
2株感染蠼螋的球孢白僵菌的分离鉴定及对草地贪夜蛾的毒力
2022-08-08 03:56温绍海庞继鑫杜广祖张立敏
植物保护 2022年4期
温绍海, 庞继鑫, 杜广祖, 陈 斌, 张立敏
(云南农业大学植物保护学院, 云南生物资源保护与利用国家重点实验室, 昆明 650201)
蠼螋属于有翅亚纲Pterygota革翅目Dermaptera,也称夹板虫、剪刀虫等[1]。自然感染蠼螋的昆虫病原真菌种类较少,已报道有球孢白僵菌Beauveriabassiana感染欧洲球螋Forficulaauricularia和松球螋Forficulasp.[2],神农架被毛孢Hirsutellashennongjiaensis感染未知种蠼螋[3],因而对蠼螋上球孢白僵菌菌株及其毒力的研究报道较为少见。
草地贪夜蛾Spodopterafrugiperda(J. E. Smith)是一种严重为害玉米等粮食作物的跨境迁飞性害虫,寄主植物范围广,繁殖力强,迁飞扩散快,为害程度重,防控难度大[4]。草地贪夜蛾自2019年1月入侵我国并迅速定殖、蔓延,严重威胁我国玉米生产安全[5]。昆虫病原真菌对草地贪夜蛾具有良好的致病作用,如莱氏绿僵菌Metarhiziumrileyi[6]、玫烟色拟青霉Paecilomycesfumosoroseus[7]、金龟子绿僵菌M.anisopliae和球孢白僵菌Beauveriabassiana[8-9]。然而,不同种类昆虫病原真菌对草地贪夜蛾的侵染致病作用不同,且在田间自然感染草地贪夜蛾的昆虫病原真菌还不多见,仅郑亚强等和雷妍圆等分别从云南和广东玉米田发现并分离鉴定了感染草地贪夜蛾幼虫的莱氏绿僵菌[10-11],程东美等在广州玉米田发现草地贪夜蛾僵虫并分离获得了球孢白僵菌[12]。因此,开展昆虫病原真菌对草地贪夜蛾的毒力测定,筛选高致病力菌株十分必要。
本研究于2020年8月从云南省红河州元阳县大果公路和保山市隆阳区蒲缥镇黄泥村玉米田采集到自然染病的蠼螋,根据虫体形态特征和地理分布初步鉴定采集自元阳县的蠼螋为宽翅球螋PterygidaeurypygaBey-Bienko,采集自隆阳区的蠼螋为华球螋ForficulasinicaBey-Bienko。通过形态与分子鉴定病原物为球孢白僵菌,并测定了其基本生物学特性和对草地贪夜蛾3龄幼虫的毒力,旨在为草地贪夜蛾生物防治提供菌种资源,同时也可为该两株球孢白僵菌的生防潜力研究奠定基础。
1 材料与方法
1.1 菌株的分离培养
将采集到的蠼螋僵虫,带回室内用无菌水保湿培养,待产孢后挑取虫体上的孢子,接于SDAY培养基[13],于温度(25±1)℃,光周期L∥D=12 h∥12 h,相对湿度(75±5)%的光照培养箱中培养。培养7 d后,将分生孢子转接到新的培养基上进行纯化。经过反复纯化,得到纯的菌株,纯化的菌株采用SDAY 斜面培养基保存于4℃冰箱备用。
1.2 菌株生物学特性
将保存的菌株作为备用菌株,以SDAY培养基作为供试培养基,设置22℃、25℃和28℃ 3种温度条件(误差±1℃),RH(75±5)%,光周期L∥D=12 h∥12h条件下的不同光照培养箱中培养。菌落生长速率测定:采用菌液点接法[14-15]接种,配制1×107个/mL的孢子悬浮液,用移液枪吸取5 μL滴于培养基正中间,等待菌液干后封好培养基,在恒温光照培养箱(RG-300)中培养;采用十字交叉法[16],定期用数显游标卡尺(ARZ-1331)测量菌落直径,并记录其菌落形态和颜色等。产孢量测定:培养15 d后,用直径为5 mm的打孔器,在培养基平板距离菌落中心位置到边缘1/2处打孔截取菌块,重复3次,分别放入含有10 mL 0.05%吐温-80无菌水的无菌容器中,涡旋振荡均匀,用微量移液器吸取适量孢子悬浮液滴在血球计数板上,如果显微镜视野下孢子数过多,以10倍进行稀释,至血球计数板上的孢子数可数为止,其中计算产孢量的公式中“80”代表血球计数板上80个正方形小格,静置30 s后在显微镜下观察和计数孢子数量,统计产孢量[17]。孢子萌发率测定:将菌株孢子配制成1.0×107个/mL的悬浮液,吸取1 mL加入含有10 mL SDAY液体培养基的小锥形瓶中,在摇床中恒温培养,培养24 h后在显微镜下用血球计数板观察孢子萌发情况,以芽管长度>孢子半径的孢子计为萌发孢子,观察n次视野,观察孢子总数>300个[18-19],计算萌发率。分生孢子大小及分生孢子梗测量:制作临时玻片在显微镜下找到分散的分生孢子和分生孢子梗,用莱卡光学显微镜(Leica,750)电脑配套软件LAS中的图片处理-距离线测量孢子的大小和孢子梗的长度等。
1.3 菌株的分子鉴定
病原菌在SDAY培养基培养7 d后,取一皿长势良好的菌刮取菌体,DNA提取使用CTAB法[20]略有改动,65℃恒温水浴时间由1 h变为30 min,将TE缓冲液换成ddH2O,其他步骤基本相同;采用真菌通用引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′),引物由生工生物工程(上海)股份有限公司合成;PCR反应体系25 μL,ITS1F和ITS4各1 μL,PCR MasterMix 12.5 μL,模板DNA 1 μL,ddH2O 9.5 μL;PCR反应条件:95℃预变性3 min;94℃变性1 min,55℃退火1 min,72℃延伸1.5 min,35个循环;1% 琼脂糖凝胶电泳检测,在凝胶成像系统下观察扩增结果。PCR产物送生工生物工程(上海)股份有限公司测序。
1.4 系统发育分析
依据测序得到的结果,将序列进行编辑,去除两端质量不好的碱基,优化好的序列提交到NCBI网站(http:∥blast.ncbi.nlm.nih.gov/Blast.cgi),选择BLAST进行比对,获得与目标菌株具有同源性的其他白僵菌菌株的碱基序列,从中下载同源性高的序列,使用MEGA 7.0软件通过邻接法(neighbor-joining method,NJ),运行1 000次bootstrap验证,构建菌株的系统发育树[21]。
1.5 菌株对草地贪夜蛾3龄幼虫的毒力研究
用0.05% 吐温-80无菌水冲洗培养基上的白僵菌分生孢子获得孢子悬浮液,并在摇床(210 r/min)中振荡30 min混匀,在显微镜下用血球计数板配制1.0×108、1.0×107、1.0×106、1.0×105个/mL和1.0×104个/mL 5个浓度的孢子悬浮液,采用浸渍法[22]接种,用无菌镊子挑选个体大小基本一致的草地贪夜蛾3龄幼虫,浸入孢子悬浮液中5 s,然后挑出置于滤纸上吸干菌液,选择活动性强的个体转移至已消毒的饲养盒,放入剪好的大小一致的新鲜玉米叶,每一处理20头,重复3次,对照组(CK)用0.05% 吐温-80无菌水处理。饲养盒为塑料格子盒(24格,单格尺寸4.2 cm×2.2 cm×3.5 cm),每虫1格单头饲养,放入(25±1)℃,RH(75±5)%,光周期L∥D=12 h∥12 h的恒温培养箱中饲养,每隔24 h更换新鲜的玉米叶和清理粪便,并观察供试草地贪夜蛾的行为及活动情况、虫体染菌情况,统计存活和死亡数量[23]。将死亡的虫体,在灭菌培养皿中于(25±1)℃光照培养箱中保湿培养,每天定时观察死亡虫体上菌体的生长和产孢情况,从而确定其为白僵菌感染致死。
1.6 数据处理
根据如下公式分别计算不同处理幼虫的死亡率、孢子萌发率和产孢量。毒力测定数据采用Probit方法分析,求出毒力回归方程,计算LC50和LT50,分析与计算均用SPSS 25.0 数据处理软件完成。


产孢量=孢子总数/80格×400×104×稀释倍数/打孔器面积(个/cm2)。
2 结果与分析
2.1 菌株分离培养及形态鉴定
YY-1菌株在25℃、SDAY培养基上的生长特性,菌落最外有很小一圈是菌丝生长扩散区域,白色绒毛状;中间大部分是产孢区域,孢子白色,后期白色稍偏暗粉状;菌落背面特征和正面相对应,正面凹陷的区域从背面看深入到培养基内部也比较明显,内圈深褐色,外圈淡黄色或橘色;分生孢子近球形,大小(1.664~2.523)μm×(2.097~3.628)μm,分生孢子梗弯曲,产孢结构端部膨大,分生孢子梗长55~120 μm,直径2.9~4.0 μm。
HNC-1菌株在25℃、SDAY培养基上,菌落正面颜色先为白色,后期变为淡黄色,在产孢区域有淡黄色透明液滴渗出,正面形态大致分为两个不同区域,最外圈是白色的菌丝生长扩散区,内部区域是白色稍偏暗的产孢区,孢子粉状,后期变为橘色;菌落背面大致分为二个区域,光滑无褶皱,外圈橘黄色,内圈黄褐色;分生孢子椭圆形,大小(1.622~3.183)μm×(2.250~4.420)μm,产孢结构二分叉。分生孢子梗长80~160 μm,直径2.3~3.7 μm。
2.2 菌株的分子鉴定
对菌株的测序结果进行编辑,去除序列两端质量不好的碱基,将其在NCBI数据库进行BLAST比对,YY-1菌株与NCBI数据库中的球孢白僵菌菌株Beauveriabassiana4510(KX901309.1)的亲缘关系最近,相似性高达99.43%,HNC-1菌株与NCBI数据库中的球孢白僵菌菌株BeauveriabassianaBbTDK005(HQ222969.1)的相似性为99.81%,说明该菌株与数据库中的球孢白僵菌菌株具有高度的相似性,综合形态特征鉴定和ITS序列相似性分析,将该菌株确定为球孢白僵菌。
2.3 菌株的生物学特性
YY-1和HNC-1菌株在SDAY培养基上25℃的生长产孢情况,菌落生长速率分别为(3.06±0.20)mm/d和(4.57±0.23)mm/d;培养第15天时的产孢量分别为(5.20±1.06)×107个/cm2和(1.83±0.47)×108个/cm2,其孢子振荡培养24 h后孢子的萌发率分别为(93.33±7.37)%和(87.67±6.43)%(表1);不同的温度条件下YY-1菌株的生长速率(F=9.824,P<0.05)、产孢量(F=14.425,P<0.05)和孢子萌发率(F=4.123,P<0.05)差异显著;HNC-1菌株的生长速率(F=27.422,P<0.05)、产孢量(F=28.728,P<0.05)和孢子萌发率(F=19.330,P<0.05)也有显著差异。试验结果表明,25℃是比较合适的生长产孢温度,温度过高或过低都不利于菌株的生长产孢。
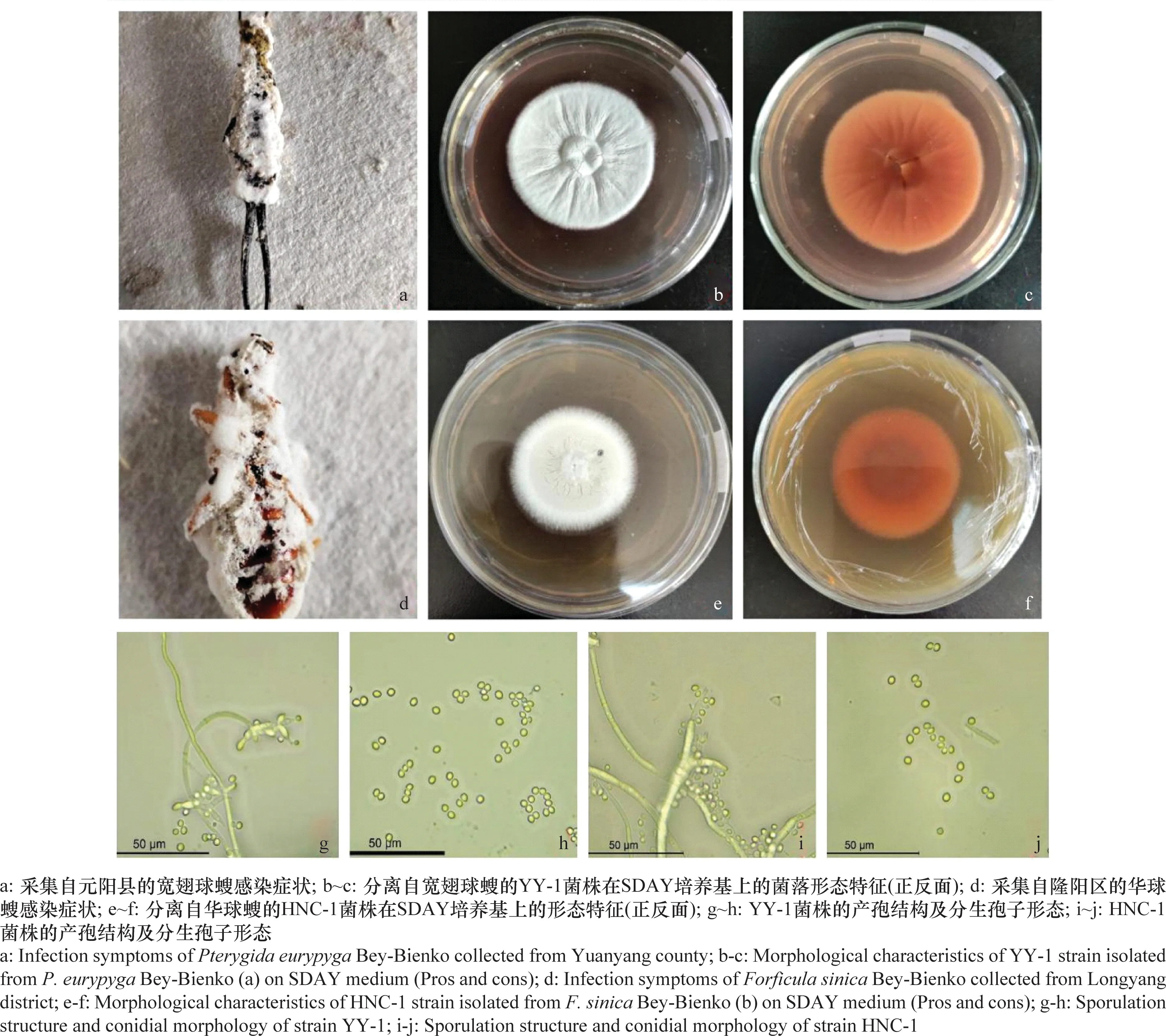
图1 蠼螋感染症状、白僵菌产孢结构和分生孢子的形态特征Fig.1 Field infection symptoms of earwig, sporulation structure and conidia of Beauveria bassiana

表1 球孢白僵菌YY-1和HNC-1在SDAY培养基上的培养特征1)Table 1 Culture characteristics of Beauveria bassiana YY-1 and HNC-1 on SDAY medium

图2 基于ITS基因序列构建YY-1和HNC-1菌株与其他球孢白僵菌的系统发育树(NJ 法)Fig.2 Construction of phylogenetic tree of the YY-1 and HNC-1 strain and other related Beauveria bassiana strains based on ITS region sequence by neighbour-joining method
2.4 菌株对草地贪夜蛾3龄幼虫的毒力
YY-1和HNC-1菌株在不同浓度孢子悬浮液下对草地贪夜蛾3龄幼虫具有不同程度的致死作用,随着孢子浓度的增加,致死作用逐渐增强,接种孢子悬浮液后最早于第2天开始出现幼虫死亡。在1.0×108个/mL浓度下,第8天草地贪夜蛾3龄幼虫累计死亡率达83.3%和63.3%,在此试验条件下YY-1菌株的毒力效果好于HNC-1菌株(图3)。
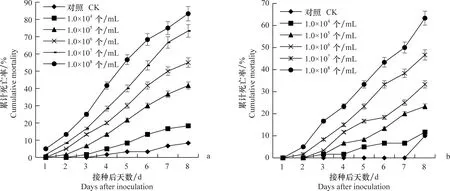
图3 YY-1(a)和HNC-1(b)菌株对草地贪夜蛾3龄幼虫的累计死亡率Fig.3 Cumulative mortality of YY-1 (a) and HNC-1 (b) strains to the third instar larvae of Spodoptera frugiperda
球孢白僵菌YY-1和HNC-1菌株对草地贪夜蛾3龄幼虫的LC50为1.56×105个/mL和2.11×105个/mL(表2)。YY-1和HNC-1菌株对草地贪夜蛾3龄幼虫的致死时间效应随其孢子悬浮液浓度的增加,草地贪夜蛾幼虫的致死中时缩短,当孢子悬浮液浓度从1×108个/mL降低到1×104个/mL,YY-1菌株的致死中时从4.23 d增加至20.23 d;HNC-1菌株的致死中时从6.12 d增加至22.14 d(表3)。可以看出在此试验条件下,YY-1菌株的效果好于HNC-1菌株。

表2 球孢白僵菌YY-1和HNC-1对草地贪夜蛾3龄幼虫的毒力Table 2 Toxicity of Beauveria bassiana YY-1 and HNC-1 to the third instar larvae of Spodoptera frugiperda
3 讨论
本研究从玉米田间采集到2头自然罹病的蠼螋成虫,目前关于革翅目蠼螋的病原物报道较为少见,可能与革翅目身体构造革质坚硬有关,本研究根据外形特征初步鉴定采集自元阳的蠼螋为宽翅球螋,采集自隆阳区的蠼螋为华球螋,但鉴定结果可能存在误差,因为部分身体形态已被真菌感染难以辨别;从2头蠼螋虫体上分离到2株球孢白僵菌YY-1和HNC-1,在SDAY培养基上,25℃下两株菌的生长产孢好于22℃和28℃,孢子萌发率无明显差异,最适生长温度与许多球孢白僵菌菌株相似[24-25]。
球孢白僵菌是一种常见的昆虫病原真菌,作为一种常规生物杀虫剂,具有高效、对非靶标生物较安全等特点,已被广泛应用于农林害虫的生物防治[26-27]。近年文献报道了多株球孢白僵菌对草地贪夜蛾幼虫的毒力,但是毒力效果差别较大,如分离于土壤的bbbj菌株对草地贪夜蛾3龄幼虫的LC50为3.37×105个/mL,毒力比较高;而另一株bbhn菌株在108个/mL孢子浓度下处理7 d对草地贪夜蛾3龄幼虫的致死率低于50%[28]。分离自稻黑蝽Scotinopharalurida若虫的GZSL-1菌株,在孢子浓度1×108个/mL时对草地贪夜蛾3龄幼虫的累计校正死亡率可达100%[29];而ZJU435菌株对草地贪夜蛾3龄幼虫接种10 d后的校正死亡率为23.3%,对4龄幼虫几乎无杀虫活性[30];本试验从蠼螋上分离到的球孢白僵菌YY-1对草地贪夜蛾3龄幼虫致病力较高,HNC-1菌株致病力一般。此外,本研究中HNC-1菌株虽然生长产孢较YY-1菌株好,但对草地贪夜蛾幼虫的毒力效果却低于YY-1菌株,因此还应加强对YY-1菌株培养条件的研究,以期为该菌株开发提供依据。
猜你喜欢
中国生物防治学报(2022年3期)2022-07-09
检验医学(2021年11期)2021-11-29
疯狂英语·新策略(2021年9期)2021-11-02
湖南林业科技(2021年3期)2021-07-02
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
中国抗生素杂志(2019年6期)2019-07-06
热带作物学报(2019年4期)2019-06-11
湖北林业科技(2019年2期)2019-05-05
中国森林病虫(2019年2期)2019-01-11
