
不同品系绒山羊皮肤毛囊生长发育周期中chi-miR-107-3p及其相关基因的表达分析
2022-07-25 06:39王立忠吴铁成马跃军赵若阳高玉林闫新刚李玉荣
中国农业大学学报 2022年7期
刘 斌 王立忠 吴铁成,2 马跃军,3 赵若阳 高玉林 闫新刚 李玉荣*
(1.内蒙古自治区农牧业科学院,畜牧研究所,呼和浩特 010031;2.肉羊遗传资源评价与繁育技术内蒙古自治区工程实验室,呼和浩特 010031;3.阿拉善白绒山羊种羊场,内蒙古 阿拉善 750336;4.鄂尔多斯市立新实业有限公司,内蒙古 鄂尔多斯 017000)
绒山羊是重要的绒用经济动物,其绒毛生长具有较强的季节性,但不同品种之间有很大的差异,大多数绒山羊在非长绒期(5~7月)不长绒,次级毛囊生长发育分为休止期(3~4月)、兴盛前期(5~7月)、兴盛期(8~12月)和退行期(1~2月)等4个阶段,而有少数绒山羊可以常年长绒(6月开始长绒)。内蒙古白绒山羊是经过长期选育而形成的绒肉兼用型地方良种,分为巴彦淖尔市的二狼山型、鄂尔多斯市的阿尔巴斯型和阿拉善盟的阿拉善型三个类群,具有季节性长绒,羊绒细,纤维长,光泽好,绒毛手感柔软,综合品质优良的特性。鄂尔多斯市伊金霍洛旗苏布尔嘎培育的敏盖绒山羊(未经品种命名),全舍饲养殖,具有常年长绒、产绒量高等特点,成为当地牧民主要的养殖家畜。2016年“敏盖”商标被认定为鄂尔多斯市知名商标,2019年被国家知识产权局认定为驰名商标,因此暂命名为“敏盖绒山羊”。绒山羊绒毛生长受遗传、营养和光照、温度等环境因素的影响,但遗传占主要影响因素,从基因调控的方向研究皮肤毛囊生长发育显得尤为重要。miRNA是真核生物中存在的小分子非编码RNA家族,具有抑制靶向mRNA转录、翻译或者能够剪切靶向mRNA并促进其降解的功能。
近年来,国内外许多学者研究报道miRNA在毛囊形态发生、毛囊干细胞的增殖、分化和凋亡的稳态平衡过程中发挥着重要作用。Yuan等发现陕北绒山羊、内蒙古绒山羊不同时期的皮肤毛囊miRNA表达存在特异性。Yuan等发现miR-22可以促进毛囊退化,维持毛囊处于静止期。马涛发现miRNA-203在绒山羊毛囊发育的生长期和休止期均有表达,且在休止期的表达量显著高于生长期,其候选靶基因的表达则相反。吴志红通过实时荧光定量PCR和Western blot研究表明在胎儿皮肤成纤维细胞和上皮细胞中,chi-miR-130b-3p、chi-miR-301a-3p 和 chi-miR-532-3p 在mRNA和蛋白水平上均可不同程度的调控WNT10A的表达。chi-miR-107-3p是一类小分子RNA,Wu等通过高通量测序技术在萨能奶山羊睾丸组织中发现。Li等研究发现应用纳米载体(TLN)可使chi-miR-107-3p缺失,dicer表达上调,从而加速角质形成细胞的分化,从而促进皮肤伤口的愈合。尽管有众多的miRNA参与绒山羊皮肤毛囊周期性生长过程,但鲜少有chi-miR-107-3p在不同品系间绒山羊毛囊生长发育的比较研究。本研究基于课题组前期对阿尔巴斯型绒山羊光控增绒处理组和对照组皮肤毛囊样品进行全转录组测序的结果(相关数据尚未发表),筛选到chi-miR-107-3p及其相关基因ADCK5
、DSBC1
、ARSA
、FMAP4
、RHBDF2
和ALDH3A2
,可能与绒山羊皮肤毛囊的周期性生长发育有关。因此,以绒毛生长周期差异显著的阿拉善型白绒山羊和敏盖绒山羊为研究对象,应用组织切片技术对不同品系绒山羊处于不同生长周期毛囊结构进行比较分析,并通过qPCR方法对chi-miR-107-3p和相关基因的表达规律进行探究,旨在为绒毛生长周期变化分子基础提供理论参考。1 材料与方法
1.1 试验材料
选取内蒙古阿拉善白绒山羊种羊场和鄂尔多斯敏盖绒山羊羊场体重相近的2岁绒山羊母羊各3只,分别在1、4、7和9月采集体侧部位直径1 cm的皮肤2份,一份用4%多聚甲醛固定进行组织切片制作,另一份快速保存于液氮罐中,用于qRT-PCR检测。采样前在取样区域周围皮下注射2%利多卡因进行局部麻醉,采样过程符合动物福利要求。
1.2 试剂及仪器设备
主要试剂:伊红染液、苏木素染液等购自北京索莱宝科技有限公司;4%多聚甲醛、石蜡、TRIZOL、Tris碱、乙二胺四乙酸二钠(EDTA)、AL 2000、琼脂糖、RNase-Free dd HO、氯仿、75%冰乙醇、2×Phanta Max Master Mix、1×TAE缓冲液;PrimeScript(TM) RT Master Mix (Perfect Real Time)、TB Green(TM) Premix Ex Taq(TM) II (RNaseH Plus)、Mir-X miRNA First-Strand Synthesis和TB Green qRT-PCR User Manual等反转录试剂盒、荧光定量试剂盒等购自TaKaRa。
主要仪器设备:LEICA RM2016切片机、梯度PCR仪(Eppendorf,德国)、荧光定量PCR仪(CFX 96 Bio-Rad,美国)、正倒置一体荧光显微镜。
1.3 试验方法
1
.3
.1
皮肤组织HE染色将皮肤样品在4%多聚甲醛中固定24 h后取出,梯度酒精脱水,二甲苯透明,石蜡包埋,切成7 μm的薄片,用常规HE染色方法染色,中性树脂封片,干燥后观察。由于皮肤在脱水透明过程中会发生收缩现象,因此在研究皮肤毛囊形态时需对皮肤收缩率进行校正。
皮肤收缩率=脱水透明后皮肤面积/
活体样皮肤面积×100%
表皮厚:真皮层到皮肤表面的垂直距离,μm;
真皮厚:皮下脂肪组织到表皮层的垂直距离,μm;
初、次级毛球宽:用显微镜微测尺选择不同视野10个完整的初、次级毛囊,测量其毛球的宽度,μm;
初、次级毛囊深:初、次级毛囊到表皮层的垂直距离,μm;
初、次级毛囊密度:利用显微镜微测尺选择视野内10个完整初、次级毛囊个数,再进行换算为个/mm。
1
.3
.2
实时荧光定量法1.3.2.1 引物设计 chi-miR-107-3p序列为AGCAGCAUUGUACAGGGCUAU,U6
作为内参基因。使用软件Primer 5.0设计RHBDF2
、ADCK5
、ARSA
、ALDH3A2
、DSBC1
和FMAP4
基因荧光定量引物,选取GAPDH
为内参基因。引物由生工生物工程(上海)股份有限公司(Sangon Biotech (Shanghai)Co., Ltd.)合成(表1)。表1 引物信息
Table 1 Primer information
基因名称Gene name登录号Login number引物序列(5'→3')Primer sequence (5'→3')片段大小/bpProduct length退火温度/℃Annealingtemperaturechi-miR-107-3pNC_030833.1AGCAGCATTGTACAGGGCTAT10656 ℃RHBDF2XM_018063741.1F: GTCCCAGAGCATCCGCAAGTR: TCCACGCCTCCCACAGAA13158 ℃ADCK5XM_018058741.1F: CGCTCCTCTTCCCTCCTTR: CCACCCATTCTGGCAACT14864 ℃ARSAXM_005701596.3F: AGAGCTTTCCAGGGCACTCAR: CATCGTCTCAGGTCCGTTGT15562 ℃ALDH3A2XM_018064739.1F: ATTTACTCCAGCCTGCCTACR: CAGCCAAGTGTCACCTCA12662 ℃DSBC1XM_005676701.3F: GCAGGCTCTGACCGAATCR: CTGGGCTGGTGACAAGGA8858 ℃FMAP4XM_018064747.1F: TCCACCTTTGACCGAGACCR: GTTGATGCCATTGGCGTAG14758 ℃U6NM_001161346.2F: GGAACGATACAGAGAAGATTAGCR: TGGAACGCTTCACGAATTTGCG6862 ℃GAPDHXM_005680968.3F: CCACGAGAAGTATAACAACACCR: GTCATAAGTCCCTCCACGAT12060 ℃
1.3.2.2 总RNA提取及cDNA合成 用液氮将皮肤样品研磨后,使用TRIZOL提取总RNA,采用紫外分光光度计法检测总RNA浓度,并用琼脂糖凝胶电泳和A/A检测RNA质量,保存于-80 ℃冰箱中备用。采用PrimeScript(TM) RT Master Mix (Perfect Real Time)试剂盒合成cDNA(mRNA),采用Mir-X miRNA First-Strand Synthesis and TB Green qRT-PCR User Manual试剂盒合成cDNA(miRNA),保存于-20 ℃备用。
1.3.2.3 qRT-PCR检测 利用SYBR Green实时荧光定量PCR方法,以GAPDH
基因为内参,对mRNA进行荧光定量;以U
6为内参,对chi-miR-107-3p进行荧光定量。反应体系为TB Green(TM) Premix Ex Taq(TM) II (RNaseH Plus) 10 μL,Primer F 1 μL,Primer R 1 μL,cDNA 1 μL,RNase-Free ddHO 7 μL。反应程序为95 ℃预变性2 min,(95 ℃ 变性10 s,62 ℃退火10 s,72 ℃延伸 20 s)40个循环后进入溶解曲线,55 ℃到95 ℃,每0.6 s上升0.5 ℃。1.4 数据分析
用2值法对chi-miR-107-3p及mRNA的表达量进行计算。运用SPSS 20.0对所得数据进行相关性分析,使用双变量Pearson进行检测,实验数据均以“平均值±标准差”表示,以P
<0.05为差异显著,P
<0.01为差异极显著。2 结果与分析
2.1 不同品系绒山羊皮肤毛囊组织显微结构分析
校准并测量皮肤收缩率发现,2个品系绒山羊皮肤收缩率均约为65%。皮肤毛囊组织显微结构观察分析发现2个品系毛囊群结构均以三元型为主(3个初级毛囊和若干个次级毛囊),可见少量的二元型和四元型。在退行期,次级毛囊细胞核增大,开始死亡,同时有部分细胞开始向外扩散,呈现刷状分布,细胞破碎,逐渐成为一点甚至消失,毛囊群之间的结缔组织较薄,连接不紧密(图1(a)和(b),图2(a)和(b));在休止期,次级毛囊基本退化,极少看到具有红色内跟鞘的毛囊,毛乳头完全消失,次级毛囊密度达到最低,毛囊群结构之间的结缔组织变薄,次级毛囊排布散乱,大小不一,多成散状分布(图1(c)和(d),图2(c)和(d));兴盛前期,级毛囊再次重建,初级毛囊周围出现多个小毛囊,同时也有几个小毛囊紧密相靠,这说明有新的毛囊产生。如图1(e)、图2(g)、图2(e)和图2(g),具有常年长绒特点的敏盖绒山羊次级毛囊重建均已基本完成,初、次级毛囊均呈椭圆形,排列紧密、整齐。而阿拉善型绒山羊次级毛囊正处于重建阶段,次级毛囊分布杂乱,仍有部分新生毛囊出现;在兴盛期,次级毛囊重建完成,此时次级毛囊密度最大,具有红色内跟鞘的毛囊也最多,次级毛囊排布整齐,毛囊群之间的结缔组织连接紧密,结构明显。如图1(f)、图2(h)、图2(f)和2(h)。结合表2可见,2个品系绒山羊初级毛囊密度在整个生长周期内几乎没有变化,而次级毛囊密度变化显著。皮肤毛囊兴盛前期,阿拉善型绒山羊次级毛囊密度极显著低于敏盖绒山羊(P
<0.01),次级毛囊深、初级毛球宽、次级毛球宽和S/P值(次级毛囊数/初级毛囊数)等性状均显著小于敏盖绒山羊(光控增绒)(P
<0.05)。在皮肤毛囊兴盛期,阿拉善型绒山羊次级毛囊密度、次级毛囊深和次级毛球宽显著低于敏盖绒山羊(P
<0.05)。表明两种品系的绒山羊初级、次级毛囊在形态特征上基本一致,但在同一时期的生长规律不同。表2 不同品系内蒙古绒山羊皮肤毛囊组织结构统计分析
Table 2 Statistical analysis of transverse skin and hair follicle tissue structure of different strains of Inner Mongolia cashmere goats
性状Character月份Month阿拉善型绒山羊Alashan cashmere goats敏盖绒山羊Mingai cashmere goats1月26.29±0.18 a27.13±1.90 a表皮厚度/μmDepth of secondary follicle4月20.24±1.58 a21.31±0.64 a7月27.51±3.48 a34.19±3.72 a9月32.36±1.64 a34.16±1.43 a1月1 015.94±14.34 a1 100.04±9.56 a真皮厚度/μmThickness of dermis4月1 012.42±61.44 a976.52±37.76 a7月1 091.89±19.50 a1 126.77±37.63 a9月1 093.35±10.38 a1 192.60±10.75 a1月2 015.54±20.19 a2 041.60±42.67 a初级毛囊深度/μmDepth of primary follicle4月1 707.83±24.75 a1 733.48±25.48 a7月2 073.33±7.51 Bc2 281.1±49.29 ABb9月2 394.65±34.89 a2 488.45±65.85 a1月1 162.54±16.75 a1 151.79±5.64 a次级毛囊深度/μmDepth of secondary follicle4月1 008.70±20.52 a1 070.24±2 977 a7月1 155.50±31.09 b1 276.27±19.01 a9月1290.91±29.86 b1 456.91±76.32 a1月224.44±7.85 a252.48±8.38 a初级毛球直径/μmDiameter of primary hair bulb4月157.77±6.40 a181.36±13.77 a7月239.30±19.64 b305.57±7.59 a9月317.54±8.07 a313.70±10.74 a1月96.14±1.40 a101.71±3.37 a次级毛球直径/μmDiameter of secondary hair bulb4月84.53±2.93 a91.53±9.70 a7月84.30±0.91 b112.56±5.31 a9月101.92±3.30 b118.73±5.69 a1月2.91±0.79 a2.63±0.76 a初级毛囊密度/(个/mm2)Density of primary follicle4月2.45±0.61 a2.54±0.57 a7月2.99±0.71 a2.90±0.54 a9月2.99±0.75 a3.00±0.75 a1月26.27±2.13 a27.18±3.36 a次级毛囊密度/(个/mm2)Density of secondary follicle4月22.54±3.11 a23.09±3.59 a7月28.36±2.29 A35.63±2.98 B9月39.09±4.04 a43.91±3.86 b1月9.61±2.40 a10.83±2.14 a初级毛囊数/次级毛囊数S/P4月9.49±1.67 a9.37±1.95 a7月10.41±2.72 a12.54±1.67 b9月13.85±2.64 a15.20±2.30 a
注:同行数据肩标不同小写字母表示差异显著(<0.05);肩标不同大写字母表示差异极显著(<0.01);肩标相同字母表示差异不显著(>0.05)。
Note: In the same line, values with different small letters superscripts mean significant difference (<0.05); Different capital letters superscripts mean extremely significant difference (<0.01); While the same letter superscripts mean no significant difference (>0.05).
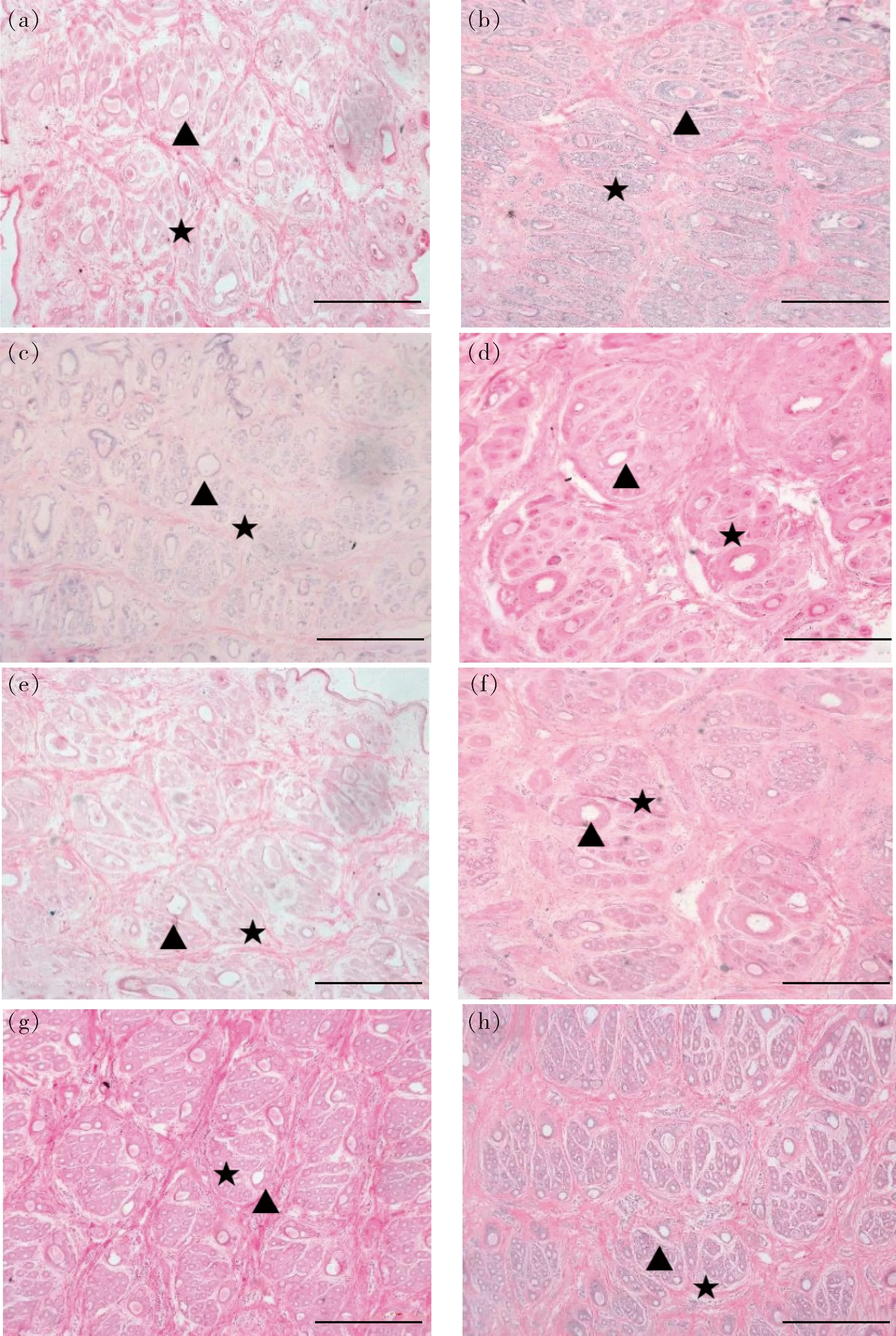
(a)、(c)、(e)、(g)分别是阿拉善型绒山羊皮肤毛囊退行期、休止期、兴盛前期、兴盛期横切切片;(b)、(d)、(f)、(h)分别是敏盖绒山羊皮肤毛囊退行期、休止期、兴盛前期、兴盛期横切切片;▲初级毛囊;★次级毛囊。比例尺为100 μm。(a), (c), (e) and (g) were transverse sections of the skin follicles of Alxa cashmere goats in anagen, resting stage, prosperous and regression stage respectively; (b), (d), (f) and (h) were transverse sections of the skin follicles of Mingai cashmere goats in anagen, resting stage, prosperous and regression stage respectively; ▲ Primary hair follicle; ★ Secondary hair follicle. Bar is 100 μm.图1 不同品系绒山羊皮肤毛囊横切切片Fig.1 Transverse skin sections of different strains on cashmere goats

(a)、(c)、(e)、(g) 分别是阿拉善型绒山羊皮肤毛囊退行期、休止期、兴盛前期、兴盛期横切切片;(b)、(d)、(f)、(h) 分别是敏盖绒山羊皮肤毛囊退行期、休止期、兴盛前期、兴盛期横切切片;▲初级毛囊;★次级毛囊。比例尺为100 μm。(a), (c), (e) and (g) were localisation sections of the skin follicles of Alxa cashmere goats in anagen, resting stage, prosperous and regression stage respectively; (b), (d), (f) and (h) were localisation sections of the skin follicles of Mingai cashmere goats in anagen, resting stage, prosperous and regression stage respectively; ▲ Primary hair follicle; ★ Secondary hair follicle. Bar is 100 μm.图2 不同品系绒山羊皮肤毛囊纵切切片Fig.2 Localisation skin sections of different strains on cashmere goat
2.2 不同品系绒山羊皮肤组织chi-miR-107-3p相关基因在不同毛囊周期的表达分析
2
.2
.1
不同品系绒山羊皮肤组织中chi-miR-107-3p的表达分析由图3(a)可见,阿拉善型绒山羊皮肤毛囊兴盛前期chi-miR-107-3p达量极显著高于敏盖绒山羊(P
<0.01)。对同一品种不同时期chi-miR-107-3p的表达量进行分析,由图3(b)可见,阿拉善型绒山羊皮肤组织中,chi-miR-107-3p表达量在兴盛前期极显著大于兴盛期(P
<0.01),且显著大于退行期和休止期(P
<0.05)。敏盖绒山羊皮肤组织中,chi-miR-107-3p的表达量在休止期最高,且显著大于其他3个时期(P
<0.05)。因此说明,不同品系绒山羊皮肤组织中chi-miR-107-3p的表达规律不同。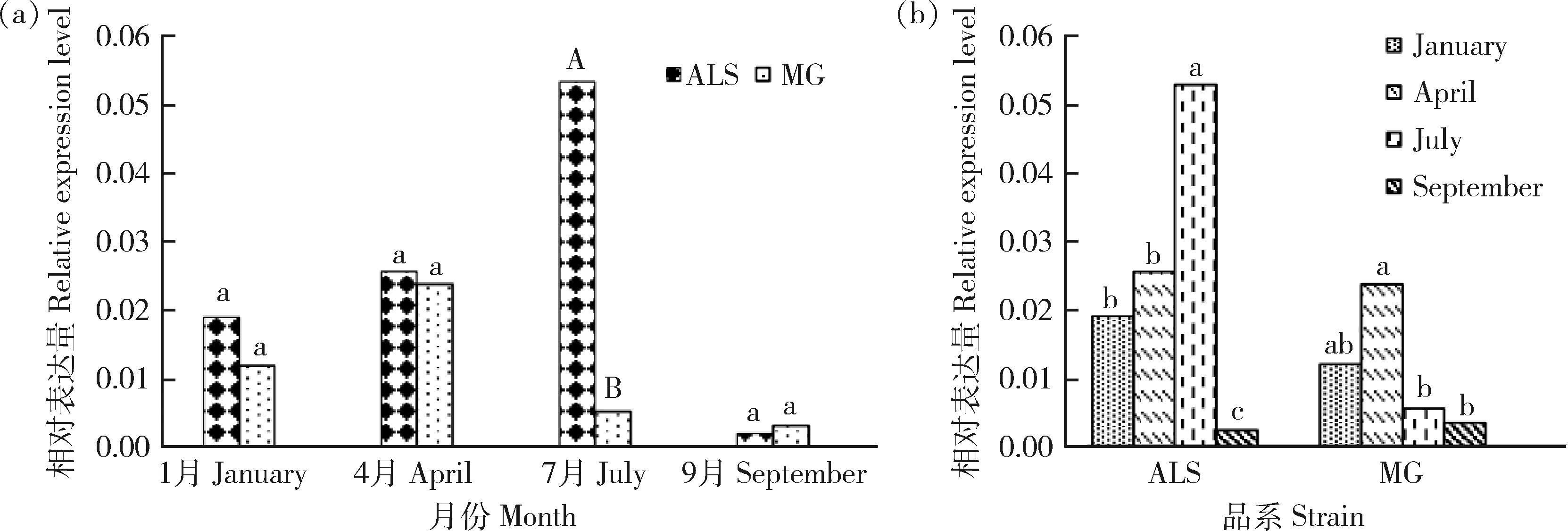
ALS:阿拉善型绒山羊; MG:敏盖绒山羊; January:绒山羊皮肤毛囊退行期; April:绒山羊皮肤毛囊休止期; July: 绒山羊皮肤毛囊兴盛前期;September:绒山羊皮肤毛囊兴盛期。图3a为相同时期不同品系之间的比较,图3b为相同品系不同时期的比较,组间比较肩标不同小写字母表示差异显著(P<0.05);肩标不同大写字母表示差异极显著(P<0.01);肩标相同字母表示差异不显著(P>0.05);下同。ALS: Alxa cashmere goat; MG: Mingai cashmere goat; January: the skin follicles of Alxa cashmere goats in anagen; April: the skin follicles of Alxa cashmere goats in resting stage; July: the skin follicles of Alxa cashmere goats in prosperous; September: the skin follicles of Alxa cashmere goats in regression stage respectively. Different small letters superscripts mean significant difference (P<0.05); Different capital letters superscripts mean extremely significant difference (P<0.01); While the same letter superscripts mean no significant difference (P<0.05); The same as below.图3 不同品系绒山羊皮肤毛囊不同生长时期chi-miR-107-3p相对表达量Fig.3 Relative expression of chi-miR-107-3p in skin and hair follicles of different strains Cashmere goats
2
.2
.2
不同品系绒山羊皮肤组织中chi-miR-107-3p相关基因在不同毛囊周期的表达量分析对不同品系绒山羊皮肤毛囊不同发育时期皮肤组织中ADCK
、DSBC1
、RHBDF2
、ALDH3A2
、ARSA
和FMAP4
基因的表达量进行差异性分析(图4)。结果显示,在休止期(4月),阿拉善型绒山羊皮肤组织中ADCK
5基因的表达量极显著大于敏盖绒山羊(P
<0.01)。兴盛前期(7月),阿拉善型绒山羊皮肤组织中RHBDF2
基因极显著低于敏盖绒山羊(P
<0.01)。兴盛期(9月),阿拉善型绒山羊和敏盖绒山羊皮肤组织中ADCK5
、DSBC1
、RHBDF2
、ALDH3A2
、ARSA
和FMAP4
基因的表达量差异不显著(P
>0.05)。退行期(1月),阿拉善型绒山羊皮肤组织中DSBC1
基因的表达量极显著低于敏盖绒山羊(P
<0.01)。在整个皮肤毛囊的生长发育周期中,RHBDF2
基因的表达量在兴盛前期到兴盛期的过程中逐渐升高,在兴盛期达到最高,之后逐渐降低,到达退行期,其表达量最低;ALDH3A2
基因的表达量在休止期最高,随着皮肤毛囊的生长发育,其表达量逐渐降低,到达兴盛期,其表达量最低,到休止期表达量逐渐升高。由此可见,不同品系绒山羊皮肤组织中chi-miR-107-3p相关基因在同一毛囊发育阶段表达量不一致。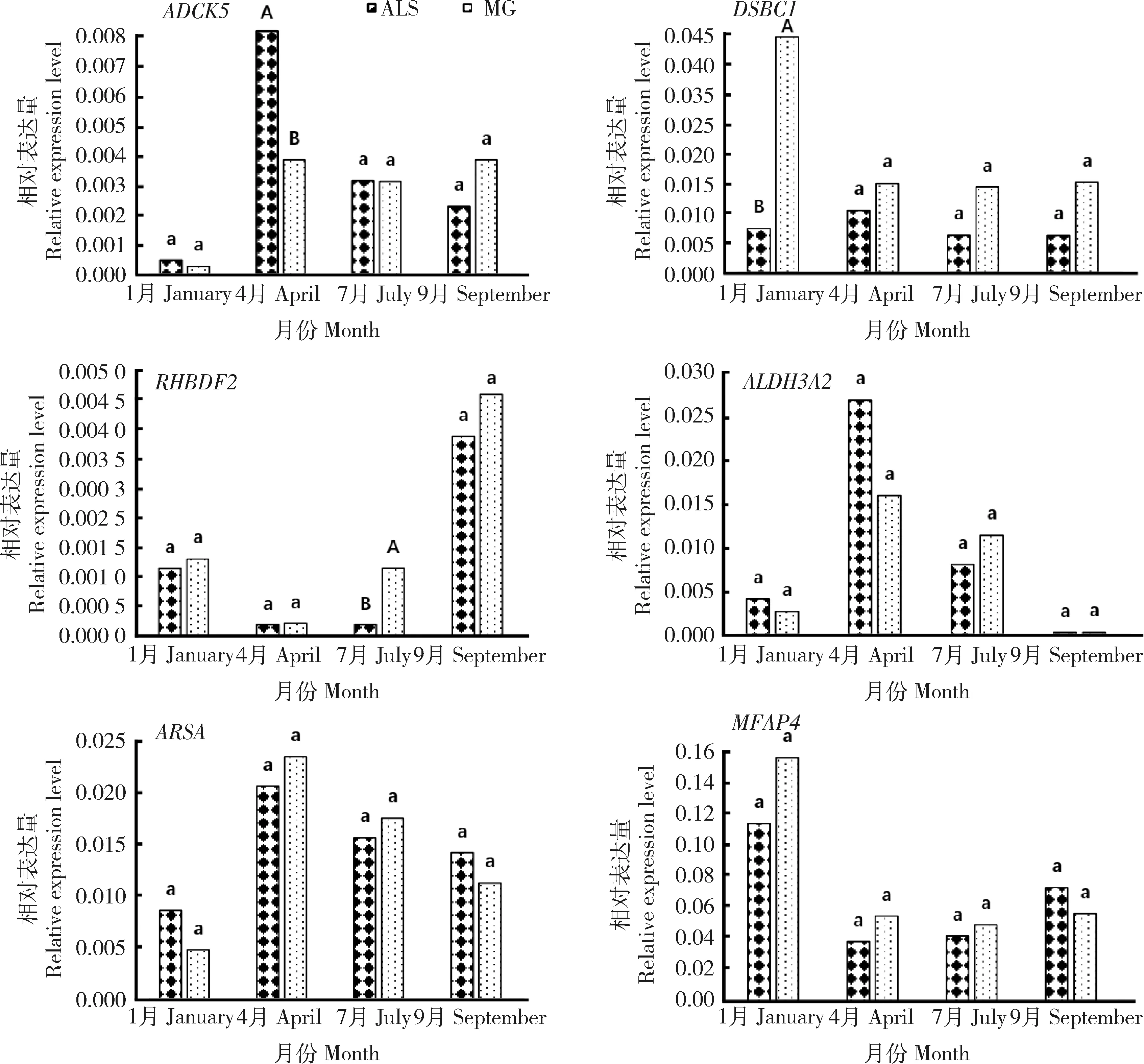
图4 不同品系绒山羊皮肤毛囊不同生长时期mRNA的相对表达量Fig.4 Relative expression of mRNA in skin and hair follicles of different breeds(strains) Cashmere goats
2
.2
.3
chi-miR-107-3p及其相关基因和皮肤及毛囊性状间的相关性分析对chi-miR-107-3p以及6个基因和两品系绒山羊皮肤毛囊性状数据进行相关性分析(表3),chi-miR-107-3p和RHBDF2
基因与皮肤毛囊初级毛囊密度和真皮厚显著相关(P
<0.05),与次级毛囊密度、S/P、表皮厚、初次级毛囊深及初次级毛球宽均极显著相关(P
<0.01);ALDH3A2
基因与初级毛囊密度、真皮厚和次级毛球宽显著相关(P
<0.05),与次级毛囊密度、S/P、表皮厚、初次级毛囊深及初级毛球宽均极显著相关(P
<0.01);DSBC1
基因仅与初级毛囊密度显著相关(P
<0.05),其他基因与皮肤毛囊性状均未表现相关性。表明差异表达的chi-miR-107-3p及其相关基因并不完全与皮肤毛囊性状相关。表3 chi-miR-107-3p和相关基因与绒毛性状的关联分析
Table 3 The association analysis of chi-miR-107-3p and mRNA with fleece traits
性状Characterchi-miR-107-3pADCK5DSBC1RHBDF2ALDH3A2ARSAFMAP4初级毛囊密度Density of primary follicle0.0390.9550.0120.0380.0100.0870.606次级毛囊密度Density of secondary follicle0.0010.7480.0840.0010.0050.2140.848初级毛囊数/次级毛囊数S/P0.0010.6900.1640.0010.0060.2630.994真皮厚度Thickness of dermis0.0110.7350.5610.0110.0450.1290.891表皮厚度Thickness of epidermis0.0030.5110.1290.0030.0090.1490.930初级毛囊深度Depth of primary follicle0.0010.5880.2120.0010.0020.046*0.941次级毛囊深度Depth of secondary follicle0.0010.2560.1080.0010.0030.2260.756初级毛球直径Diameter of primary hair bulb0.0010.2680.1900.0010.0040.1690.926次级毛球直径Diameter of secondary hair bulb0.0010.2090.4550.0010.0100.1570.581
注:表中数据表示相关性值。
Note: The data in the table represent correlation values.
3 讨 论
3.1 不同品系绒山羊皮肤组织不同毛囊发育时期的形态结构分析
产绒量是绒山羊最主要的经济性状,次级毛囊结构特点和兴盛期的长短很大程度上影响产绒量绒毛品质,其中遗传力较高的3个性状分别是毛囊深度、毛囊密度以及S/P值,且均与产绒量相关。对绒山羊皮肤组织显微结构进行分析,发现初级毛囊密度虽然伴随着毛囊的退化、更新有一定幅度的变化,但是其毛囊密度在各生长阶段基本不发生变化。李长青等在对内蒙古绒山羊与辽宁绒山羊皮肤毛囊生长规律的研究中也得到相同的结论。由表2可见,阿拉善型绒山羊皮肤毛囊S/P值在兴盛前期显著低于敏盖绒山羊(P
<0.05),S/P值越大,说明皮肤毛囊的生长发育越完善。范一星在不同毛被类型内蒙古绒山羊皮肤毛囊周期性生长及相关分子机理研究中指出,活性S/P值较大时,表明次级毛囊生长发育更完善,与本研究结构相似。由于敏盖绒山羊属于常年长绒型绒山羊,次级毛囊生长发育较早,皮肤毛囊兴盛前期大部分毛干已经长出体表,次级毛囊毛球结构完整,而阿拉善型绒山羊此时期次级毛囊正处于重建过程当中,大部分毛囊发育仍不完整。在皮肤毛囊兴盛期,阿拉善型绒山羊次级毛囊密度显著低于敏盖绒山羊(P
<0.05),阿拉善型绒山羊次级毛囊深显著小于敏盖绒山羊(P
<0.05),且次级毛球宽显著小于敏盖绒山羊(P
<0.05),原因是阿拉善型绒山羊属于季节性长绒绒山羊,次级毛囊进入兴盛期较迟。赵艳丽等对辽宁绒山羊常年长绒型和季节性长绒型绒山羊皮肤毛囊组织结构研究指出,由兴盛前期转入兴盛期的过程中,常年型次级毛囊深、次级毛球宽均显著大于季节型,这与本研究一致。3.2 不同品系绒山羊皮肤组织中chi-miR-107-3p在不同毛囊发育时期的的表达分析
miRNA是最新发现在绒山羊皮肤毛囊生长发育过程中起调控作用的一类小分子非编码RNA,何玉龙等对不同发育阶段小尾寒羊皮肤毛囊特异性miRNA鉴定分析,为从miRNA层面进一步挖掘改良羊毛品质及产量相关功能基因方面提供数据支撑与研究思路。本研究发现,皮肤毛囊兴盛前期,chi-miR-107-3p在阿拉善型绒山羊皮肤组织中的表达量极显著高于敏盖绒山羊(P
<0.01)。同时结合皮肤组织显微结构分析,阿拉善型绒山羊次级毛囊正处于重建阶段,而敏盖绒山羊次级毛囊已经重建完成。这说明在皮肤组织中高表达的chi-miR-107-3p抑制次级毛囊生长发育。在兴盛期,chi-miR-107-3p在2个品系绒山羊皮肤组织中的表达量均较低,此时皮肤毛囊的生长发育最快。进入退行期,chi-miR-107-3p的表达量逐渐增大,此时次级毛囊正处于退化阶段。同时,经过chi-miR-107-3p与皮肤毛囊性状进行相关性分析发现,chi-miR-107-3p与皮肤毛囊各性状均显著相关(P
<0.05)。Li等研究发现,应用TLN可使chi-miR-107-3p缺失,dicer表达上调,可以加速皮肤伤口的愈合,说明chi-miR-107-3p的缺失可以加速角质形成细胞的分化,从而加速皮肤的生长。由此推测chi-miR-107-3p是绒山羊皮肤毛囊周期性生长发育的重要影响因子之一,可能参与次级毛囊重建。3.3 不同品系绒山羊皮肤组织RHBDF2和ALDH3A2基因在不同毛囊发育时期的表达分析
李晓凯利用毛被类型的名义显著性候选基因和不同毛被类型不同毛囊周期比较转录组数据,进行WGCNA数据的挖掘分析,发现RHBDF2
基因可能与不同毛被类型存在潜在一定的相关性。本研究结果表明,阿拉善型绒山羊皮肤组织中RHBDF2
基因的表达量在兴盛期最高,生长发育至退行期,RHBDF2
基因的表达量逐渐降低,在休止期和兴盛前期的表达量最低。敏盖绒山羊皮肤组织中RHBDF2
基因的表达量在兴盛期最高,且在退行期和兴盛前期的表达量大于休止期休止期。同时对RHBDF2
基因与绒山羊皮肤毛囊性状进行相关性分析发现,RHBDF2
基因与皮肤毛囊各性状均显著相关(P
<0.05)。说明高表达的RHBDF2
基因可以促进皮肤毛囊生长。Hosur等在小鼠皮肤损伤伤口愈合反应的研究中发现,RHBDF
2基因通过表皮生长因子受体(Epidermal growth factor receptor,EGFR)信号的转导参与上皮再生,可以通过增强表皮生长因子受体来加速皮肤伤口愈合,这与本研究结果相同。推测RHBDF2
基因是绒山羊皮肤毛囊周期性生长发育的重要影响因子;两个品系绒山羊皮肤组织中ALDH3A2
基因的表达量均是在休止期最高,且极显著大于其他3个时期(P
<0.01),退行期和兴盛前期极显著大于兴盛期(P
<0.01),兴盛前期显著大于退行期(P
<0.05)。可以发现,ALDH3A2
基因的表达量在皮肤毛囊休止期至兴盛期的过程中逐渐降低,由退行期至休止期的过程中逐渐增高。通过相关性分析发现,ALDH3A2
基因与皮肤毛囊各性状均显著相关(P
<0.05)。说明高表达的ALDH3A2
可能抑制皮肤毛囊的生长发育,是调控皮肤毛囊生长发育的重要影响因子。研究发现,Sjögren-Larsson综合征(SLS)是一种遗传性神经皮肤疾病,由脂肪醛脱氢酶(FALDH)基因诱导ALDH3A2
突变引起,导致脂肪醛脱氢酶缺乏,脂肪醛和脂肪醇积聚,会增加角质细胞的生长并且延缓皮肤屏障的恢复,ALDH3A2
在绒山羊羊绒发育过程中的作用有待进一步验证。3.4 chi-miR-107-3p和RHBDF2基因的表达差异相关性分析
通过对chi-miR-107-3p和RHBDF2
基因表达量进行分析发现,chi-miR-107-3p和RHBDF2
基因在绒山羊皮肤毛囊不同发育周期的表达量呈相反趋势。在次级毛囊重建初期,chi-miR-107-3p表达量较高,而RHBDF2
基因的表达量极低,随着次级毛囊重建逐步完成,chi-miR-107-3p表达量显著降低,RHBDF2
基因的表达量逐渐增高。直到进入退行期,chi-miR-107-3p的表达量逐渐增高,而RHBDF2
基因的表达量逐渐降低,休止期,chi-miR-107-3p的表达量达到最大值,此时RHBDF2
基因的表达量最小。同时,结合基因表达与皮肤毛囊性状的相关性分析,发现chi-miR-107-3p和RHBDF2
基因均与初级毛囊密度和真皮厚度显著相关(P
<0.05),且与次级毛囊密度、表皮厚度、初级和次级毛囊深度和毛球直径极显著相关(P
<0.01)。同时,利用miRbase预测了RHBDF2
基因与chi-miR-107-3p可能具有靶向关系,推测chi-miR-107-3p和RHBDF2
基因可能存在负调控关系,共同调控绒山羊皮肤毛囊的周期性生长发育,但这一关系仍需通过双荧光素酶报告进一步验证。3.5 展望
本研究通过观察发现不同品系的绒山羊皮肤毛囊组织显微结构相近。结合chi-miR-107-3p及其相关基因的表达情况,皮肤毛囊从兴盛前期到兴盛期、退行期到休止期,chi-miR-107-3p和RHBDF2
基因表达量变化总相反。说明chi-miR-107-3p可能通过负向调控RHBDF2
基因的表达来抑制绒山羊皮肤毛囊的生长发育,但这一关系需进一步证实;根据ALDH3A2
基因在绒山羊皮肤组织中的表达情况与毛囊形态密切相关发现,ALDH3A2
基因可能是绒山羊皮肤毛囊周期性生长发育的重要影响因子。接下来的研究需进一步确定chi-miR-107-3p与其相关基因的靶向关系,及其对绒山羊毛囊生长发育的调控作用,对深入了解绒山羊皮肤毛囊周期性生长发育机制具有重要的意义。猜你喜欢
汽车实用技术(2022年19期)2022-10-19
今日农业(2022年14期)2022-09-15
阅读(中年级)(2020年3期)2020-06-01
新农业(2020年5期)2020-06-01
今日财富(2020年7期)2020-04-07
新农村(2018年35期)2018-04-02
中国美术(2016年4期)2017-01-12
宠物世界·猫迷(2016年3期)2016-04-23
黑龙江史志(2014年19期)2014-11-10
现代语文(学术综合)(2014年1期)2014-04-09
