
围封、放牧、刈割对内蒙古典型草原土壤线虫群落的影响
2022-08-04 01:29侯林秀孙浩宇徐智超王永慧王立新
草地学报 2022年7期
侯林秀, 张 斌, 孙浩宇, 徐智超, 王永慧, 温 璐, 王立新
(内蒙古大学生态与环境学院, 内蒙古 呼和浩特 010021)
线虫作为陆地生态系统中数量最多、分布广泛的土壤后生动物类群,在土壤有机质分解和养分循环方面起重要作用[1-2]。它们常被看作是指示土壤健康状况、衡量生态系统抵抗力和恢复力的重要生物[3],这主要归因于土壤线虫对环境变化更为敏感的反应能力、分离鉴定方法的易操作性、食性的多样性以及与其他生物群落更为紧密的关联性等[4]。这些特点决定它们可以通过自身代谢活动和取食活动等多种方式影响土壤有机质分解[5]。近年来,越来越多的学者针对土壤线虫群落展开研究,集中于探讨线虫群落对不同生物因素(包括植被类型、微生物、其他小动物)[6-8]及非生物因素(气候变化、土质和土层深度、土壤理化性质等)[9-12]的响应机制。
内蒙古典型草原是中国北方温带草原的主体,在碳氮循环、气候调节等方面具有重要作用,同时也是当地居民赖以生存的基础[13-14]。围封、放牧和刈割作为典型草原最主要的草地利用方式,能改变草地生产力及土壤有机碳输入的数量和质量等[15-16],也会对土壤线虫群落产生影响[17-18]。已有学者分析土壤线虫群落对草地利用方式的响应情况,但基本上仅针对一种或两种利用方式展开研究,未同时对比围封、放牧、刈割对土壤线虫的影响,且互相之间尚未形成一致的结论[19-21]。此外,大部分草地位于生态脆弱带上,除区域性人为干扰所导致的环境变化对土壤线虫群落产生显著影响之外,线虫群落对全球性气候变化也十分敏感[22]。近年来,地表气温、CO2浓度、氮沉降量等有明显的升高趋势,并将持续性上升,降水格局也将发生改变[23-24],在此影响下,土壤线虫群落对草地利用方式的响应情况必将会产生新的变化,不同的研究时间和环境背景在很大程度上决定了前人研究结果不相同。
因此,探究当前气候环境下不同草地利用方式对温带典型草原土壤线虫群落组成的影响十分必要。通过揭示围封、放牧和刈割处理下土壤线虫种类、丰度、生态特征指数的变化规律,综合评估不同利用方式下草地受干扰程度和生态系统可持续利用潜力,以期为全球环境变化大背景下实现高效合理的放牧管理提供生物学依据。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区锡林浩特市毛登牧场内蒙古大学草地生态学实验基地(44°10′ N,116°28′ E),海拔1 102 m,属典型温带半干旱大陆性气候,年平均气温2.36℃,年平均无霜期约为150 d,年平均降水280 mm左右,且降雨多集中在6-9月,约占全年的80%。土壤类型以栗钙土为主,土壤pH值平均为8.07,地上植物优势种为羊草(Leymuschinensis)、克氏针茅(Stipakrylovii)等[25-26],属典型草原。
1.2 试验设计
实验平台建立于2012年,选取地势平坦、土壤类型相近的3个100 m×100 m样区,样区之间的间距为33 m,按照随机区组设计在每个样区设置3个大小相同的样地(33 m×33 m),间距约为1 m,分别对应围封(F)、放牧(G)、刈割(C)3种利用方式。每年7-8月在放牧样地连续放牧6只羊,持续到植物留茬高度为6 cm(该高度被认为是草地可持续利用的合理高度[27],以研究区优势种高度为准);刈割处理在每年8月下旬进行,植物留茬高度为6 cm;围封处理不实施人为干预。
1.3 样品采集及数据测定
于2019年8月采用多点混合采样法采集土样,每个样地沿对角线选取3个样点,用直径为8 cm的土钻钻取0~10 cm土壤3钻,同一样地的9钻土壤混合为1个样品,共计9个土样,尽快带回实验室置于-80℃冰箱内冷冻。运用E.Z.N.A.®soil试剂盒(Omega Bio-tek,Norcross,GA,U.S.)对土壤样品进行总DNA提取,DNA质量利用1%琼脂糖凝胶电泳进行检测;采用引物nf1F-18sr2bR,即nf1F5′-GGTGGTGCATGGCCGTTCTTAGTT-3′和18sr2bR5′-TA CAAAGGGCAGGGACGTAAT-3′进行PCR扩增[28],正式试验则通过TransGen AP221-02进行。委托上海美吉生物医药科技有限公司利用Illumina Miseq PE 3000平台进行土壤线虫测序,测序数据比对物种注释数据库为NCBI(NT)。
此外,在8月上旬的生物量峰值阶段于每个样地随机选择1 m×1 m的小样方,记录样方内的植物物种数以表征植物物种丰富度。
1.4 土壤线虫群落生态学指数计算
用Jaccard相似性指数(ISj)[29]表征线虫群落相似性:
(1)
用丰富度指数(S)、Shannon指数(H′)、优势度指数(λ)、均匀度指数(J′)来衡量土壤线虫的物种多样性,用以描述线虫群落在局域生境内的类群数量和分布情况[30]。利用成熟度指数(Maturity index,MI)、植物寄生线虫成熟度指数(Maturity index of plant parasite,PPI)、基础指数(Basic index,BI)、结构指数(Structure index,SI)、富集指数(Enrichment index,EI)来表征线虫群落结构复杂度和土壤养分富集等[31]。
(2)
(3)
(4)
式中,S为群落物种数目;Pi为物种i的相对重要值,即物种i的个体在全部个体中的比例。
(5)
(6)
式(5)中,vi为第i类自由生活线虫的Colonizer-persister值(c-p值),Pi为物种i的相对重要值,即物种i的个体在全部个体中的比例。
式(6)中,vi为第i类植物寄生线虫的c-p值,Pi为物种i的相对重要值,即物种i的个体在全部个体中的比例。
BI=100×[b/(e+b+s)]
(7)
SI=100×[s/(b+s)]
(8)
EI=100×[e/(e+b)]
(9)
式中,b为线虫基础组分,包括c-p值为2的食细菌线虫、食真菌线虫;e为线虫富集组分,包括c-p值为1的食细菌线虫和c-p值为2的食真菌线虫;s为线虫结构组分,包括c-p值为3~5的食细菌线虫、食真菌线虫和杂食-捕食性线虫。
1.5 数据处理
在上海美吉生物医药科技有限公司云平台进行在线分析(https://cloud.majorbio.com/),在线虫属水平上利用基于Abund-Jaccard算法的ANOSIM分析反映组间差异,并基于Bray-Curtis距离算法对不同样地的土壤线虫群落进行主坐标分析(Principal coordinate analysis,PCoA)分析。采用SPSS 22软件对线虫营养类群相对丰度和线虫区系指数进行单因素方差分析,并对样地地上植物丰富度与土壤线虫丰富度指数进行线性回归。冗余分析(Redundancy analysis,RDA)使用Canoco 5完成。此外,为分析土壤线虫群落的代谢活动特征,表征生态系统受干扰程度,将线虫结构指数和富集指数整合在同一个坐标系内划分象限进行分析[1,7](图1)。绘图使用Origin 2019b软件。

图1 土壤线虫群落结构指数、富集指数权重区系分析的意义解释Fig.1 The significance of analysis of soil nematode fauna weight between SI and EI注:图中Ⅰ、Ⅱ、Ⅲ、Ⅳ为象限分区,具有指示意义。下同Note:In the figure,Ⅰ,Ⅱ,Ⅲ,and Ⅳ are quadrant zones with indicative significance. The same as below
2 结果与分析
2.1 土壤线虫测序结果及群落组成
如表1所示,所有土壤样品总计测得线虫有效序列2 404 条,共归为38 个操作分类单元(OTU),分属于3纲6 目16 科18 属19 种。对比不同利用方式样地,土壤线虫在科、属、种水平上均呈现围封>放牧>刈割的规律。在考虑物种丰度影响的前提下,ANOSIM距离分布箱式图结果表明,不同样地处理之间存在显著差异(ANOSIM=0.383,P=0.007)(图2)。

表1 不同利用方式下草地土壤线虫群落的测序结果统计Table 1 Statistics of soil nematode community sequencing results under different utilization modes
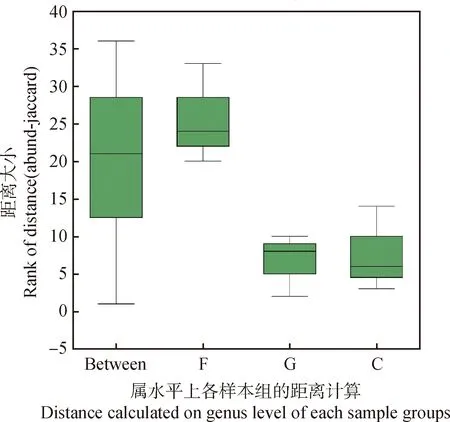
图2 不同利用方式下草地土壤线虫组内、组间样本距离分布箱式图Fig.2 Box plot of soil nematode sample distance distribution within and between groups under different utilization modes注:Between指的是样地之间的差异,其他分别代表各自组内差异Note:Between refers to the differences Between sample sites,while the others respectively represent differences within their respective groups
将土壤样本中相对丰度小于1%的线虫归为其他线虫属。基于线虫属水平,在围封、放牧、刈割样地中,未分类线虫相对丰度更高,分别占各样地土壤线虫群落的30.99%,35.09%,38.60%(图3)。除未分类线虫外,8个线虫属为3个样地所共有,分别是吻球属(Hoplotylus)、Amblydorylaimus、牙咽属(Dorylaimellus)、拟丽突属(Acrobeloides)、针属(Paratylenchus)、真头叶属(Eucephalobus)、丝尾垫刃属(Filenchus)、轮属(Criconemoides)。不同利用方式下温带典型草原土壤线虫群落组成差异较大,围封、放牧、刈割样地中占土壤线虫群落主导地位的线虫属分别是未分类的细齿科(unclassified_Leptonchidae)、吻球属(Hoplotylus)、牙咽属(Dorylaimellus),在对应样地的相对丰度分别为31.96%,28.82%和28.38%。
为消除各样地之间未分类线虫的影响,对能具体鉴定到属或个别无法详细到属水平但能确定科水平的土壤线虫重新计算其相对丰度,结果如表2所示。所有样地共有线虫17 属,其中围封样地共检测到线虫13 属,群落优势类群(个体数量占10%以上的类群)为未分类的细齿科(unclassified_Leptonchidae)、针属(Paratylenchus)和拟丽突属(Acrobeloides);放牧样地共有线虫12 属,优势类群为拟丽突属(Acrobeloides)、吻球属(Hoplotylus)、Amblydorylaimus;刈割样地共有线虫10 属,Amblydorylaimus、牙咽属(Dorylaimellus)、吻球属(Hoplotylus)、拟丽突属(Acrobeloides)为该样地线虫优势属。所有样地中,土壤线虫常见属有轮属(Criconemoides)、真头叶属(Eucephalobus)、丝尾垫刃属(Filenchus)等,茎属(Ditylenchus)、独壁齿属(Campydora)、筒咽属(Cylindrolaimus)等为稀有属(个体数量占总体数量1%以下的类群)。
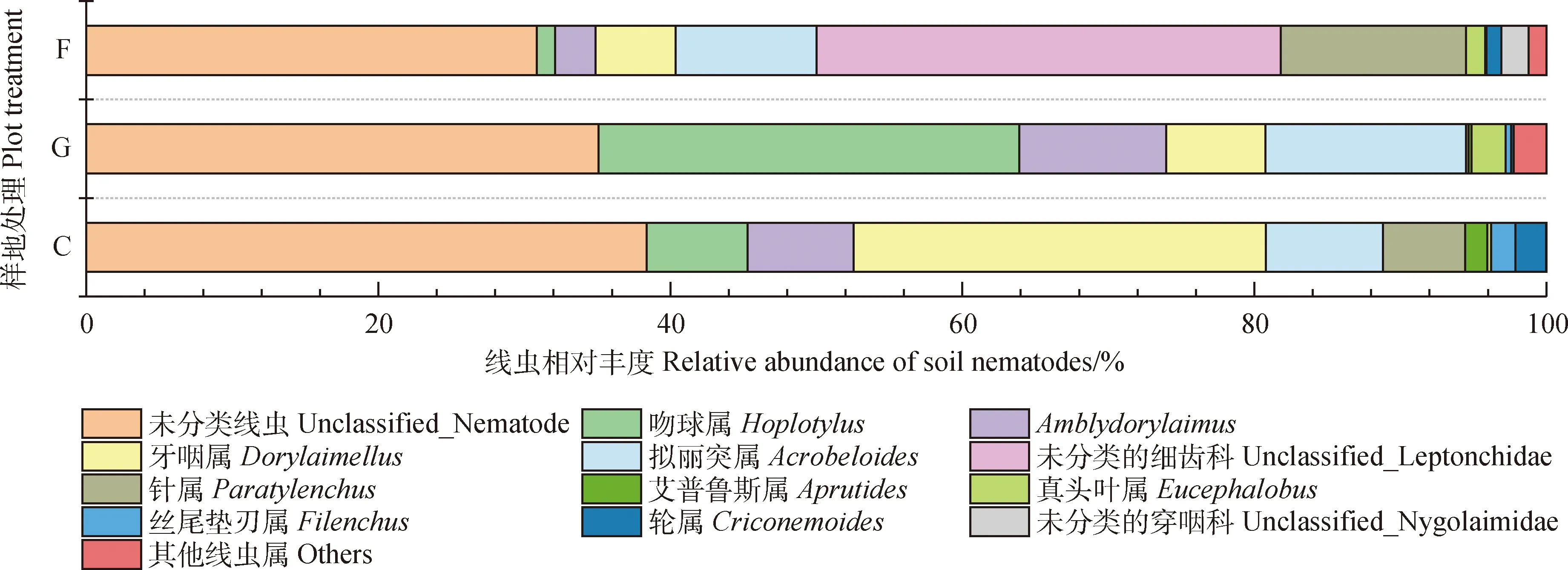
图3 属水平上不同利用方式下草地线虫群落相对丰度占比Fig.3 Proportion of nematode community relative abundance under different utilization modes on the genus level

表2 不同利用方式下草地土壤线虫群落组成及营养类群、c-p值(不包括未分类线虫)Table 2 Soil nematode community composition,functional group and c-p value under different utilization modes (excluding unclassified nematodes)
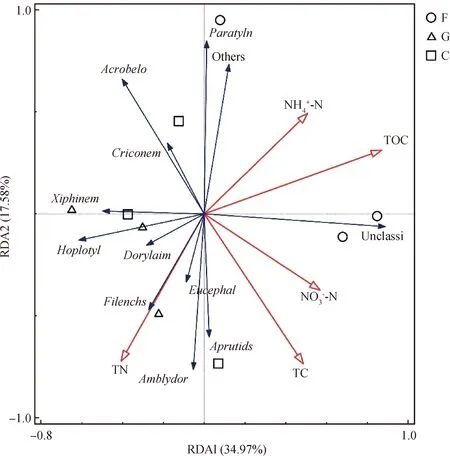
图4 土壤理化性质与土壤线虫群落的冗余分析Fig.4 Redundancy analysis (RDA) of soil physical and chemical properties and soil nematode community注:Acrobelo代表拟丽突属,Amblydor为Amblydorylaimus,Aprutids代表艾普鲁斯属,Criconem代表轮属,Dorylaim代表牙咽属,Eucephal代表真头叶属,Filenchs代表丝尾垫刃属,Hoplotyl代表吻球属,Paratyln代表针属,Xiphinem代表剑属,Unclassi为unclassified_Leptonchidae(未分类的细齿科),Others代指除未分类线虫以外,所有相对丰度小于1%的线虫类群。TC为总碳,TN为全氮,TOC为总有机碳,和分别为铵态氮和硝态氮Note:Acrobelo is Acrobeloides,Amblydor is Amblydorylaimus,Aprutids is Aprutides,Criconem is Criconemoides,Dorylaim is Dorylaimellus,Eucephal is Eucephalobus,Filenchs is Filenchus,Hoplotyl is Hoplotylus,Paratyln is Paratylenchus,Xiphinem is Xiphinema,Unclassi is unclassified_Leptonchidae,Others refers to all nematode groups with relative abundance less than 1% except for unclassified nematodes
2.2 土壤线虫组间差异分析
PCoA分析结果显示P值为0.054 0,3种草地利用方式下的线虫群落结构未呈现显著性差异,2个主成分分别可以解释37.13%和24.6%的群落结构变化。但围封样地与放牧、刈割样地相比,其土壤线虫结构组成有相对更为明显的差异性(图5)。
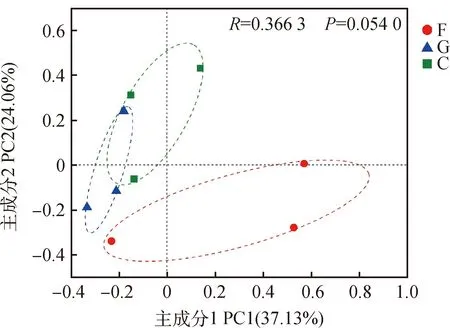
图5 属水平上不同利用方式下草地线虫群落结构的PCoA分析Fig.5 PCoA analysis of nematode community structure under different utilization modes on the genus level
为进一步探讨土壤线虫在种水平上的组间差异性,计算不同样地之间的土壤线虫相似性指数,可得出放牧和刈割样地的相似性指数最高,为0.67;其次是放牧和围封,为0.59;围封和刈割样地的土壤线虫群落组成相似性指数最低,为0.53(表3)。

表3 不同利用方式下草地土壤线虫群落组成Jaccard相似性指数Table 3 Jaccard index of soil nematode community composition between different utilization modes
2.3 土壤线虫营养结构
线虫的营养类型和c-p值如表2所示,所有样地中共测得植物寄生线虫6属、食细菌线虫4属、食真菌线虫4属、杂食-捕食性线虫3属。3种利用方式下的属于植物寄生线虫的属均多于同一样地其他线虫类群,具体为围封5属、放牧5属、刈割4属;食细菌线虫属数表现为围封>放牧>刈割;与其他样地相比,放牧条件下食真菌线虫属最多,但其相对丰度在三个样地均低于1%;在放牧样地中杂食-捕食性线虫所包含属数最少,仅有Amblydorylaimus一种,同时该属线虫在围封和刈割样地杂食-捕食性线虫中占比分别为52.08%和99.71%,是整个研究区杂食-捕食性线虫的重要组成部分。
植物寄生线虫在放牧和刈割样地中占据主导地位,其相对丰度分别为57.75%和67.41%,而在围封处理下其相对丰度仅为29.89%(图6)。此外,围封样地中食真菌线虫的相对丰度显著高于其余两个样地(P<0.01),达47.15%。与其他线虫类群相比,杂食-捕食性线虫丰度在围封处理下最小,为6.87%,但在放牧和刈割样地中其丰度可达15%以上。食细菌线虫在3种不同处理方式下的相对丰度差异不显著。从线虫营养类群组成看,放牧和刈割样地之间不存在显著差异。
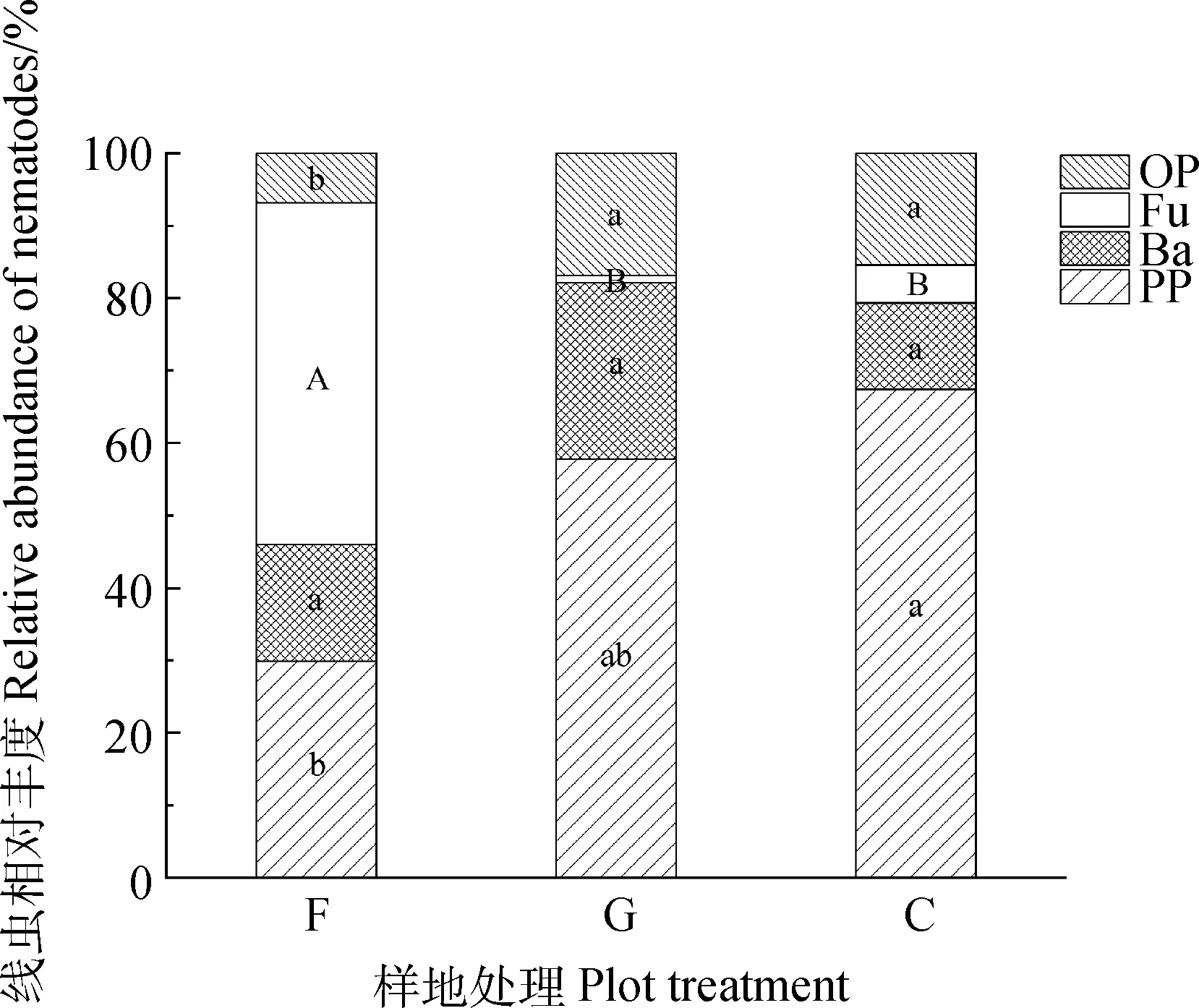
图6 不同利用方式下各营养类群线虫群落相对丰度占比(不包括未分类线虫)Fig.6 Relative abundance ratios of nematodes in different trophic groups under different utilization modes (excluding unclassified nematodes)注:同一线虫营养类群数据中a,b表示在0.05水平上样地间存在显著差异性,A,B表示在0.01水平上样地间存在显著差异性Note:In the data of the same functional group of nematode,a and b indicated that there were significant differences among the sample plots at the 0.05 level,A and B indicated that there were significant differences among the sample plots at the 0.01 level
2.4 土壤线虫区系分析
基于OTU水平计算不同草地利用方式下线虫物种多样性指数和线虫功能团指数(表4)。不同利用方式对草地土壤线虫群落丰富度指数影响不显著,但显著影响线虫群落的Shannon指数、优势度指数和均匀度指数(P<0.05)。各处理间Shannon指数和均匀度指数均表现为围封和放牧样地显著高于刈割样地(P<0.05),而刈割样地土壤线虫的优势度指数显著高于围封和放牧样地(P<0.05)。虽然土壤线虫丰富度指数大小对利用方式的响应不显著,但研究区地上植物丰富度与土壤线虫丰富度指数的线性回归结果表明在P<0.05水平上二者之间呈现显著线性关系(图7)。

表4 不同利用方式下土壤线虫群落物种多样性指数Table 4 Statistics of soil nematode community species diversity index under different utilization modes
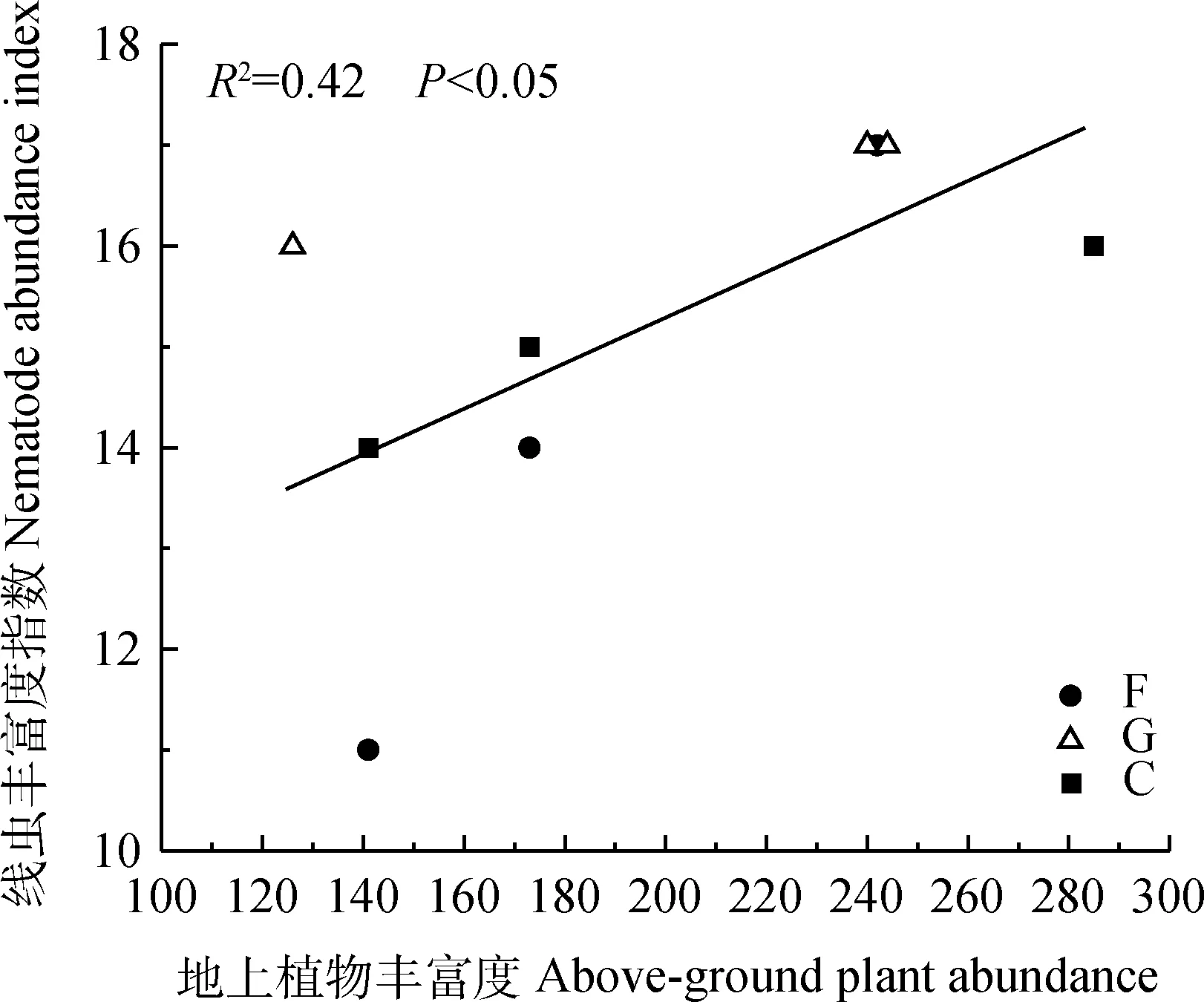
图7 研究区土壤线虫丰富度指数与地上植物丰富度的线性回归分析Fig.7 Linear regression analysis of soil nematode abundance index and above-ground plant abundance in the study area
受利用方式的影响,放牧样地的成熟度指数显著低于刈割和围封样地(P<0.05),而围封样地植物寄生线虫成熟度指数显著低于放牧和刈割样地(P<0.05)(表5)。放牧样地的基础指数显著高于刈割样地(P<0.05),而围封样地显著低于刈割样地(P<0.05)。结构指数整体变化规律性与基础指数相反,为围封>刈割>放牧,且围封与其他处理间差异显著(P<0.05)。刈割处理下的线虫富集指数显著高于其他利用方式(P<0.05)。土壤线虫结构指数、富集指数权重区系分析结果显示围封样地落于Ⅲ象限,未受到明显的外界干扰,食物网稳定;而放牧和刈割样地落于Ⅳ象限,C∶N高,受到环境胁迫,食物网呈现一定的退化特征(图8)。

表5 不同利用方式下土壤线虫功能团指数对比Table 5 Comparison of soil nematode functional group index under different utilization modes

图8 不同利用方式下土壤线虫群落结构指数、富集指数权重区系分析Fig.8 Analysis of soil nematode fauna weight between SI and EI under different utilization modes注:图中横线表示结构指数的标准差,竖线表示富集指数的标准差Note:In the figure,the horizontal line represents the standard deviation of the structure index,the vertical line represents the standard deviation of the enrichment index
3 讨论
3.1 不同利用方式对土壤线虫群落特征的影响
在属水平上,除未分类的土壤线虫以外,围封、放牧、刈割样地中线虫优势类群均不相同,说明草地利用方式在一定程度上改变了线虫群落结构组成。而基于Abund-Jaccard算法的ANOSIM分析与基于Bray-Curtis距离算法的PCoA分析结果表明利用方式对土壤线虫群落结构的影响主要是由于线虫丰度差异造成,而非类群差异。例如放牧显著增加了吻球属(Hoplotylus)的相对丰度,而刈割使得牙咽属(Dorylaimellus)丰度显著增大。
同时,群落生物组成结构可以通过物种多样性来量化,用以反映群落内物种的多少以及各生境之间的相似性和差异性[33]。与其他2种处理方式相比,刈割对温带典型草原土壤线虫群落Shannon指数、优势度指数和均匀度指数均影响显著(P<0.05)。与围封样地相比,刈割处理对线虫多样性指数的影响大于放牧处理,出现这一现象的原因,可能与放牧环境下羊在进行选择性取食的同时留下粪便等有机物促进土壤养分循环有关。研究表明不同草地利用方式会对土壤线虫群落结构产生影响,并形成组间差异。这可能是因为草地土壤养分状况会随着人为干扰而发生变化[34],同时土壤动物的生存受到土壤环境和营养状况制约[35]。土壤线虫群落与土壤理化性质的冗余分析结果对这一结论起到一定支撑作用,有机碳含量显著影响了线虫群落结构。
线虫群落特征在受到土壤因素影响的同时,与地上植物也存在紧密相关关系。植物群落的物种多样性及其特性会对土壤线虫群落产生影响[36-37],土壤线虫也会对植物生长产生反馈作用间接影响植物生产力发生变化[38-39]。一方面,研究表明地上植物丰富度指数与土壤线虫丰富度指数相关性显著(P<0.05);另一方面,该研究区的地上植物总量为:放牧>刈割>围封[40],这与本研究土壤线虫丰富度指数大小的结果一致。与物种多样性指数结果不同的是,在不考虑线虫类群相对丰度大小而只考虑线虫物种重叠度(即是否存在)的情况下,放牧与刈割样地的线虫群落组成相似度更高,说明与围封相比,由于地上生物量的减少,牛羊啃食和刈割使得土壤线虫在草地生态系统中所处的环境背景更为接近,进而导致二者的线虫群落结构更为类似,即植物生长状况会对土壤线虫产生一定影响。
3.2 土壤线虫对生态系统的指示功能
线虫作为土壤生态系统的重要指示生物之一,其相关指标能较为准确地反映土壤状况。线虫群落中各营养类群所占比例大小可以用来指示生态系统健康程度的变化[41]。有研究表明植物在人为干扰下更易被植物寄生类动物所侵害[42],本研究结果显示刈割和放牧样地中植物寄生线虫的相对丰度均显著高于围封样地(P<0.05),该结果在一定程度上支撑了前人的研究结论。Wasilewska[43]曾提出,当其他生态系统向牧场生态系统转变时,食细菌线虫数量增多,而食真菌线虫丰度有所降低。本研究结果与该结果基本一致,即与其他样地相比,放牧样地中食细菌线虫相对丰度最高而食真菌线虫相对丰度最低。在放牧活动干扰下,牛羊啃食和粪便堆积为细菌生长繁殖提供更为有利的环境条件,细菌所参与的降解途径更为活跃[44],这可能是食细菌线虫数量增多的主要原因。杂食-捕食性线虫是土壤微食物网中营养级别较高的参与组成部分,也是决定其复杂程度的关键生物类群[45],放牧和刈割处理下的杂食-捕食性线虫相对丰度显著高于围封对照组(P<0.05),表明适当的外部干扰会使得土壤线虫群落结构组成趋于复杂,放牧处理下相对更高的线虫物种多样性也在另一层面反映了这一规律。
除线虫营养类群外,不同的草场利用方式也显著改变了土壤线虫功能团指数,并对后续草场管理实践有指示作用。研究表明围封样地的土壤线虫群落结构化程度高,连通性强;而其余两种草地利用方式下的线虫群落在外界干扰下呈现一定的退化特征,其中放牧条件下生态系统干扰退化程度更强,土壤线虫群落对外部环境干扰的相对抵抗力有增大的趋势。这些均与乌尼尔等[19]对典型草原0~10 cm土壤线虫的研究结果相一致,在一定程度反映草地生态系统在外界干扰下的自我调节能力可以作用于土壤线虫群落,并通过土壤线虫的指示作用传达出来。此外,刈割样地的富集指数显著高于其他处理(P<0.05),表明刈割处理对土壤的养分富集有促进作用,其可能原因是与轻度放牧或不放牧利用相比,合理有序的刈割更有利于植被更新,增加土壤碳库含量[46]。围封、放牧和刈割作为常见的草场管理手段,在后续实践中应得到合理利用,既要合理控制放牧强度,以免加快草场退化进程,同时也要进行间歇性围封和刈割处理,分别以增强土壤微食物网的连通性和促进土壤养分循环和富集。
4 结论
围封、放牧和刈割改变了土壤线虫群落的类群数量、结构组成和生态指数。基于线虫属水平,线虫群落受到土壤有机碳含量的显著影响;按营养类群分类,围封样地的线虫群落结构与其它两个样地差异显著。围封和刈割样地的线虫群落组成相似性指数最低,在围封环境下,线虫连通性和捕食关系有所加强,线虫群落结构化程度高;刈割则显著降低了线虫Shannon指数和均匀度指数,而且有利于土壤养分富集。线虫丰富度与地上植物丰富度呈显著正相关关系,草地利用方式的改变或许会通过植物生长状况间接影响土壤线虫。
猜你喜欢
绿色天府(2022年9期)2022-11-19
今日农业(2022年14期)2022-09-15
世界热带农业信息(2022年8期)2022-07-19
新农村(浙江)(2022年7期)2022-07-07
贵州畜牧兽医(2022年3期)2022-06-28
中国生物防治学报(2022年1期)2022-04-22
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
江苏农业科学(2017年17期)2017-11-15
山西果树(2014年6期)2015-03-12
