongifolia)、鸭脚木等;灌木层盖度95%,主要种类有山油柑(Acronychiapedunculata)、鸭公树(Neolitseachui)、九节、龙船花、鸭脚木等;草本层盖度10%,主要种类有沿阶草和露兜树(Pandanusaustrosinensis)等。
马占相思人工林(记为马占相思,下同)乔木层平均高13.7 m,平均胸径15.9 cm,郁闭度0.5,植被总盖度65%,主要乔木树种是马占相思(A.mangium);灌木层盖度75%,主要种类有乌药(Linderaaggregata)、桃金娘(Rhodomyrtustomentosa)、岗松(Baeckeafrutescens)、展毛野牡丹(Melastomanormale)、潺槁木姜子(Litseaglutinosa)等;草本层盖度30%,主要种类有荩草(Arthraxonhispidus)、毛果珍珠茅(Scleriaherbecarpa)等。
1.2 试验方法
1.2.1 凋落物样品采集 于2018年2月用收集网法采集凋落物。在3种不同林分类型内植物分布均匀处分别设置20 m×30 m的样方,采用对角线五点法各安装5个1 m×1 m的孔径0.833 mm(120目)的凋落物尼龙收集网,用PVC塑料管支撑使其最低点与地面隔离50 cm左右,为防止台风影响在收集网内放一石块。自2018年3月至2019年2月,每月月底固定时间收集凋落物。将收集到的凋落物按照枝、叶、花果以及杂物(树皮、碎屑、动物和昆虫粪便及残体等)进行分组,并将同一林分的相同组分合并为一个样品。
1.2.2 凋落物分解试验 凋落物分解试验用野外分解袋法。在每个林分类型内随机设置9个1 m×1 m的样方,于2018年1月收集新鲜的凋落物去杂,按照枝、叶、花果以及杂物(树皮、碎屑、动物和昆虫粪便及残体等)进行分组后自然风干,一份样品用于测定各组分初始化学养分含量;另取一份风干样品(20 g)装入规格为15 cm×15 cm、孔径1 mm的尼龙分解袋内。根据前期试验测定的凋落物各组分的干质量,确定叶、枝、花果、杂物放入分解袋的质量比为16∶2∶1∶1,于当月安装凋落物收集网的同时将分解袋放回对应的林分内。每个林分内按10 m间距放置3列分解袋作为3个处理,每列再按5 m间距放置12个分解袋,共36个分解袋。放置前先去除地表凋落物,将分解袋平放地上再按原状覆盖凋落物。在每个月收集凋落物的同时,从每种林分类型3列处理中各取回1个凋落物分解袋(3个重复)。
1.2.3 凋落物养分含量测定 凋落物中全C含量采用浓硫酸-重铬酸钾法测定,全N含量采用凯氏定氮法测定,全P含量采用钼锑抗比色法测定,全 K含量采用火焰光度计法测定, Ca、Mg含量采用原子吸收分光光度计测定[17]。
1.3 研究内容
1.3.1 不同林分类型年凋落物量及各养分平均含量的比较 将每月收集的各林分类型的凋落物样品,于80 ℃烘箱中烘干至恒质量,称其总干质量(精确到0.01 g),然后按收集网面积换算成每公顷凋落物量(kg/hm2),即为月凋落物量,12个月凋落物量之和则为年凋落物量。将测定的每个月凋落物中C、N、P、K、Ca、Mg等养分含量进行平均,计算不同林分各养分的平均含量及其变异系数。

(1)
1.3.2 不同林分类型凋落物各养分的月动态变化 绘制3种林分类型凋落物C、N、P、K、Ca、Mg含量的月动态变化曲线,比较不同林分凋落物各养分含量的月变化规律。
1.3.3 不同林分类型凋落物各养分年归还量特征 清除每月取回的凋落物分解袋样品中的土壤、树根和其他杂物,在80 ℃烘箱中烘干至恒质量,测定各月凋落物分解后的残留质量及C、N、P、K、Ca、Mg等养分残留量,再利用凋落物收集网中各组分月凋落量平均值与月凋落物量的平均值之比、凋落物中各组分每种养分的初始含量与所有组分同一养分初始含量之和的比值,计算凋落物养分年归还量和各组分养分年归还量。凋落物各养分年归还量是指12个月内植物以凋落物形式向单位面积(m2或hm2)环境中归还的凋落物各组分的同一养分之和,计算公式如下:
(2)
(3)
(4)
式中:X表示凋落物各养分年归还量(kg/hm2);Xi表示凋落物各组分同一养分年归还量(kg/hm2);i表示各组分,i=1~4;j表示各养分元素,j=1~6;n表示月份,n=1~12;Cin表示凋落物某一组分在n月养分含量(g/kg);Cjn表示凋落物分解残留物中n月某一养分含量(g/kg);Cij0表示凋落物某一组分中某一养分的初始含量(g/kg);Ci0表示凋落物中某一养分总初始含量(g/kg);Mn表示第n月凋落物的残留物量(g/m2);M(n+1)表示第n+1月凋落物的残留物量(g/m2);Ci(n+1)表示凋落物某一组分在n+1月的养分含量(g/kg)。
1.3.4 不同林分类型凋落物各组分同一养分年归还量特征 将不同林分叶、枝、花果、杂物各组分每个月的C、N、P、K、Ca、Mg归还量相加,得到其各养分年归还量(Xi,见公式(3)),比较不同林分类型各组分同一养分年归还量的差异。
1.3.5 不同林分类型凋落物各养分归还量的月动态变化 绘制3种林分凋落物C、N、P、K、Ca、Mg归还量的月动态变化曲线,比较不同林分凋落物各养分归还量的月变化规律。
1.3.6 不同林分类型凋落物养分含量及其C、N、P化学计量比与凋落物养分归还量的相关性 通过3种林分凋落物的 C、N、P 含量计算C/N、C/P和N/P,对各林分C、N、P、K、Ca、Mg含量,C/N、C/P、N/P以及C、N、P归还量进行相关性分析。
1.4 数据处理方法
采用统计软件SPSS 26.0对凋落物养分含量和养分归还量进行单因素方差分析和多重比较分析,采用R软件ggplot 2包对凋落物养分含量及其C、N、P化学计量比和养分归还量进行相关性分析并绘图。
2 结果与分析
2.1 不同林分类型年凋落物量及各养分平均含量的比较
表1结果表明,3种林分年凋落物量表现为高山榕/假苹婆>红鳞蒲桃>马占相思,且高山榕/假苹婆与红鳞蒲桃间不存在显著差异(P>0.05),但二者均显著高于马占相思(P<0.05)。3种林分类型凋落物各养分含量大小均表现为C>Ca>N>K>Mg>P,变异系数在5.24%~71.78%,其中Mg含量的变异系数相对较大(54.39%~66.93%),C含量的变异系数最小(5.24%~6.69%);红鳞蒲桃K含量的变异系数(71.78%)明显大于高山榕/假苹婆(32.67%)和马占相思(27.18%)。在3种林分类型凋落物的养分中,C含量差异不显著(P>0.05);N含量存在显著差异(P<0.05 ),表现为高山榕/假苹婆>红鳞蒲桃>马占相思;马占相思与红鳞蒲桃和高山榕/假苹婆间P含量存在显著差异(P<0.05),表现为高山榕/假苹婆>红鳞蒲桃>马占相思;K含量在3种林分类型间均存在显著差异(P<0.05),表现为红鳞蒲桃>高山榕/假苹婆>马占相思;马占相思的Ca含量最高,红鳞蒲桃的Ca含量最低且与高山榕/假苹婆、马占相思间存在显著差异(P<0.05);3种林分类型Mg含量均较低且不存在显著差异(P>0.05)。

表1 滨海沙地不同林分类型的年凋落物量及各养分平均含量
2.2 不同林分类型凋落物各养分的月动态变化
3种森林类型凋落物各养分含量的月动态变化呈双峰型、三峰型或多峰型(图1)。由图1-A可以看出,各林分类型凋落物中C含量变化相对稳定,呈多峰型变化趋势,其中高山榕/假苹婆凋落物C含量为456.46~550.52 g/kg,在4月和12月各出现一个较大的峰值,其中12月为最大峰值;红鳞蒲桃凋落物C含量在2月达到最大(490.6 g/kg);马占相思凋落物C含量在4月出现最大峰值(524.57 g/kg)。由图1-B可见,高山榕/假苹婆和马占相思凋落物的N含量变化模式均呈双峰型,红鳞蒲桃凋落物的N含量变化模式呈多峰型,三者的最大峰值均出现在11月,分别为23.15,11.72和15.24 g/kg,第2峰出现在4月。图1-C显示,各林分类型凋落物P含量在12个月内波动变化较大,高山榕/假苹婆的P含量从7月至次年2月呈持续下降趋势,呈双峰型变化,变化幅度为0.21~1.55 g/kg;红鳞蒲桃和马占相思凋落物P含量呈三峰型变化趋势,其中红鳞蒲桃的最大值和最小值分别出现在4月和7月,变化幅度为0.32~1.78 g/kg,而马占相思的P含量最大值和最小值分别出现在12月和2月,变化幅度为0.45~1.15 g/kg。红鳞蒲桃凋落物K含量变幅最大,总体上呈递减的趋势,分别在3月(17.56 g/kg)和11月(8.86 g/kg)出现较大的峰值;高山榕/假苹婆和马占相思在1年中凋落物K含量较低且变化平稳,变化范围分别为1.38~5.44 g/kg和1.23~3.54 g/kg(图1-D)。由图1-E可知,高山榕/假苹婆、红鳞蒲桃及马占相思凋落物的Ca含量变化规律基本一致,呈先升后降的变化趋势,最高值分别出现在9月、8月和7月,分别为49.00,33.38和39.03 g/kg,其中高山榕/假苹婆的Ca含量变化幅度较大。3种林分类型的凋落物Mg含量变化趋势也基本一致,3-9月处于较高水平,而后呈快速下降趋势,12月至次年2月达到最低值(图1-F)。
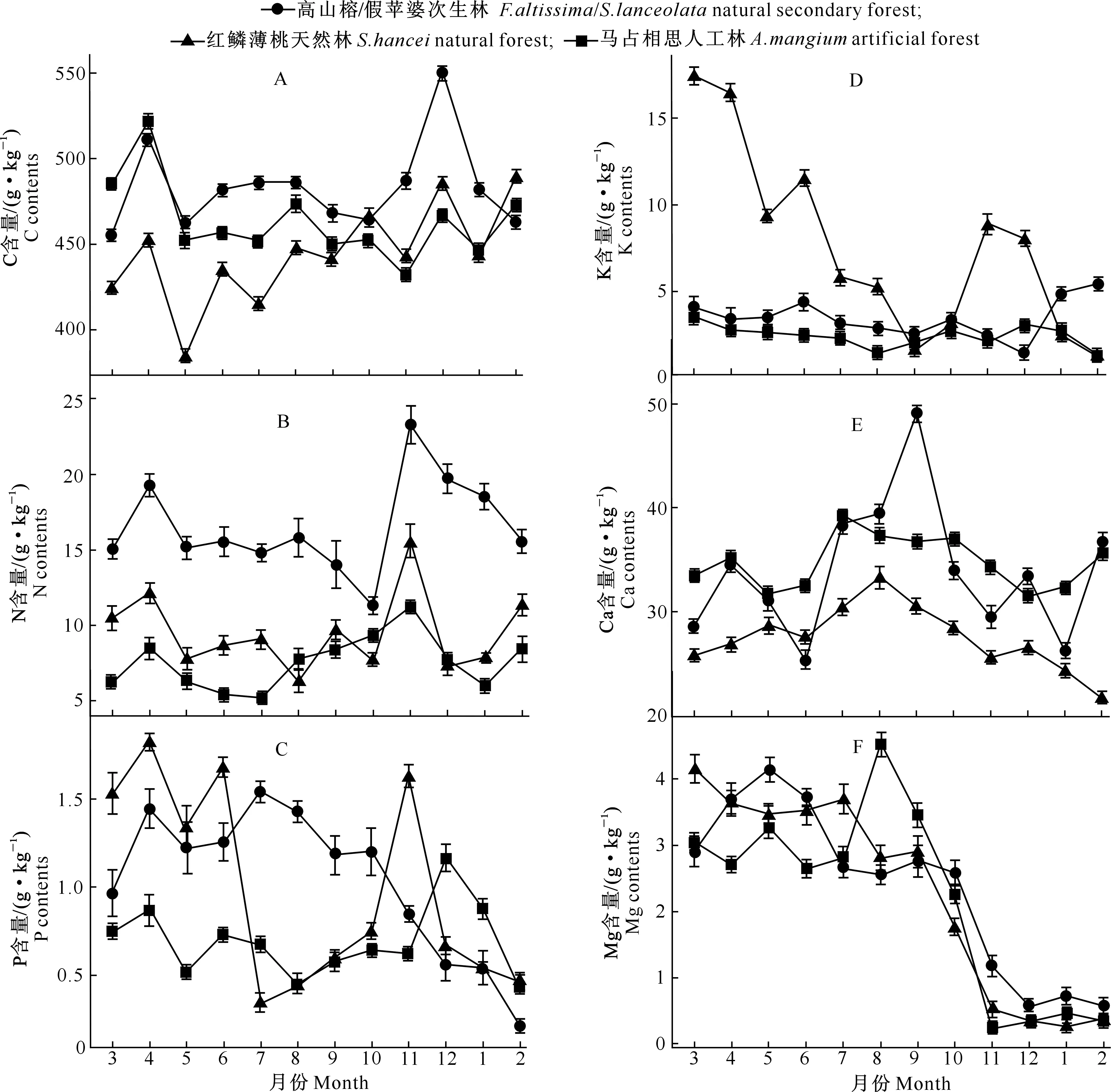
图1 滨海沙地不同林分类型凋落物各养分含量的月动态变化
2.3 不同林分类型凋落物各养分的年归还量特征
由表2可知,凋落物各养分年归还量与其养分含量大小排序相同。3个林分类型的N、K、Mg归还量间均存在显著差异(P<0.05),表现为高山榕/假苹婆>红鳞蒲桃>马占相思;马占相思凋落物的C、P、Ca归还量显著小于红鳞蒲桃和高山榕/假苹婆(P<0.05),但红鳞蒲桃与高山榕/假苹婆间均不存在显著差异(P>0.05) 。

表2 滨海沙地不同林分类型凋落物各养分年归还量特征
2.4 不同林分类型凋落物各组分同一养分的年归还量特征
3种林分类型叶、枝、花果、杂物的N、K、Mg,叶、枝、杂物的Ca以及枝、花果和杂物C的年归还量间均存在显著差异(P<0.05) (图2);高山榕/假苹婆和红鳞蒲桃叶及杂物P的年归还量与马占相思存在显著差异(P<0.05);高山榕/假苹婆和红鳞蒲桃各组分养分年归还量顺序均表现为叶>枝>杂物>花果,而马占相思凋落物各组分养分年归还量顺序均为叶>花果>枝>杂物,其中叶的养分归还量比重占绝对优势,其他组分的养分归还量均极低;叶的C、N、K、Mg归还量表现为高山榕/假苹婆>红鳞蒲桃>马占相思,P和Ca归还量则表现为红鳞蒲桃>高山榕/假苹婆>马占相思。

图2 滨海沙地不同林分类型凋落物各组分同一养分的年归还量特征
2.5 不同林分类型凋落物各养分归还量的月动态变化
由图3可以看出,2种天然林各养分归还量的月变化趋势基本相似,呈双峰型或三峰型。高山榕/假苹婆除P外(6月),C、N、K、Ca、Mg的归还量第1峰出现在5月,归还量分别为1 802.55,54.14,14.13,106.45和10.88 kg/hm2,第2峰均出现在8月,随后逐渐递减;红鳞蒲桃各养分归还量第1峰均出现在5月,C、N、P、K、Ca、Mg归还量分别为1 593.28,47.70,5.05,10.26,124.75和8.80 kg/hm2,第2峰出现在8月;马占相思各养分归还量均处于较低水平,呈双峰型,除K外(11月),C、N、P、Ca、Mg归还量最高峰均出现在5月,分别为651.59,31.40,1.14,39.96和1.63 kg/hm2,第2峰均出现在11月。

图3 滨海沙地不同林分类型凋落物各养分归还量的月动态变化
2.6 不同林分类型凋落物养分含量及其C、N、P化学计量比与凋落物养分归还量的相关性
由表3~表5可知,不同林分类型中凋落物养分含量及化学计量特征与养分归还量在一定程度呈显著(P<0.05)或极显著(P<0.01)相关。

表3 高山榕/假苹婆天然次生林凋落物养分含量及其C、N、P化学计量比和养分归还量间的相关性
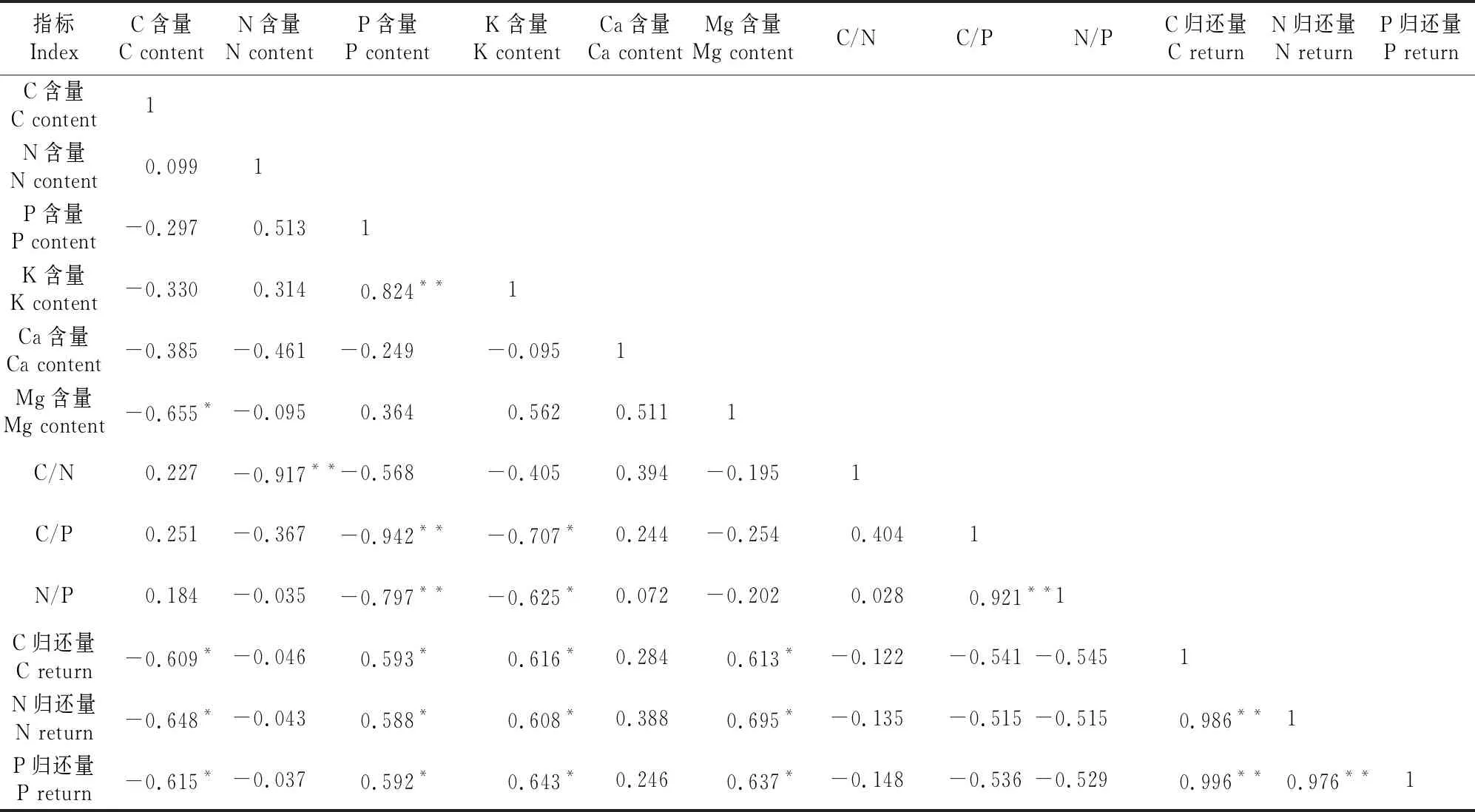
表4 红磷蒲桃天然林凋落物养分含量及其C、N、P化学计量比和养分归还量间的相关性

表5 马占相思人工林凋落物养分含量及其C、N、P化学计量比和养分归还量间的相关性
高山榕/假苹婆凋落物C含量与K含量呈显著负相关,N含量与C/N呈极显著负相关,P含量与Mg含量、N归还量、P归还量均呈极显著正相关。C/P与P含量、Mg含量呈极显著负相关,C/P与N/P呈极显著正相关,N/P与P含量、Mg含量呈极显著负相关。C归还量与P含量、Mg含量呈显著正相关关系,N归还量和P归还量均与C/P、N/P呈显著负相关,N归还量和P归还量均与Mg含量呈极显著正相关。
红鳞蒲桃凋落物C含量与Mg含量及C、N、P归还量均呈显著负相关,N含量与C/N呈极显著负相关,P含量与K含量呈极显著正相关,P含量与C/P、N/P均呈极显著负相关。C/P与N/P呈极显著正相关,C/P和N/P均与K含量呈显著负相关。C、N、P归还量均与P、K、Mg含量存在显著正相关关系。
马占相思凋落物P含量与K含量呈显著正相关,C/N与N含量、N/P呈极显著负相关,C/P和N/P均与P含量、K 含量极显著负相关,N/P与N含量、C/P存在显著正相关关系。
3 讨 论
3.1 不同林分类型凋落物养分含量及动态特征
凋落物养分含量的高低可以反映植物对养分的利用效率,养分通常通过转移机制得到再利用。在同一气候条件下,林分类型是造成凋落物养分含量差异的主要因素,其他诸如林龄、林分密度、植被生产力状况、凋落物元素初始含量等亦是影响凋落物养分含量的重要因素[18-19]。本研究中3种林分类型凋落物养分含量大小均为C>Ca>N>K>Mg>P,其中N、P、K、Ca的养分含量在林分类型间存在不同程度的显著差异(P<0.05),这与齐泽民等[20]对川西亚高山林线交错带植被养分含量的研究结果一致,而与福建沿海纹荚相思人工林(N>K>Ca>Mg>P)[21]、广东鼎湖山常绿阔叶林(N>K>Ca>Mg>P)[22]、湖南亚热带典型的针阔混交林(C>N>Ca>K>S>Mg>P)[23]有所不同,但均存在P含量较低的特点,可能与树种组成和立地条件有关。
3种林分类型凋落物的C含量大小表现为高山榕/假苹婆>马占相思>红鳞蒲桃,均高于四川官司河流域5种植被类型[24]、福建滨海沙地尾巨桉和木麻黄人工林[25],表明本区域高温、多雨,林木生长周期长,具有较高的生产力。
3种林分类型凋落物的N含量均低于Han等[26]报道的全国753种陆地植物的N平均含量(18.6 g/kg),红鳞蒲桃和马占相思凋落物N含量均低于福建滨海沙地肯氏相思凋落物(15.49 g/kg)[27]。滨海沙地林分凋落物较低的N含量可能与滨海沙地养分涵养能力较差和较强的养分重吸收有关,当植物体内和生境养分含量较低时,在叶片脱落之前部分养分会转移到植物体内其他部位,以维持植物体的正常生长发育[26];另外该区域年降雨量大,也会对凋落物产生高强度的淋溶作用[28]。本研究区夏季雨热同期,林木生长旺盛,N的消耗量和降雨淋溶作用都较大,在5-10月凋落物N含量均处于较低水平,但在11 月出现N元素的快速积累,并达到最大峰值,说明凋落物N含量与植物生物学特性存在密切的关系,可能这一时期是养分再吸收的主要节点,这有待下一步深入研究。
3种林分类型凋落物的P平均含量均低于全球范围内森林生态系统的P平均含量(1.77 g/kg)[29],也低于我国753种陆地植物的P平均含量(1.21 g/kg)[26],但高于福建滨海沙地尾巨桉和木麻黄人工纯林的凋落物P含量[25]。植物在较贫瘠生境和衰老过程中P元素存在较高的转移率,养分再吸收明显[30],本研究结论支持这一观点。3种林分类型凋落物的P含量月变化表现为双峰型和三峰型,变化幅度和规律差异较大,高山榕/假苹婆的P含量变化规律不同于另外2种林分,7月后一直呈递减趋势,未表现出因养分的再吸收而再次形成峰值,这是因为在凋落物分解前期P还不能被微生物利用,所以发生暂时积累,但随着分解的进行,凋落物内部结构发生变化,在微生物作用下被矿化,因此P含量表现出下降趋势。本研究中2种天然林凋落物的P含量和变异系数均显著高于马占相思人工林,反映了同一区域中林分类型是凋落物P含量的主要影响因素之一。相关研究表明,滨海沙地土壤P含量偏低,林业发展受到P限制[27,31],表明在滨海沙地利用乡土树种营造近自然的混交林或复层林,更有利于地力恢复。
K在植物体内既不是有机质的组分, 也不是代谢的中间产物,主要以离子状态存在,因此再转运能力强[32]。植株快速生长阶段需要吸收大量N、K合成蛋白质,以增强光合作用[33]及提高对恶劣环境的抗性[34]。高山榕/假苹婆和马占相思凋落物全年K含量均处于较低水平且较平稳,而红鳞蒲桃凋落物K含量呈夏季快速下降冬季回升的趋势。高山榕/假苹婆(3.45 g/kg)和红鳞蒲桃(7.60 g/kg)凋落物K平均含量均高于神农架巴山冷杉天然林凋落物K含量(2.92 g/kg)[35]和福建沿海沙地吊丝竹林凋落物K含量(2.84 g/kg)[36],马占相思凋落物K含量(2.43 g/kg)与其相近。马占相思凋落物K含量较低,可能与其林龄有关,植株在生长较快阶段细胞代谢增强,需要合成大量的蛋白质和核酸[33],因此叶片在脱落前将养分转移到植株其他器官,造成凋落物中K含量较低。
3种林分的凋落物Ca含量均在7-10月间出现最大峰值。Ca是移动性相对较低的元素,在成熟叶片中积累较多,导致凋落叶中Ca含量较高[20,37]。而Mg是可移动的生物元素,是构成叶绿素的主要矿质元素,3种林分Mg含量在3-9月的生长季节维持较高水平,随后急剧下降。
3.2 不同林分类型凋落物养分归还量及动态特征
3种林分类型凋落物的各养分年归还量大小排序均为C>Ca>N>K>Mg>P,与川西亚高山林线交错带群落凋落物养分归还量结果一致[20],与福建沿海纹荚相思人工林(N>K>Ca>Mg>P)[21]、亚热带7种常绿阔叶林优势树种落叶[38]的养分年归还量结果存在一定差异。各养分元素归还量受凋落物养分含量的影响,均在5月出现最大峰值,随后呈递减的趋势,在8月出现第2峰,这与当地的水热条件和林木生物学特性的季节变化同步。由于叶片是森林地上凋落物的主要组成部分,凋落物的养分归还量及变化规律与凋落叶量变化一致。3种林分类型养分年归还总量表现为高山榕/假苹婆>红鳞蒲桃>马占相思,其中,三者N、K、Mg归还量间均存在显著差异,马占相思凋落物C、P、Ca归还量显著低于红鳞蒲桃和高山榕/假苹婆,而红鳞蒲桃与高山榕/假苹婆间不存在显著差异,表现出天然林在养分循环中存在较大的优势。
3种林分凋落物各组分养分年归还量平均值大小均表现为C>Ca>N>K>Mg>P。就各组分而言,高山榕/假苹婆和红鳞蒲桃均表现为叶>枝>杂物>花果,马占相思表现为叶>花果>枝>杂物,这与李洁冰等[39]对湖南樟树林凋落物的研究结果相似。本研究结果表明,凋落叶养分归还量占有较大的比例,与川西亚高山林线交错带养分归还特征相似[20],经由落叶的养分归还是本研究中养分归还的主要形式。从马占相思凋落物各组分的养分归还量来看,凋落枝中养分归还量明显低于叶、繁殖器官等组分。与枝和繁殖器官相比,叶中各养分归还量最高,原因在于叶是植物生命活动中最活跃的器官,是植物进行光合作用的主要场所,其中C是叶片合成有机物质的主要原料,在维持植物生长和代谢方面具有重要作用。
3.3 凋落物养分含量及化学计量比与养分释放的关系
植物C∶N∶P值反映了生态系统碳循环、氮磷元素相互作用与制约的变化规律[40]。凋落物质量和C/N被认为是影响凋落物分解和养分归还的主要因素之一,C/N 越低,凋落物分解速率越快[41]。红鳞蒲桃和马占相思的凋落物C/N分别为49.43和64.84,与中国4种不同林型的平均值44.76和6种生态系统的平均值52.9相近[42],而高山榕/假苹婆凋落物C/N(30.15)则偏低,但3种林分均高于典型喀斯特峰丛洼地常绿落叶阔叶林(22)和顶极群落(18)的C/N[43],表明本研究中2种天然次生林凋落物的分解速率相对较慢,更有利于滨海沙地的养分存储和水源涵养。研究表明,N/P值越高,凋落物的分解受P的限制越强,特别是N/P值大于25和P含量低于0.22 g/kg的状况下[43]。本研究中高山榕/假苹婆、红鳞蒲桃和马占相思的凋落物N/P分别为22.31,12.67和11.82,红鳞蒲桃和马占相思凋落物N/P低于全国36个地点的平均值(21.35)和全球范围内的平均值(18.3)及25的阈值,P平均含量分别为1.03,0.99和0.69 g/kg,表明凋落物的分解受P的限制较小。
对于不同的林分类型,凋落物的P、K、Mg与C/P和N/P之间存在不同程度的负相关关系。研究表明,P添加可提高叶凋落物分解速率30%,而微量元素(如B、Ca、Cu、Fe、Mg、Mn、Mo、S、Zn)的添加则可提高81%,表明除P之外这些微量元素参与并促进凋落物的分解[5],本研究支持这一结果,认为凋落物的分解与养分释放受多种元素的共同影响。然而,不同的林分类型、结构、密度和立地条件,以及凋落物养分含量和化学计量比对养分归还量的影响也存在较大差异,高山榕/假苹婆凋落物P、Mg含量与C、N、P的归还量呈显著或极显著正相关,C/P、N/P与N、P的归还量呈显著负相关;红鳞蒲桃的凋落物C含量与C、N、P的归还量呈显著负相关,P、K、Mg含量均与C、N、P归还量呈显著正相关;而各养分元素含量及化学计量比对马占相思的C、N、P归还量无显著影响。凋落物的分解及养分释放除受各养分元素含量及化学计量比的影响外,还受气温、湿度、微生物种群结构和数量及凋落物基质等因素的影响,因此对不同森林生态系统还需结合群落类型、环境因子、凋落物和微生物特性等进行研究,以进一步探明生态系统的养分循环机制。
4 结 论
3种林分类型的年凋落物量和总养分归还量表现为高山榕/假苹婆>红鳞蒲桃>马占相思,凋落物的各养分含量和各养分归还量均表现为C>Ca>N>K>Mg>P;与其他生态系统相比,凋落物各养分含量和归还量相对较低。凋落物养分含量和归还量的月变化曲线以三峰型为主,其次是双峰型和多峰型,峰值主要出现在5、8、11月。在各组分中,凋落叶的养分归还量比重占绝对优势。研究区不同林分类型凋落物分解及养分释放受各养分元素含量及化学计量比影响差异较大,其中凋落物分解受P的限制较小。相对于马占相思人工纯林,高山榕/假苹婆和红鳞蒲桃2种天然次生林凋落物的分解速率相对较慢,更有利于滨海沙地养分存储和水源涵养。因此,在南亚热带滨海地区根据乡土树种特性采用近自然混交模式进行植被恢复和防护林建设,同时结合科学合理的森林经营方式,更有利于改善滨海沙地生态系统和土壤质量,提高土壤的碳汇能力。
猜你喜欢
纺织标准与质量(2022年2期)2022-07-12
中国土壤与肥料(2022年4期)2022-06-14
当代水产(2022年4期)2022-06-05
煤气与热力(2021年12期)2022-01-19
四川蚕业(2021年2期)2021-03-09
落叶果树(2021年6期)2021-02-12
天津医科大学学报(2021年1期)2021-01-26
文苑(2020年12期)2020-04-13
当代化工(2019年3期)2019-12-12
海峡姐妹(2019年1期)2019-03-23
