
甜玉米籽粒体积和粒重的全基因组关联分析
2022-07-25 06:38肖颖妮李高科于永涛李光玉高颖珊胡建广
中国农业大学学报 2022年7期
肖颖妮 李高科 李 坤 于永涛 李光玉 李 武 高颖珊 胡建广
(广东省农业科学院 作物研究所/广东省农作物遗传改良重点实验室,广州 510640)
甜玉米是普通玉米中1个或几个基因发生突变而形成的一种特用玉米,其营养丰富,适口性好,风味独特,深受广大消费者青睐。随着人们生活水平的提高,甜玉米由于容易种植、经济效益好、生长周期短而深受种植农户的青睐,种植面积逐年增加,2019年,我国种植面积已达53万hm。高产稳产一直都是作物育种重要的目标。粒重和籽粒大小作为产量的主要构成因子,与产量直接相关。甜玉米籽粒鲜重和干重显著正相关(r
=0.89,P
=2.2×10),提高甜玉米干粒重不仅可以提高制种产量,也意味着鲜穗产量的提高,增加籽粒体积一定程度上也可以提高产量。因此,深入剖析甜玉米籽粒体积和粒重的遗传基础对提高甜玉米产量具有极其重要的意义。大量的籽粒突变体为克隆调控玉米籽粒粒重和籽粒大小的基因提供了宝贵的遗传资源。通过突变体克隆影响籽粒粒重和籽粒大小的基因主要包含3类:第一类是胚乳中淀粉合成代谢相关基因(sh2
,sh1
,bt1
,bt2
和su1
等),这些基因突变主要导致胚乳淀粉含量下降,从而使得粒重降低;第二类是影响胚发育的基因(emp2
,emp12
,dek14
和dek15
等),这些基因突变导致胚败育,最终籽粒发育不良;第三类是基部胚乳转移层区相关基因(mn1
),这类基因突变主要导致籽粒变小,粒重也随之降低。但是,突变体导致籽粒发生剧烈变化,极端的表型很难直接用于籽粒的遗传改良。利用QTL定位以及全基因组关联分析方法是解析数量性状遗传基础的重要手段,截止目前,已经定位了上千个与玉米粒重和籽粒大小相关的QTL (www.maizegdb.org/qtl)或位点,并成功克隆了4个与粒重相关基因,分别为编码己糖转运蛋白的ZmSWEET4c
、编码细胞壁转移酶的ZmINCW1
、编码BAM相关蛋白激酶的ZmBAM1d
和编码三角状五肽重复(PPR)蛋白的qKW9
。随着玉米基因组参考序列的测序完成以及测序技术的发展,全基因组关联分析(genome-wide association study, GWAS)已成功解析玉米多个复杂数量性状的遗传基础,包含籽粒品质性状、植株抗逆性状和株型性状等。然而,对甜玉米籽粒体积和粒重遗传基础解析的研究却鲜少报道。本研究以209份具有广泛代表性的甜玉米自交系为关联群体,结合全基因组重测序获得980万个SNP基因型,对甜玉米籽粒体积和粒重进行全基因组关联分析,挖掘控制甜玉米籽粒体积和粒重的相关基因,以期为甜玉米高产育种提供参考。
1 材料与方法
1.1 试验材料及田间试验
供试甜玉米材料共包含209份甜玉米自交系,其中,国内来源158份,国外来源51份(表1),由广东省农业科学院作物研究所提供。所有材料在广州市天河区连续种植3年(2017、2018和2019年),采用完全随机区组设计,每个材料种1 行,每行11株,行距0.5 m,株距0.25 m,按照当地栽培方法进行田间管理。
表1 本研究所使用的甜玉米自交系材料
Table 1 Sweet corn inbreds used in this study
编号ID名称Name地理来源Original region编号ID名称Name地理来源Original regionC11022中国C41新美中国C21132中国C42国区中国C3C5-1中国C44超甜201中国C4日超1日本C48航1132中国C5C4中国C49航日超日本C6794中国C50改良794中国C7日超2日本C51群2-7中国C9夏威夷美国C52群2白-3中国C10鲜美选系中国C53群2白-4中国C11华珍1中国C5408秋群1耐寒后代-2中国C12华珍3中国C5508秋群1耐寒后代-3中国C13265OR1美国C5608秋群1耐寒后代-4中国C15世珍中国C5708秋群1耐寒后代-5中国C16中农419中国C58王朝开放美国C17Z5阿根廷C59超引黄中国C18N16中国C60奥甜8706美国C20MH70美国C61金煌1号泰国C21库普拉选系美国C62国区T12-1中国C22金珠蜜美国C63国区T12-2中国C23金珠蜜白美国C64GLAY10黄-2美国C24曾061487泰国C65曾引me153-1泰国C25南引3中国C66曾引me153-2泰国C26群1穗选1中国C67QX331开放选黄中国C27群1穗选2中国C68QX331开放选白中国C28群1穗选3中国C69热群-3中国C30农甜88白中国C70群2-8中国C31农甜88黄中国C71群2-9中国C32225美国C72群2-10中国C33群1穗选4中国C73群2-11中国C35热群穗选中国C7408秋群1耐寒后代-6中国C36群1穗选6中国C75MH70×诱选甜中国C37SSC016美国C76群2甜诱选甜白中国C38奥甜美国C77李美引M6美国C39田杂中国C78开放华珍-1中国C40穗甜202中国C79开放华珍-2中国
表1(续)
编号ID名称Name地理来源Original region编号ID名称Name地理来源Original regionC8108秋泰引亲本泰国C118群2-1中国C8208秋泰引开放泰国C119群2-2中国C8308秋泰引组合-2泰国C120群2-3中国C8408秋泰引组合-3泰国C12108秋耐寒后代中国C8508秋泰引组合-4泰国C12208秋耐寒后代白-1中国C86HSC0803美国C12312春高引白-1中国C87HSC0803白美国C12412春高引-2白中国C88金银08白美国C12512春高引-3白中国C89田蜜6号中国C12612春高引-4白中国C90黄金15-2美国C12712春高引-5白中国C91泰引超甜-2泰国C12812春高引-6-1中国C92泰引白泰国C12912春高引-6-2中国C93泰引6高泰国C131GLAY10黄-1美国C94HIBRIX51大粒泰国C132群1-3中国C96湖北引-5白-1中国C133群2-4中国C97湖北引-5白-2中国C134群2-5中国C98湖北引-10中国C135群2-6中国C9909引-21中国C13709秋国区T9中国C10010春国区T9中国C138群1甜诱白中国C10110春国区T9白中国C139MH70×诱中国C102华珍母本中国C140群2甜诱-1中国C10309国区T4中国C141群2甜诱白中国C104群1-1中国C142群1甜诱 中国C105群2白-1中国C143群2甜诱 混中国C106群2白-2中国C144群2甜诱 单穗中国C107德甜36白中国C145群2甜诱-2中国C108S103中国C146MH80开放美国C109大28-2中国C14708秋泰引组合-1泰国C11012春高引9-1中国C148黄金15-1美国C11112春高引9-2中国C149泰引超甜-4泰国C112丰蜜中国C15009引-2-1中国C113群1-2中国C15109引-2-2中国C114热群-1中国C15209引-11中国C115热群-2中国C15510秋柯引彩-4中国C116群1-1-1中国C15710秋柯引彩-7白中国C117群1-1-2中国C158库普拉×诱美国
表1(续)
编号ID名称Name地理来源Original region编号ID名称Name地理来源Original regionC15911春GT3-1中国C202华宝8号中国C16011春GT3-2中国C203泰丰母泰国C16111春GT8-1中国C204泰丰父泰国C16211春GT8-2中国C2525#-7中国C16311春GT11中国C253A453海-4-2中国C164群1-1-3中国C256A003海中国C16508秋耐寒后代白-2中国C25704冬Z15-1中国C166SB903美国C25804冬Z19中国C167921似亲本泰国C25904冬Z52-1中国C169群1-4中国C260HN-6中国C171华美甜似亲本白-1中国C261xsj2号中国C172华美甜似亲本白-2中国C264A126-1中国C173美国225美国C272ZHU2中国C175山西引超甜中国C274JFM中国C17612春高引-7中国C27507SHXJF中国C17712春高引-10中国C27610QXJFM中国C17812春高引-10白-1中国C277XJLN中国C17912春高引-10白-2中国C281737中国C18012春高引-13中国C2825#中国C18112春高引-17中国C2841167-广西TF中国C182奥费兰选系美国C285AB1000 白粒超甜中国C183SC1388泰国C28936-1 白粒超甜中国C18608秋群1耐寒后代-1中国C292A02中国C18708秋群1耐寒后代×诱选甜中国C294A04中国C189群2 叶披,多中国C307A17中国C193农宝白中国C308A18中国C194麦哥娜姆选系美国C311A21 白粒超甜中国C195华威2045泰引开放系中国C315A25中国C196新美208中国C317A27中国C197SC858泰国C318A28 白粒超甜中国C198Yes37泰国C320A30中国C199改良泰引组合6白泰国C323A33中国C200仲鲜甜3号中国C329A39 白粒超甜中国C201群1白中国
1.2 表型调查及统计分析
所有材料自交授粉,收获后自然晾干至含水量13%以下,每个材料选取5个均匀一致的果穗混合脱粒,取其中300粒装袋,再从中随机选取30粒干籽粒进行后续表型鉴定。将籽粒去麸后,使用分析天平称量30粒籽粒重量(w
),计算得单粒重(w
/30),g;采用排酒精法对籽粒体积进行测量,取30粒干籽粒倒入装有酒精的滴定管中,读取加入籽粒前后液面刻度变化的差值,即为30粒籽粒体积(V
),计算得单粒体积(V
/30), mL。利用R软件对3年表型进行统计分析和相关性分析,并计算广义遗传力和最佳线性无偏预测值(best linear unbiased prediction, BLUP)。广义遗传力计算公式为:
(1)
式(1)中:代表遗传变异,代表误差,e
代表环境个数。BLUP计算公式为:
y
=μ
+g
+e
+ε
(2)
式(2)中:y
代表第i
个家系的表型值,μ
代表所有环境下的均值,g
代表遗传效应,e
代表环境效应,ε
代表随机误差。1.3 基因型分析
取每个自交系3~5棵植株的幼嫩叶片,采用改良的CTAB方法提取DNA,利用illumina HiSeq 2500平台对209份材料进行深度约11.5×的全基因组重测序,用BWA软件将测序序列比对到玉米‘B73’参考基因组(AGPv4),而后用SAMTOOLS软件对SNP进行分析,最后用GATK软件对SNP进行提取,保留缺失率≤20%以及最小等位基因频率≥5%的SNP,最终获得980万个覆盖全基因组的高质量的SNP(数据未发表)用于后续分析。
1.4 全基因组关联分析及候选基因提名
将基因型和表型导入TASSEL5.0中,采用一般线性模型(General Linear Model, GLM)进行全基因组关联分析。由于非独立的SNP之间有较强的LD距离,为了进一步更准确的确定阈值,利用PLINK软件(window size 50, step size 50,R
≥0.2)计算得到767 964个独立标记数,因此,将关联分析的显著性阈值设为P
<1×10(1/767 964)。根据之前对甜玉米自交系LD的计算,甜玉米的衰减距离为500 kb,因此,当500 kb区间内存在多个显著SNP时,合并到1个最显著的SNP,查找其上下游500 kb内的基因,根据基因注释并结合基因表达谱数据,找寻与其他植物籽粒体积或粒重有关基因为候选基因,如无相关基因,提名离显著位点最近且检测到表达量有差异的基因为候选基因。针对每个表型,所有显著位点解释的表型变异用R软件中的lm
进行线性回归分析计算而得。1.5 候选基因表达量与表型的相关分析
所有材料自交授粉,采摘授粉后15 d(15 DAP)的果穗,分别选取每个自交系的3~4穗,每穗取1~2粒籽粒,混合提取籽粒RNA构建总RNA文库,利用Illumina Hiseq 2500平台对文库进行150 bp的双端转录组测序,每个样品获得了大约24.76 M高质量的Reads,利用R软件对每个样品中所有基因的FPKM进行计算,最终获得了27 133个基因在不同自交系中的表达谱,利用R软件对候选基因在不同自交系中的表达量和其对应的表型进行相关分析。
2 结果与分析
2.1 甜玉米籽粒体积和粒重表型分析
由图1可知,籽粒体积变异范围为0.06~0.15 mL,变异倍数为2.5倍,单粒重变异范围为0.08~0.17 g,变异倍数为2.1倍(表2),表明这2个性状在群体中都具有广泛的表型变异,并且这2个性状在群体中都呈正态分布(图1)。此外,籽粒体积和粒重呈显著正相关(r
=0.92)(图1)。利用3年的表型进行遗传力分析,籽粒体积和粒重的遗传力分别为0.70和0.79(表2),表明2个性状的表型变异主要受遗传因素控制,受环境因素影响较小。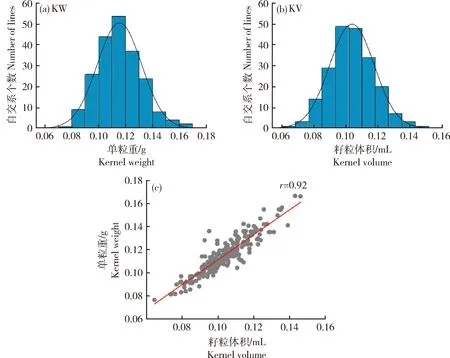
(a)籽粒粒重表型分布;(b)籽粒体积表型分布;(c)粒重和体积的相关性分析(a) Distribution of kernel weight; (b) Distribution of kernel volume; (c) The correlation coefficients between kernel weight and kernel volume图1 甜玉米群体籽粒体积和粒重的表型分析Fig.1 Phenotypic analysis of kernel volume and kernel weight in the sweet corn panel
表2 甜玉米籽粒体积和粒重的表型分析和遗传力分析
Table 2 Phenotypic variation and heritability of two traits in sweet corn kernels
性状Trait自交系个数No. of lines最小值Min最大值Max平均值±标准差Mean±SD广义遗传力H2籽粒体积/mLKV2090.060.150.10±0.010.70单粒重/gKW2090.080.170.12±0.020.79
2.2 籽粒体积和粒重全基因组关联分析
由表3可知,利用GLM模型,当阈值为P
<1×10时,共检测到15个显著SNP位点,其中,籽粒体积检测到9个位点,单个位点解释的表型变异范围为10.6%~13.6%,总共解释55.7%的籽粒体积表型变异;籽粒粒重检测到10个位点,单个位点解释的表型变异范围为9.3%~12.6%,总共解释52.1%的粒重表型变异。由表4可知,在这15个位点中,有4个位点和2个性状均显著关联,表现“一因多效” 性(图2),这与籽粒体积和粒重呈显著正相关的结果是一致的(图1)。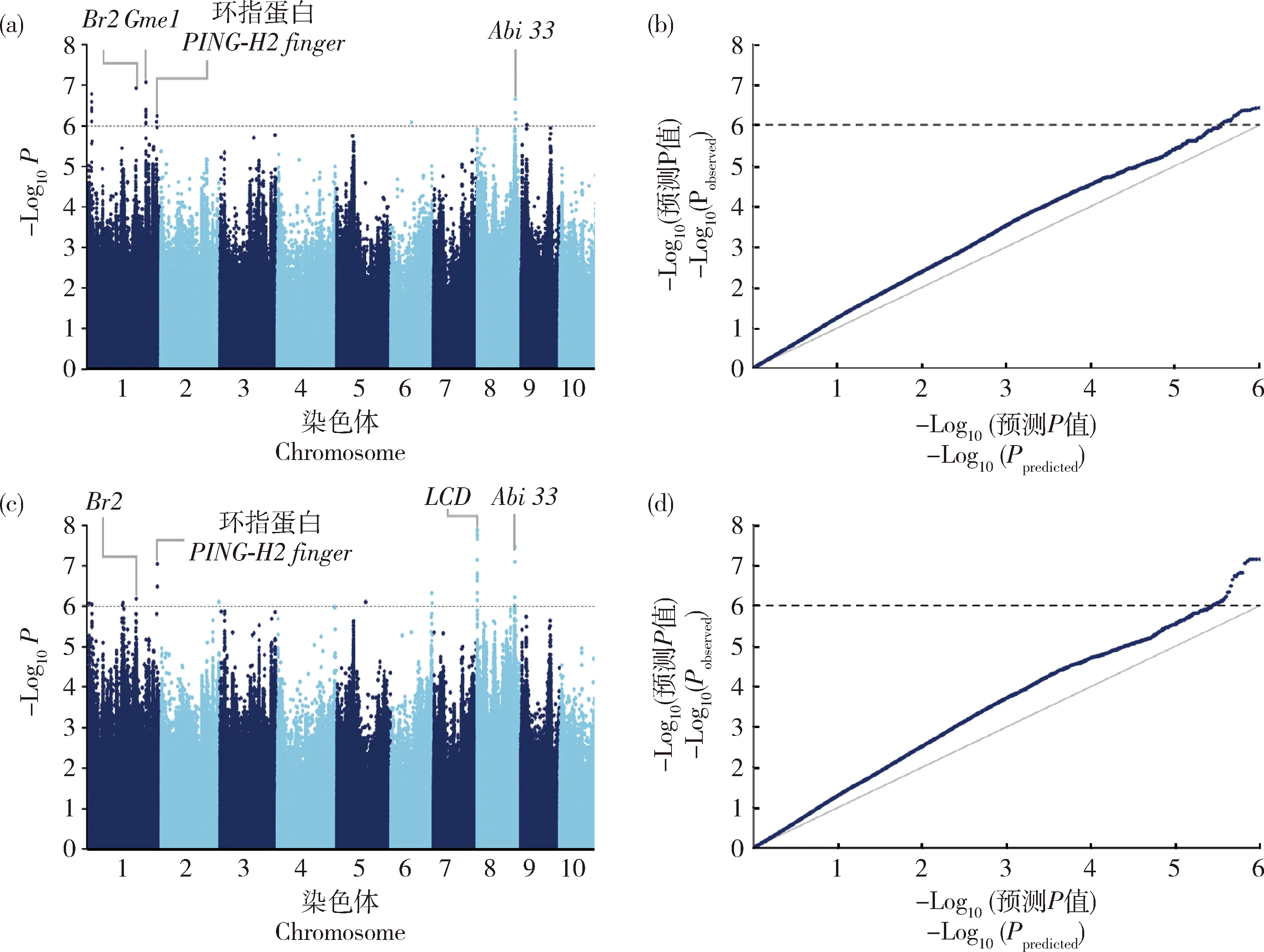
(a)籽粒体积的曼哈顿图;(b)籽粒体积的Q-Q图;(c)单粒重的曼哈顿图;(d)单粒重的Q-Q图(a) Manhattan plot of kernel volume; (b) Quantile-quantile plot of kernel volume; (c) Manhattan plot of single kernel weight; (d) Quantile-quantile plot of single kernel weight水平虚线代表阈值(1×10-6)。The dashed horizontal line depicts the Bonferroni significance threshold (1×10-6).图2 甜玉米籽粒体积和粒重全基因组关联分析Fig.2 Genome-wide association study of kernel volume and kernel weight in sweet corn
表3 甜玉米籽粒性状显著位点
Table 3 Summary of significant SNPs identified for kernel traits in sweet corn
性状Trait显著位点数目Significant SNPs单个位点解释表型变异/%Variation explained by each SNP所有位点解释表型变异/%Variation explained by all SNPs籽粒体积/mLKV910.6~13.655.7单粒重/gKW109.3~12.652.1
2.3 与普通玉米研究位点比较
由表4可知,3个与籽粒体积关联的位点均位于普通玉米籽粒大小相关性状的QTL区间内。其中,S1_204753078位点与籽粒体积和粒重均显著关联, S1_288353444与籽粒体积显著关联,这2个位点均落在控制粒宽的QTL区间内,S9_24601539与籽粒体积显著关联,该位点落在粒长的QTL区间内;此外,S1_291060760与粒重显著关联,该位点落在普通玉米千粒重的QTL区间内。综上,通过检测的位点和普通玉米籽粒有关性状位点进行比较分析,说明利用全基因组关联分析方法挖掘与甜玉米籽粒性状显著关联位点方法可靠。
2.4 候选基因预测和功能分析
基于玉米参考基因组数据,对显著关联位点上下游500 kb范围内的基因进行预测分析,结合甜玉米籽粒15 DAP的表达谱数据,最后挖掘到15个候选基因。大部分候选基因都是转录因子等调控基因,见表4,比如控制籽粒体积的Zm00001 d034277和Zm00001 d011730分别为NAC转录因子和乙醇脱氢酶转录因子,控制粒重的Zm00001 d030577是bZIP转录因子,此外,控制籽粒体积和粒重的Zm00001 d034369和Zm00001 d011639分别是环指蛋白和脱落酸不敏感转录因子。
1号染色体上的位点S1_204753078与籽粒体积、粒重均显著关联,该位点位于基因Br2
(Zm00001 d031871)第3个内含子上,Br2
基因编码生长素转运蛋白。通过对该显著位点进行单倍型分析发现,基因型CC与基因型TT的植株相比,株高有一定程度的增加,穗位高、籽粒体积和粒重均有显著增加,因此,Br2
和甜玉米籽粒体积、粒重均显著关联,并且,大部分自交系都包含有利等位基因CC(n=129),见图3。由图4可知,不同玉米材料授粉15 d 后Br2
表达量与籽粒体积和粒重均呈显著正相关(r
=0.37,P
=1.18×10和r
=0.36,P
=1.28×10),说明该基因是通过调控基因的表达量从而引起表型发生变化的,亦即随着该基因表达量的增加,籽粒体积和粒重均随之增加。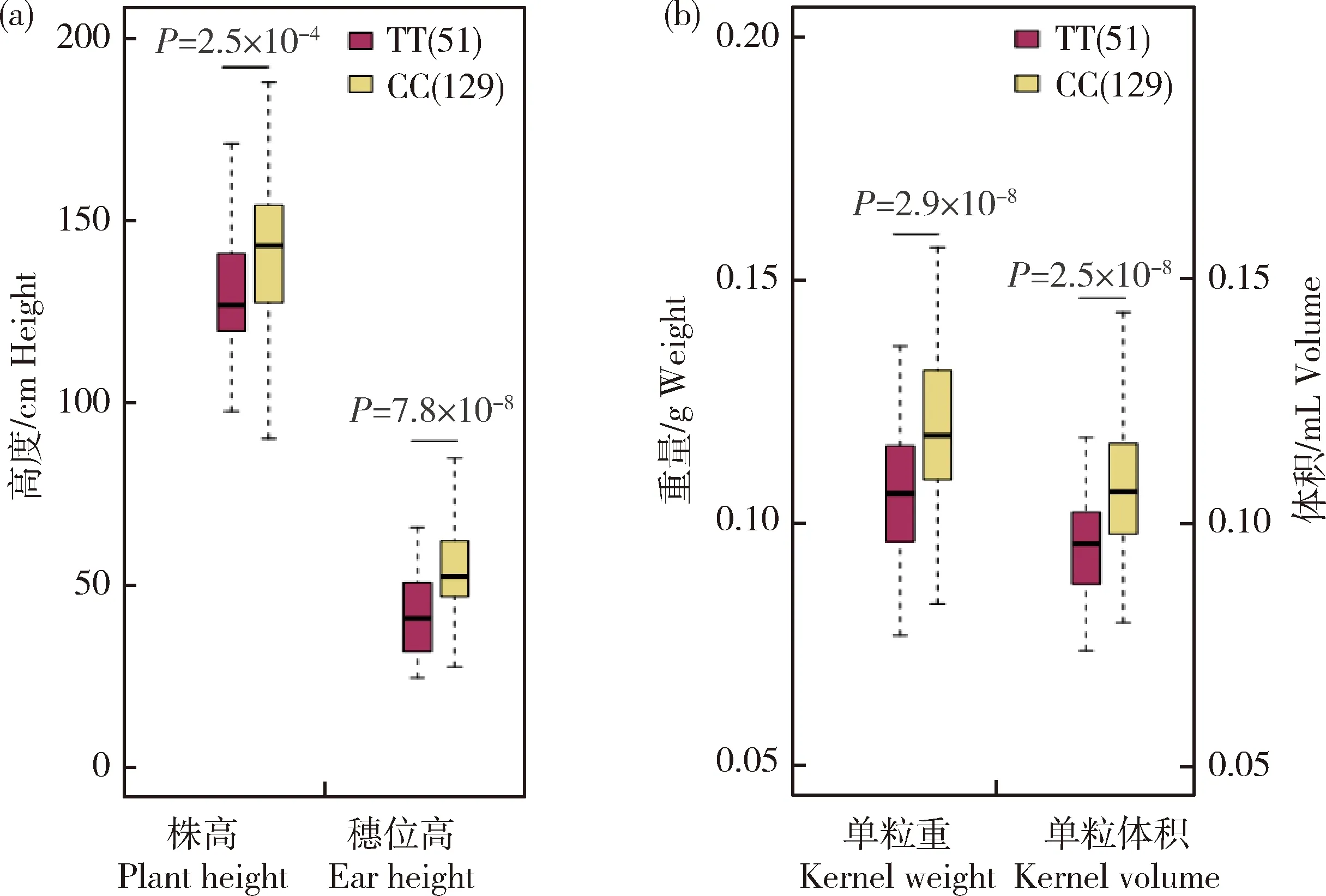
图3 Br2上显著位点与不同农艺性状的方差分析Fig.3 ANOVA based on the most significant associated SNP of Br2 with agronomic traits

图4 Br2基因表达量与籽粒体积(a)和单粒重(b)的相关性分析Fig.4 Correlation of expression level of Br2 with kernel volume (a) and kernel weight(b)
3 讨 论
3.1 甜玉米籽粒体积和粒重的全基因组关联分析
尽管甜玉米育种更多关注的是其风味和营养品质,然而提高产量也是甜玉米育种研究的重要目标之一,籽粒体积和粒重是与甜玉米产量相关的最重要的农艺性状。本研究对该关联群体进行采收期甜玉米籽粒含水量的测定,发现籽粒的含水量平均为75%,变异范围为61%~85%,籽粒干粒重和鲜重相关性极显著(r
=0.89,P
=2.2×10),因此,甜玉米籽粒干重的增加,意味着籽粒鲜重也增加,有助于提高甜玉米的产量。GWAS是解析复杂性状的遗传结构以及挖掘关键基因的重要手段,本研究连续3年对209份甜玉米自交系的干籽粒体积和粒重进行了测定,共检测到了15个显著位点,但单个位点的遗传效应均不高(9.3%~13.6%),表明这2个性状主要受微效多基因控制,符合产量是复杂数量性状的特点。通过与徐运林等报道的甜玉米百粒重的关联SNP进行比较,并未发现有共同检测到的位点,这可能因为所使用的材料不同,也说明了粒重性状的复杂性。
3.2 甜玉米籽粒体积和粒重的候选基因挖掘
本研究中检测到Br2
基因与籽粒体积、粒重均显著关联,该基因编码的是一个生长素极性运输蛋白PGP;生长素转运蛋白和信号因子通过调节植物生长素的极性运输对植物的生长发育起着重要作用,玉米de
18
突变体中生长素合成受到影响,从而抑制了细胞的分裂,最终使种子粒重降低。在普通玉米中,Br2
基因突变体主要通过减少植株纵向细胞分裂,导致植株变矮,穗位变低,并且不影响产量,此外,Li等发现在普通玉米自然变异群体中,Br2
与籽粒长度显著关联。在甜玉米中,检测到与籽粒体积、粒重最显著关联的位点位于基因Br2
内含子上,这与Li等的研究结果类似,该位点可能并不是影响籽粒体积和粒重的功能位点,而是与功能位点存在连锁不平衡(LD)而被检测到的。Br2
的表达量和籽粒体积以及粒重均显著相关(r
=0.37和r
=0.36),这与该位点解释的表型变异(13.6%和10.6%)的结果相符,说明该位点并不是主效位点,对籽粒体积和粒重的变异只起到微效作用,推测功能位点可能存在于影响基因表达水平的调控区(5’UTR或3’UTR),通过调控基因的表达水平,引起粒长的变化,最终导致籽粒体积和粒重都增加,因此,还需要通过候选基因重测序以及基因验证等手段进一步来验证。此外,在甜玉米中,该基因在调控籽粒产量(单粒重和籽粒体积)的同时,也调控穗位高,而对株高的影响相对较小(图3),因此,通过合理选育,有望选育出籽粒体积和粒重均增加,而株高和穗位高相对合理的优良品种。本研究在9号染色体上发现1个与籽粒体积关联的候选基因Zm00001 d045499,该基因位于Liu等报道的控制籽粒粒长的QTL区间内,编码1个E3泛素蛋白连接酶,水稻中克隆的控制粒宽和粒重的GW2
基因也是1个E3泛素蛋白连接酶,GW2蛋白缺失后,颖壳内细胞数量增加,从而导致籽粒变宽、变重。此外,在拟南芥中,DA2
编码1个E3泛素蛋白连接酶,功能缺失突变体的种子体积由于细胞分化未被抑制而显著增加。Zm00001 d045499在甜玉米中调控籽粒体积变化的作用机制还有待进一步验证。4 结 论
本研究结果表明,本群体(209份甜玉米自交系)包含广泛的遗传变异,结合覆盖全基因组高密度的980万个SNP标记,利用GLM模型通过全基因组关联分析方法来挖掘控制甜玉米产量性状的关联位点,共检测到15个显著关联位点,大部分候选基因是转录因子;其中,Br2
与籽粒体积、粒重均显著关联,该基因主要通过提高表达量来增加甜玉米籽粒体积和粒重。猜你喜欢
中国现代医生(2022年21期)2022-08-22
瞭望东方周刊(2021年6期)2021-03-30
瞭望东方周刊(2021年6期)2021-03-30
三农资讯半月报(2020年2期)2020-03-09
安徽农业科学(2019年14期)2019-08-27
现代农业研究(2019年3期)2019-06-05
农民致富之友(2017年12期)2018-01-31
农民致富之友(2017年23期)2018-01-02
吉林农业(2017年7期)2017-07-12
农业与技术(2016年8期)2016-10-21
