
扇穗茅不同居群染色体数目及核型分析
2022-08-04 01:50:16刘玉萍王亚男胡夏宇陈金元
草地学报 2022年7期
杨 萍, 苏 旭,3, 刘玉萍*, 王亚男, 胡夏宇, 陈金元
(1.青海师范大学生命科学学院, 青海 西宁 810008; 2.青海师范大学青海省青藏高原药用动植物资源重点实验室,青海 西宁 810008; 3.高原科学与可持续发展研究院, 青海 西宁 810016)
扇穗茅(Littledalearacemosa)是禾本科(Poaceae)雀麦族(Bromeae)扇穗茅属(Littledalea)的一种高原特有疏丛短根茎禾草,主要分布于青藏高原及其毗邻高山地区,仅有少部分居群处于邻近的稍低海拔地区[1]。扇穗茅通常生长于高山草坡、河谷边沙滩、灌丛草甸等极端环境下,是高山草甸的伴生种,具有较强的耐旱、耐寒、耐碱、耐贫瘠及抗病和抗风沙能力[2];同时,该物种具有相对发达的根系,是保土固沙的一种重要植物;营养价值较高,也是畜牧业上的优良牧草[3]。目前,国内外学者已对扇穗茅的形态特征、地理分布、分类修订、叶绿体基因组特点及种质资源等进行了研究[3-6]。譬如,周勇辉等[6]采用广义形态性状将藏扇穗茅归并到扇穗茅中,作为异名处理;刘玉萍等[3]通过Illumina MiSeq高通量测序技术,探讨了扇穗茅的叶绿体基因组特点及系统进化关系,建立了雀麦族物种的标准测序流程,认为其与小麦族(Triticeae)物种亲缘关系较近。然而,目前从居群水平上对扇穗茅染色体的形态特征、核型参数及进化关系的研究未见报道。
染色体是细胞遗传物质的载体[7],其形态、数目和结构往往受外界影响较小[8],在植物生长发育和繁殖过程中具有较高的稳定性。因此,通过对植物细胞染色体形态、数目、特征及核型参数的综合分析,能够有效探讨不同物种或品种间的细胞学差异,从而阐明它们的起源和进化关系,为今后植物物种界定、遗传育种和新品种选育提供细胞学佐证[9]。迄今为止,许多学者采用植物染色体常规压片技术对多种禾本科植物染色体核型的研究揭示了它们的相对进化程度及系统发育关系。譬如,张同林等[10]通过对小麦族披碱草属(Elymus)、鹅观草属(Roegneria)和赖草属(Leymus)共5个类群的核型分析,认为赖草属进化程度最低、鹅观草属最高、披碱草属介于两者之间;杨慧芳[11]利用火焰干燥法对冰草(Agropyroncristatum)不同居群进行了染色体制片并对核型作了分析,结果发现冰草的中亚、东亚与青藏高原居群间以及欧洲与中亚居群间亲缘较近,中亚居群起源较早且进化较原始、欧洲居群起源相对较晚且进化程度较高,认为中亚是冰草的起源中心,染色体结构和数目变异是对新环境的适应;王亚男等[12]通过对沙鞭(Psammochloavillosa)6个不同居群染色体形态和核型的研究,发现沙鞭染色体数目恒定,有正中部着丝粒(Median point chromosome,M)、中部着丝粒(Median region chromosome,m)、近中部着丝粒(Submedian region cheomosome,sm)或近端部着丝粒(Subterminal region chromosome,st)4种类型,并推导了不同居群间的系统进化关系。据此,本研究采用染色体常规压片技术,对扇穗茅6个自然地理居群的染色体数目、形态、核型参数等进行系统分析;在此基础上,探讨不同居群间的核型差异和亲缘关系,为扇穗茅系统进化和优良种质资源筛选提供细胞学佐证。
1 材料与方法
1.1 试验材料
本研究选取扇穗茅6个自然地理居群的成熟种子为试验材料,具体采样信息详见表1。凭证标本保存于中国科学院西北高原生物研究所青藏高原生物标本馆(QTPMB)。

表1 试验材料及来源Table 1 Experimental materials and sources
1.2 试验方法
1.2.1种子萌发 从扇穗茅6个代表居群中挑选饱满成熟的种子,且每个居群中选取10粒种子进行萌发。首先将种子在—20℃冰箱中处理24 h,然后将其均匀放置于铺有蒸馏水湿润两层滤纸的培养皿中[13],随后置于22℃恒温培养箱中培养,当种子根长为0.5~1 cm时取样预处理。
1.2.2预处理 预先将2.0 mL离心管用蒸馏水喷湿,保持管内外湿润;然后,将剪下的扇穗茅根尖放于其中,立即将盛有根尖的离心管置于充满N2O的密封罐中处理130 min,压强设置为0.75 MPa。
1.2.3固定与解离 从密封罐取出离心管,预处理的根尖迅速放入含卡诺氏固定液(95%酒精∶冰醋酸=3∶1,现配现用)的离心管中,使根尖充分浸入固定液,随后将离心管置于4℃冰箱中固定10 min以上,最后用45%乙酸解离5 min待用。
1.2.4制片、染色和观察 解离好的根尖轻轻取出并放置于载玻片中央,用刀片切除多余部分仅保留根尖顶端分生组织,滴一滴改良的石炭酸品红溶液,染色7~8 min后盖上盖玻片,用镊子尖端轻轻敲击使其充分均匀分散,迅速将载玻片放置于酒精灯上轻轻灼烧,待组织区域的液体蒸干后,用滤纸垂直向下按压载玻片,保持细胞均匀分散,利用显微镜(Olympus BX53)观察染色体的形态和数目,选取分散良好、形态清晰的染色体拍照[14]。
1.2.5数据分析 从扇穗茅每个居群中,选取10个分散良好、形态清晰、数目完整的染色体中期分裂图拍照,同时进行染色体形态观察、数目统计及数据测量与分析。即采用Adobe Photoshop 2020软件测量所有居群的染色体长臂与短臂长度[15],将数据导入Excel软件中统计分析;依据李懋学[16]方法计算染色体总长度、臂比(Arm ratio,AR)、相对长度(Relative length,RL);基于这些数据进行每个细胞同源染色体配对,统计同源染色体长臂和短臂平均值及总长度等参数,主要包括平均臂比(Mean arm ratio,MAR)、染色体长度比(Chromosome length ratio,CLR)、核不对称系数(Karyotype asymmetry coefficient,As.K%)及臂比大于2∶1的染色体百分比(Chromosome proportion of arm ratio excess to 2)等,其中臂比大于2∶1的染色体百分比=臂比大于2的同源染色体数目/同源染色体组总数;核不对称系数=(染色体长臂总长/全组染色体总长)×100%;然后,根据同源染色体核型参数,按照相对长度从大到小排序、着丝点排列在同一直线上、短臂在上、长臂在下,利用Excel和Photoshop软件绘制染色体核型图。此外,本研究涉及的相对长度系数(Relative length coefficient,I.R.L)依据Kuo等[17]方法、核不对称系数根据Arano[18]方法及核型分类依照Stebbins[19]方法进行计算、统计和分析。
另外,参照邢世岩等[20]方法进行核型进化趋势分析,利用Microsoft Excel 2010软件绘制扇穗茅核型不对称性程度散点图,即核型二维进化图;同时,聚类分析依据李晓莉等[21]方法,采用SPSS 26软件统计分析染色体臂比平均值与方差、相对长度方差、染色体长度比和正中部着丝粒(M)、中部着丝粒(m)、近中部着丝粒(sm)或近端部着丝粒(st)染色体比例等,进行扇穗茅不同居群的染色体聚类分析。
2 结果与分析
2.1 染色体数目与倍性鉴定
通过对扇穗茅中期分裂相的观察和测量,发现扇穗茅染色体均为二倍体,数目较为稳定,为2n=2x=28,不存在染色体数目和形态异常现象(图1)。
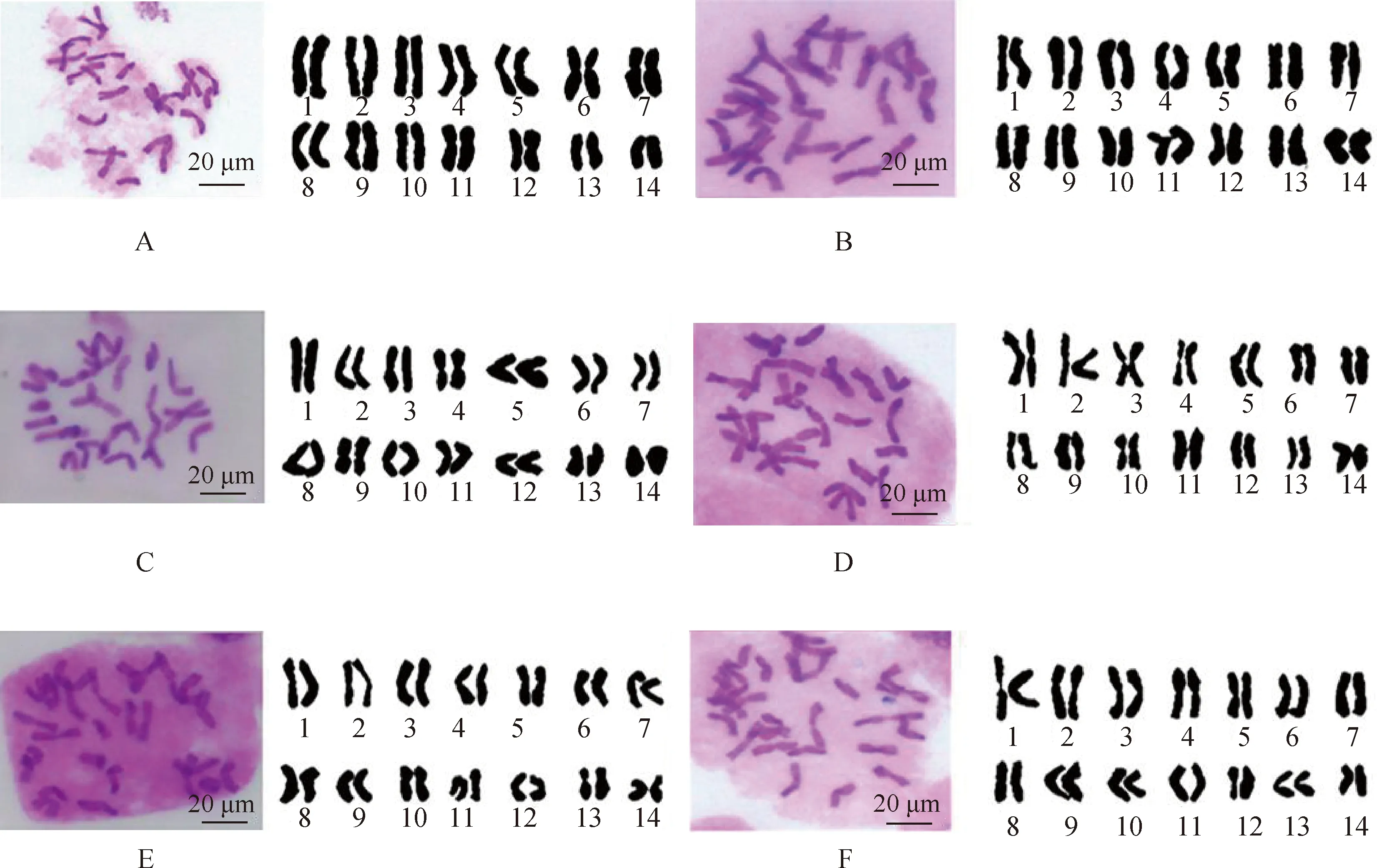
图1 扇穗茅有丝分裂中期染色体Fig.1 Mitotic chromosomes in metaphase of L. racemosa注:A,P119;B,P120;C,P067,D;P83;E,P84;F,P94。下同Note:A:P119;B,P120;C,P067;D,P83;E,P84;F,P94. The same as below
2.2 不同居群染色体形态特征与类型分析
从扇穗茅每个参试居群中,各选取10个分散良好、数目清晰、形态完整的染色体中期分裂图,依据每条染色体位置和参数进行同源染色体配对,共获得14对染色体,参数统计分析结果如表2所示。

表2 扇穗茅不同居群染色体参数Table 2 Chromosome parameters of different populations in L. racemosa
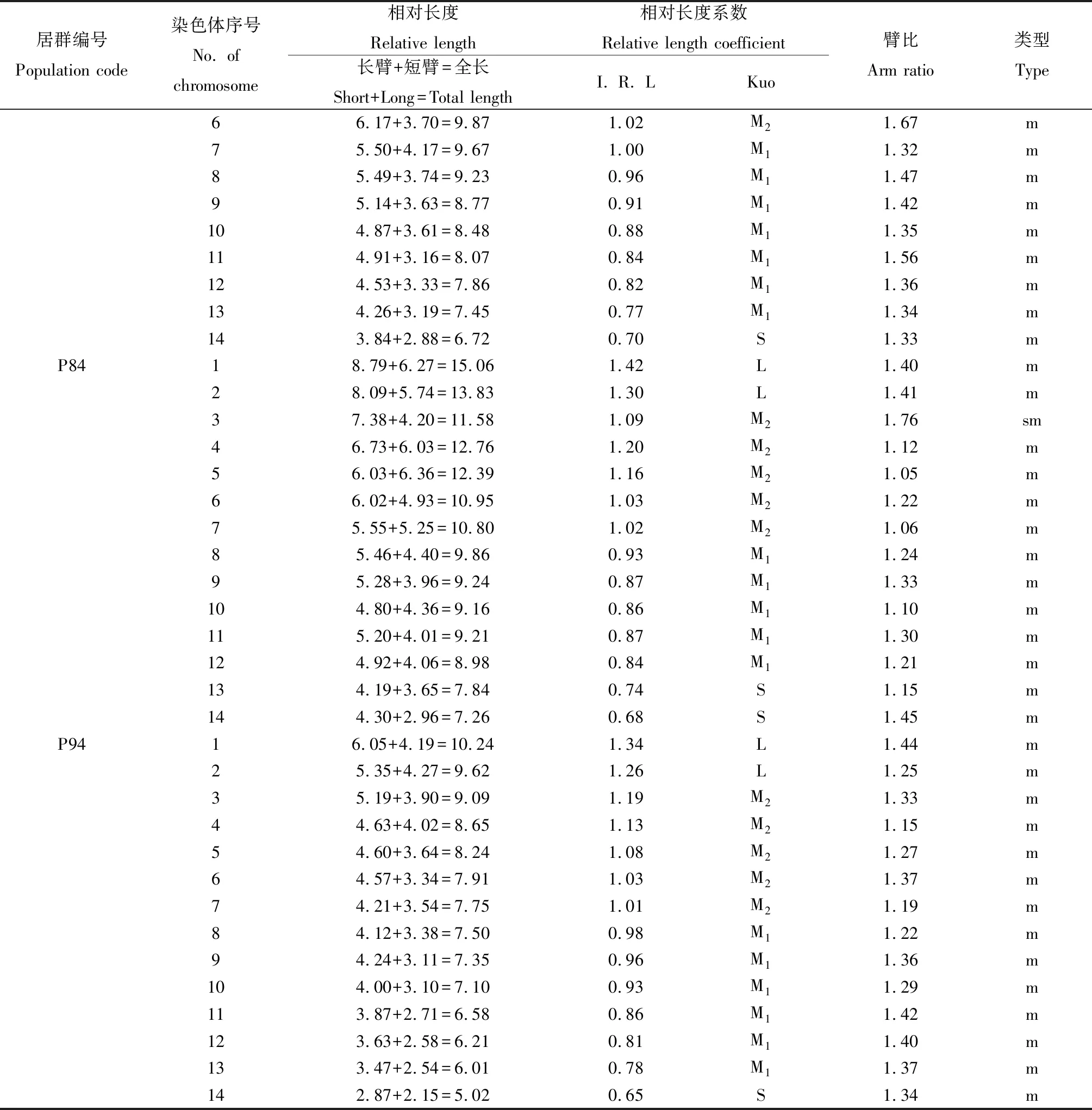
续表2
由表2可知,扇穗茅居群119和居群84的染色体为中部着丝点(m)和近中部着丝点(sm)染色体,居群120,67,83和94的染色体均为中部着丝点染色体,并且中部着丝点染色体占比较高;同时,依据染色体相对长度系数和Kuo分类标准,扇穗茅不同居群的染色体可以被划分为短染色体S型(<0.76)、中短染色体M1型(0.76~1.00)、中长染色体M2型(1.01~1.25)和长染色体L型(≥1.26)四种类型(表2)。
2.3 不同居群染色体核型分析
通过对扇穗茅不同居群中期分裂相染色体的统计分析,本研究获得了染色体长度比、臂比大于2的比率、核型公式与类型、核不对称系数等参数(表3);同时,根据各居群同源染色体参数绘制了核型模式图(图2)。

表3 扇穗茅不同居群的核型比较Table 3 Karyotype comparison of different populations in L.racemosa
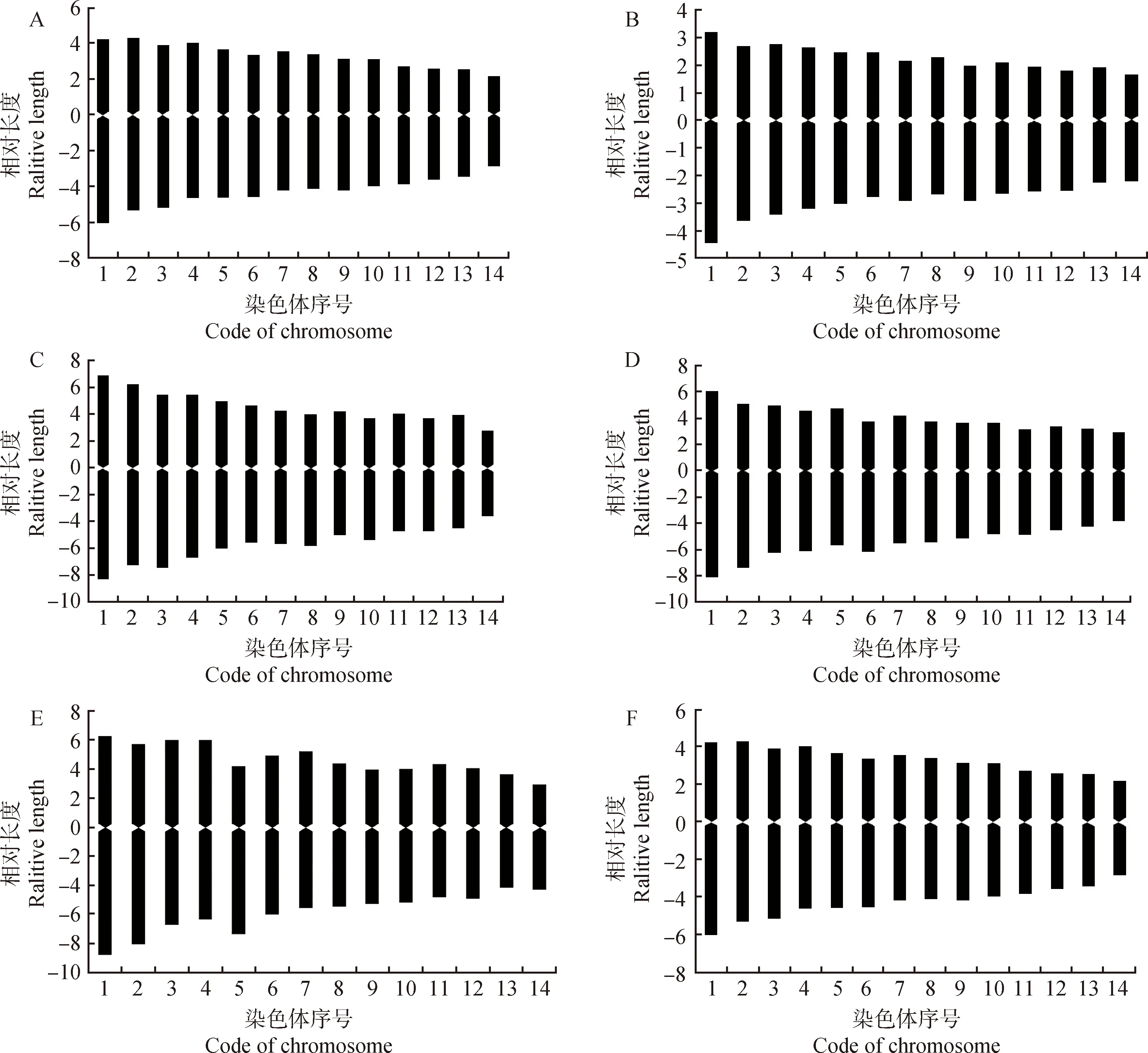
图2 扇穗茅不同居群的染色体核型模式图Fig.2 Chromosome idiograms of different populations in L. racemosa
由表3可知,扇穗茅6个居群的染色体核型类型有2A,1B和2B三种,其中居群120的染色体核型类型是2A型,居群67是1B型,其余4个居群是2B型;就扇穗茅染色体臂比而言,居群83的臂比范围为1.32~1.55(图2D),平均臂比最大(1.44),居群67和84的臂比范围分别为1.25~1.28(图2C)和1.19~1.35(图2E),平均臂比最小(1.27),其余3个居群的平均臂比介于两者之间(表3);同样,扇穗茅居群67的染色体核不对称系数最小,为55.67,而居群83的核不对称系数最大,为57.88,表明扇穗茅不同参试居群的染色体核型差异较大。
2.4 不同居群染色体核型进化趋势分析
以扇穗茅染色体的平均臂比为横坐标,以长度比和核不对称系数为纵坐标,分别绘制二维进化趋势图(图3)。结果表明,扇穗茅的染色体核型呈“双向进化趋势”,如居群83的染色体沿平均臂比方向进化较快,居群119的染色体则沿长度比方向进化较快(图3A);然而,就染色体总体进化趋势而言,居群83的进化程度较高,居群67的进化程度较低(图3A)。同时,通过对扇穗茅染色体平均臂比和核不对称系数的分析,我们发现扇穗茅染色体的总体进化趋势沿二维图右上角方向进化,即右上方的居群往往具有较高的进化程度,而左下方的居群进化程度相对较低。其中,居群83的平均臂比与核不对称系数最大,位于二维图右上方,进化程度较高;居群67的平均臂比与核不对称系数最小,位于二维图左下方,进化程度较低(图3B)。因此,本研究认为扇穗茅居群83的起源较晚,进化程度较高;居群67的起源较早,进化程度较低。

图3 扇穗茅不同居群染色体核型进化趋势Fig.3 Evolution trend of chromosome karyotypes in different populations of L. racemosa
2.5 不同居群染色体核型聚类分析
扇穗茅染色体核型的聚类分析结果显示,当遗传距离为25时,扇穗茅的6个参试居群聚为A和B两大分支,其中居群119,120,94和83聚为A支,居群67和84聚为B支,每个分支上的居群间亲缘关系相对较近,不同分支居群间则具有相对较远的亲缘关系;当遗传距离约为11时,扇穗茅的6个参试居群则形成3个分支,其中居群119,120和94聚为一支,居群83单独形成一支,居群67和84聚为一支,相比居群94,居群83与居群120和119则具有相对较远的亲缘关系;当遗传距离约为1时,居群119和120优先聚为一支,彼此间亲缘关系相对最近。

图4 扇穗茅不同居群染色体核型聚类分析Fig.4 Clustering analysis of chromosome karyotypes in different populations in L. racemosa
3 讨论
研究表明,染色体作为遗传物质的载体,其数目、形态和结构较为稳定,因而通过对植物染色体核型的综合分析,不仅可以探究不同物种以及同一物种不同居群间的系统进化关系,而且也可为该物种的细胞学研究提供科学依据[22-23]。通过对扇穗茅6个不同地理居群染色体核型的比较分析,本研究发现扇穗茅所有居群的染色体均为二倍体、数目为28条(图1),然而染色体的相对长度、长度比、平均臂比等在扇穗茅的不同居群间却存在一定差异。张建波等[24]基于川西北高原垂穗披碱草(Elymusnutans)12个居群染色体核型的研究,认为地理气候环境可以显著影响植物染色体核型的形态与结构,其也是造成染色体形态多样性的主要原因;王亚男等[12]通过对内蒙古高原沙鞭6个居群染色体核型特征和进化关系的分析,发现生长环境不同导致了沙鞭不同居群的染色体形态特征产生差异,并逐渐形成稳定的生态型[25]。本研究中,扇穗茅的6个参试居群来自不同的地域,其分布的经纬度和海拔高度存在差异(表1),据此我们认为扇穗茅6个参试居群的染色体形态差异也是由其地域分布和生态环境不同造成的,这与先前的研究结论是一致的[12,24-25]。
扇穗茅6个参试居群的染色体均为中部着丝点(m)和近中部着丝点(sm)染色体,其中中部着丝点染色体比例较高,这与先前报道的近缘种无芒雀麦(Bromusinermis)[26]和沙地雀麦(B.ircutensis)[27]的染色体核型类型相同。同时,通过对扇穗茅6个参试居群染色体核型的比较分析,本研究发现扇穗茅的染色体核型类型主要有2A,1B和2B三种,其中居群120是2A型、居群67是1B型、其余4个居群均为2B型(表3),表明它们均具有较好的对称性,与先前报道的无芒雀麦[28]、沙地雀麦[27]和扁穗雀麦(B.catharticus)[29]等的研究结果类似。Stebbins[19]认为,植物染色体核型由对称性向不对称性进化,对称性越高染色体变异越小,进化程度越低;反之,非对称性越高染色体变异越大,进化程度越高。因此,我们推测雀麦族植物可能是一个进化上较为保守的类群,进化水平相对较低;染色体形态和结构差异可能是由其生长环境不同导致的。此外,植物进化程度与其环境适应性密切相关[30],通常对环境适应性越强的植物其进化程度往往越高[31]。据此,我们推测扇穗茅的居群83理应对环境具有较强的适应性,未来遗传育种上可能具有潜在价值。
聚类分析可以利用样本的多变量信息将其划分为相对同质的集群,从而清晰地展现不同集群间的系统亲缘关系[32]。本研究结果显示,当遗传距离为25时,扇穗茅6个参试居群聚为A和B两大分支,其中居群119,120,94和83聚为A支,居群67和84聚为B支,同一分支上不同居群间具有相对较近的亲缘关系;当遗传距离约为1时,居群119和120优先聚类,彼此间亲缘关系最近。同时,我们还发现分支A的4个居群分布于海拔相对较高的区域,而分支B的两个居群分布的海拔高度相对较低。研究表明,随着海拔高度的升高温度会逐渐降低,并且降水、地表温度及升温速率等环境因子也会随之发生改变[33],因而我们认为扇穗茅6个参试居群的亲缘关系可能与其分布的海拔高度和生境存在必然联系。本研究首次报道了扇穗茅的染色体数目、核型特征及进化趋势,为将来进一步探究扇穗茅属乃至雀麦族植物的系统进化关系提供了有力的细胞学佐证。
4 结论
综上所述,扇穗茅的染色体为二倍体,数目恒定,为2n=2x=28;染色体有中部着丝粒(m)型和亚中部着丝粒(sm)型染色体,其中中部着丝粒型染色体占比较高,包含2A,1B和2B三种类型;扇穗茅不同参试居群的染色体核型臂比和平均臂比差异较大,呈“双向进化趋势”;扇穗茅染色体总体沿二维图右上角方向进化,即右上方居群具有较高的进化程度,左下方居群进化程度相对较低;聚类分析显示,当遗传距离为25时,扇穗茅6个参试居群聚为A和B两大分支,每个分支上的居群间亲缘关系相对较近,不同分支居群间亲缘关系相对较远。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
生物技术进展(2022年1期)2022-02-11 03:45:46
Rice Science(2021年4期)2021-07-13 10:00:54
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20 02:09:54
科学之谜(2018年9期)2018-12-17 09:05:08
草地学报(2018年5期)2018-11-07 02:25:00
第二课堂(小学版)(2018年6期)2018-07-09 01:06:46
快乐语文(2016年10期)2016-11-07 09:44:51
生物学教学(2016年12期)2016-04-10 23:04:28
