
基于点突变库研究丝切蛋白在烟曲霉抗逆性和致病力中的作用
2022-07-19 12:31房凌旭卢中一
中国生物制品学杂志 2022年7期
房凌旭,卢中一
1. 深圳市前海蛇口自贸区医院口腔科,广东 深圳 518067;
2. 深圳大学高等研究院深圳市海洋微生物组工程重点实验室,广东 深圳 518060
烟曲霉(Aspergillus fumigatus)是自然界中广泛存在的曲霉属真菌,其大量产生的分生孢子极易播散,可被人体吸入肺部,在一定条件下对免疫力低下人群产生致病性。烟曲霉可引起过敏性支气管肺曲霉病、侵袭性曲霉病和慢性肺曲霉病等[1-3]。特别是烟曲霉具有较高的抗逆特性和致病力,一方面体现在其细胞壁的葡聚糖和几丁质等成分可增强其对宿主免疫系统的逃逸能力;另一方面体现在其具有较强的抗氧化能力,可抵抗巨噬细胞产生的活性氧簇等[4-5]。因此,研究烟曲霉抗逆性和致病力的关键蛋白及相关分子机制是治疗烟曲霉感染的重要前提。目前,三唑类药物是临床上治疗烟曲霉感染的一线药物,但临床和环境中唑类药物及其衍生物的广泛施用导致烟曲霉逐渐产生耐药性问题[6-7]。我国的一项研究显示,从临床样品分离得到的159 株烟曲霉中,对伊曲康唑耐药的烟曲霉阳性率为4.4%[8];而SEWELL等[9]发现,从英国南部收集到的178 份土壤样品中检测到的唑类耐药烟曲霉阳性率达6.7%。
丝切蛋白(Cofilin)是一种保守存在于真核生物中的肌动蛋白结合蛋白,其通过切割肌动蛋白丝(Factin)调节肌动蛋白骨架重排。现有研究表明,Cofilin是参与真菌抗逆机制和耐药机制的关键蛋白,为具有研究价值的抗真菌药物靶点[10-11]。酿酒酵母Cofilin中部分蛋白氨基酸位点突变可影响线粒体功能,进而产生逆行信号造成部分转运蛋白基因转录水平升高,最终引起突变株对药物的耐受升高[12]。此外,烟曲霉Cofilin基因高转录水平可引起菌株内氧化应激相关基因和细胞壁多聚糖合成基因转录水平升高,提示Cofilin 在烟曲霉抗逆性和致病力方面可能具有重要作用[13]。本研究探讨了Cofilin 点突变库在烟曲霉抗逆性和致病力等方面的作用,以期为Cofilin作为治疗烟曲霉感染的潜在药物靶点提供理论依据。
1 材料与方法
1.1菌株 烟曲霉野生株CEA17ku80Δ(WT)由法国巴斯德研究所Jean Paul Latge'教授惠赠;烟曲霉Cofilin 点突变菌株S5A、D19A-R21A、E48A、K36A、K36E 和D54A 由中国人民解放军疾病预防控制所韩黎研究员惠赠。
1.2实验动物 大蜡螟幼虫购自玉米虫农场公司。选取体表光滑,颜色浅亮,体重0.3 ~ 0.35 g 的大蜡螟幼虫进行实验。
1.3主要试剂 荧光白、刚果红和FITC 偶联的伴刀豆蛋白购自西格玛奥德里奇贸易有限公司;过氧化氢(H2O2)和山梨醇购自北京国药集团化学试剂有限公司;琼脂购自青岛海博生物科技有限公司;PCR 酶体系购自宝生物工程大连有限公司。
1.4培养基的配制 基础营养培养基(MM)为深圳大学高等研究院深圳市海洋微生物组工程重点实验室配制,成分包括:1 mL Trace element solution[FeSO4·7 H2O 5 g/L、ZnSO4·7 H2O 22 g/L、MnCl2·4 H2O 5 g/L、Na4EDTA 50 g/L、(NH4)6Mo7O24·4 H2O 1.1 g/L、H3BO311 g/L、CoCl2·5 H2O 1.6 g/L 和CaCl2·2 H2O 14.7 g/L]、50 mL 20 × Salt Solution 溶液(KH2PO430.4 g / L、MgSO4·7H2O10.4g/L、KCl10.4g/L 和NaNO3120g/L)、10 g 葡萄糖和15 g 琼脂;调节MM 的酸碱度,配制成pH 10 的MM 培养基;在MM 中加入100 mg / L 荧光白,配制成荧光白培养基;在MM 中加入200 mg/L 刚果红,配制成刚果红培养基;在MM 中加入100 mg/L的H2O2,配制成H2O2培养基;在MM 中加入2.4 mol/L山梨醇,配制成山梨醇培养基。
1.5烟曲霉Cofilin 点突变库的构建 引物序列见表1,由华大基因科技有限公司合成。通过引物F1和R1(引入突变位点),以烟曲霉菌株CEA17ku80Δ基因组为模板扩增Cofilin 编码基因片段及上游同源臂(长度1 000 ~ 1 500 bp),同时利用引物F2 和R2 扩增上述基因组中Cofilin 编码基因另一部分片段;此外,分别通过引物F3 和R3、F4 和R4 以含有筛选标记的质粒和烟曲霉基因组为模板,扩增筛选标记hph 片段及Cofilin 编码基因下游同源臂(长度1 000 ~ 1 500 bp);最终利用引物F1 和R4 将上述片段按浓度比1 ∶3 ∶3 ∶1 进行片段融合,并采用原生质体转化方式导入CEA17ku80Δ 中原位替换Cofilin编码基因,经筛选并使用引物seqS 和seqA 测序验证后获得烟曲霉Cofilin 点突变株。

表1 烟曲霉Cofilin 点突变库引物Tab.1 Primers for Cofilin point mutation library of A.fumigatus
1.6烟曲霉Cofilin 点突变库的抗逆性检测 通过应激试验分析Cofilin 点突变库在不同刺激下对烟曲霉抗氧化、抗高渗透压和抗细胞壁干扰等方面的影响。将WT 和Cofilin 点突变库烟曲霉的孢子用灭菌磷酸盐缓冲液稀释至5 × 107个/ mL,分别取2 μL 接种至不同成分培养基,在37℃下培养60 ~72 h 后观察菌落生长情况。
1.7烟曲霉Cofilin 点突变库细胞壁甘露糖合成及分布的检测 通过FITC 偶联的伴刀豆蛋白荧光标记Cofilin 点突变库烟曲霉中的甘露糖,分析Cofilin点突变对烟曲霉细胞壁中甘露糖合成及分布的影响。用含0.01% Tween 20 的灭菌磷酸盐缓冲液稀释烟曲霉孢子至5 × 107个/ mL,取2 μL 接种至含2 mL MM 液体培养基的6 孔板(底部放置10 mm×10 mm 盖玻片),在37 ℃下培养14 ~16 h;加入200 μL FITC 偶联的伴刀豆蛋白,在30 ℃下孵育1 h;用去离子水清洗2 次,将盖玻片取出后倒扣至含抗荧光淬灭剂的载玻片上,于荧光显微镜下观察并拍照。
1.8烟曲霉Cofilin 点突变库的致病力分析 采用大蜡螟幼虫感染试验分析Cofilin 点突变对烟曲霉致病力的影响。用含0.01% Tween 20 的无菌磷酸盐缓冲液将5 × 107个/ mL 烟曲霉孢子重悬。用微量注射器将10 μL 孢子悬液注入大蜡螟幼虫左排最下端腹足。设2 个对照组,一组大蜡螟幼虫注射10 μL 含0.01% Tween 20 的无菌磷酸盐缓冲液,另一组不做任何处理。每组16 只。注射后置于37 ℃遮光培养,每12 h 记录大蜡螟幼虫存活情况。
1.9统计学分析 应用GraphPad Prism 5 软件进行大蜡螟幼虫感染实验生存曲线及显著性分析,P值及显著性分析采用Log-rank(Mantel-Cox)检验。以P<0.05 为差异有统计学意义。
2 结 果
2.1烟曲霉Cofilin 点突变库的位点选取及构建本实验构建了烟曲霉Cofilin 点突变位点K26A、E56A、D11A-E12A、K41A-K42A 和R59A-R61A。选取的烟曲霉Cofilin 点突变库包括:单位点突变S5A(第5 位点丝氨酸突变为丙氨酸)、K26A(第26 位点赖氨酸突变为丙氨酸)、K36A(第36 位点赖氨酸突变为丙氨酸)、K36E(第36 位点赖氨酸突变为谷氨酸)、E48A(第48 位点谷氨酸突变为丙氨酸)、D54A(第54 位点天冬氨酸突变为丙氨酸)、E56A(第56 位点谷氨酸突变为丙氨酸)以及双突变D11A-E12A(第11 位点天冬氨酸和第12 位谷氨酸均突变为丙氨酸)、D19A-R21A(第19 位点天冬氨酸和第21 位点精氨酸均突变为丙氨酸)、K41A-K42A(第41 位点赖氨酸和第42 位点赖氨酸均突变为丙氨酸)和R59A-R61A(第59 位点精氨酸和第61 位点精氨酸均突变为丙氨酸),见图1。筛选及测序结果表明烟曲霉Cofilin 点突变株构建正确。
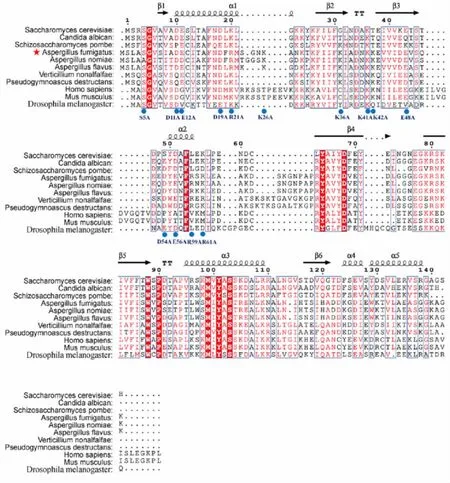
图1 烟曲霉Cofilin 点突变库中选取的突变位点Fig.1 Mutation sites selected from Cofilin point mutation library of A.fumigatus
2.2烟曲霉Cofilin 点突变库的抗逆性 相比于WT,Cofilin 点突变库中携带D11A-E12A、K26A、E56A 和R59A-R61A 的烟曲霉在MM 培养基上菌丝生长能力减弱,提示Cofilin 及其部分氨基酸位点对烟曲霉菌丝生长具有重要作用。此外,Cofilin 点突变库中烟曲霉对细胞壁干扰剂荧光白(100 mg / L)和刚果红(200 mg / L)的敏感性与WT 无显著差异,提示这些Cofilin 点突变可能不影响烟曲霉细胞壁完整性及细胞壁相关应激信号通路。在pH 10 的MM 培养基和含有山梨醇的高渗培养基中,Cofilin 点突变库中烟曲霉与WT 生长状态无显著差异,提示这些Cofilin 点突变未影响烟曲霉对酸碱度和高渗透压的应激能力。但携带D19A-R21A 和K36A 的烟曲霉菌株在H2O2培养基中菌丝的生长能力强于WT,提示这些突变导致烟曲霉对外界氧化压力的耐受水平升高;而携带K36E 的烟曲霉菌株在H2O2培养基中的生长受到明显抑制,提示该突变导致烟曲霉对氧化压力的耐受能力减弱。上述结果提示,Cofilin 及其部分氨基酸位点在烟曲霉氧化应激信号通路中具有重要作用。见图2。

图2 烟曲霉Cofilin 点突变库抗逆性试验Fig.2 Stress resistance test of Cofilin point mutants of A.fumigatus
2.3烟曲霉Cofilin 点突变库细胞壁中甘露糖的合成及分布 荧光显微镜下观察显示,Cofilin 点突变菌株与WT 的甘露糖含量无明显区别,但携带K36A和E56A 的烟曲霉菌株出现菌丝顶端甘露糖聚集现象,提示这些Cofilin 位点突变可能影响了细胞壁中甘露糖的分布,见图3。

图3 荧光标记Cofilin 点突变库细胞壁中的甘露糖观察Fig.3 Microscopy of fluorescent-labeled mannose in cell walls of Cofilin point mutants
2.4烟曲霉Cofilin 点突变库的致病力 S5A、D11AE12A、K36E、K41A-K42A、E48A、D54A 对大蜡螟幼虫的致病力与WT 相比,差异无统计学意义(χ2分别为0.182、0.495 9、0.124 4、0.567 5、0.068 89、0.236 6,P分别为0.613 4、0.561 6、0.724 3、0.551 3、0.793、0.626 7),提示这些Cofilin 点突变未显著性影响烟曲霉对大蜡螟幼虫的致病力。此外,与WT 相比,D19A-R21A、K26A 和K36A 使烟曲霉对大蜡螟幼虫的致病力显著增强(χ2分别为5.364、6.6、13.39,P分别为0.020 6、0.010 2、0.000 2),而E56A 和R59AR61A 使烟曲霉对大蜡螟幼虫的致病力显著减弱(χ2分别为4.443、4.18,P分别为0.035 1、0.040 9)。上述结果显示,Cofilin 及其部分氨基酸位点对烟曲霉的致病力具有重要作用。见图4。

图4 大蜡螟幼虫感染试验分析Cofilin 点突变对烟曲霉致病力的影响Fig.4 Analysis of effect of Cofilin point mutation on virulence of A.fumigatus by infection test in Galleria mellonella larval
3 讨 论
蛋白间相互作用是蛋白行使生物学功能的重要方式,而蛋白表面的电荷分布是决定蛋白间相互作用的关键因素[14]。本研究利用丙氨酸扫描构建的点突变库揭示了Cofilin 在烟曲霉抗逆性和致病力等方面的作用。丙氨酸扫描是将蛋白表面带电荷的氨基酸突变为丙氨酸,以评估蛋白功能及关键活性位点等。由于丙氨酸体积小且不带电荷,通常认为丙氨酸扫描仅改变蛋白表面的电荷分布而不影响蛋白结构[15-17]。本研究选择了烟曲霉Cofilin 中N-端的带电荷氨基酸,由于Cofilin 的N-端存在保守的磷酸化位点(已证明在烟曲霉Cofilin 的第五位丝氨酸[13]),而该位点附近的蛋白表面可能与Cofilin 上游调控蛋白的磷酸激酶和去磷酸酶存在相互作用,因此该处带电荷表面具有重要的生物学活性[18]。本研究中选取位点主要基于以下考虑:①Cofilin 是烟曲霉必需基因,过多的位点突变更易造成蛋白失活使菌株死亡,不利于点突变库的构建;②进行常规的多位点同时突变后,往往需要在突变框内进行下一轮单位点或双位点突变以确定关键功能位点,并不能根本性降低工作量;③本研究仅选取感兴趣的Cofilin N-端进行突变,整体点突变工作量不大;④位点的选取主要参照了前人对酿酒酵母Cofilin位点的研究工作,以便进行研究工作的横向比较[12];⑤选取的氨基酸位点主要集中于α 螺旋和β 折叠等蛋白二级结构区域,以便分析刚性区域在Cofilin生物学功能中的重要作用。其中,单位点突变包括S5A、K26A、K36A、K36E、E48A、D54A 和E56A,其中S5A 用来模拟Cofilin 去磷酸化过程,K26 用来分析曲霉属保守的Loop 区域中带电荷氨基酸位点的生物学功能,K36、E48、D54 和E56 的选取参考酿酒酵母Cofilin 的研究[12]。此外,本研究引入了K36E 作为K36A 的对照,以便分析不同突变模式在研究蛋白关键带电荷氨基酸功能中的作用。对于双突变位点,本研究选取了D11A-E12A、D19A-R21A、K41AK42A 和R59A-R61A,这些位点的选取参照了前人研究中酿酒酵母的Cofilin 点突变模式,可能参与调节线粒体功能[12]。
本研究结果显示,Cofilin 点突变D11A-E12A、K26A、E56A 和R59A-R61A 可减弱烟曲霉菌丝的生长能力,该现象的产生可能与点突变Cofilin 调控细胞骨架actin 功能的异常有关。现有研究证实,Cofilin表面的带电荷氨基酸对actin 有直接的调节作用。如酿酒酵母Cofilin 中氨基酸位点D10 和E11(分别对应烟曲霉Cofilin 中的D11 和E12)参与对细胞骨架actin 的调节功能,因此这些位点的突变可导致细胞骨架actin 形态异常[12]。其他研究显示,actin 与细胞的极性生长具有重要功能联系[11]。上述研究提示,Cofilin 及其部分带电荷氨基酸可通过调节actin的生物学功能影响烟曲霉菌丝的极性生长能力。
JIA 等[13]研究发现,Cofilin基因转录水平升高可导致烟曲霉氧化应激能力增强,提示Cofilin 可能参与烟曲霉的氧化应激信号通路。本研究进一步证实,Cofilin 中的带电荷氨基酸位点可能是该蛋白在烟曲霉氧化应激中行使功能的分子基础。值得注意的是,本研究发现,K36E 与K36A 对H2O2的耐受能力不同,提示该位点及附近的带电荷氨基酸(或化学侧链)可能调控了Cofilin 在烟曲霉氧化应激信号通路中的功能。同时也提示,除常见的丙氨酸和亮氨酸等点突变模式外,对蛋白中带电荷氨基酸进行反向电荷突变有助于对关键氨基酸功能及其分子机制的研究。
有研究发现,Cofilin基因转录水平升高可导致烟曲霉细胞壁中β-1,3-葡聚糖和几丁质等合成的增加,提示Cofilin 在烟曲霉细胞壁多糖的合成及细胞壁完整性信号通路中具有重要作用[13]。虽然本研究中的Cofilin 点突变对细胞壁中甘露糖的合成量无明显影响,但K36A 和E56A 可引起甘露糖在烟曲霉菌丝顶端的聚集,这种现象可能与细胞壁完整性信号通路或菌丝的极性生长有关。
本研究发现,Cofilin 点突变可影响烟曲霉对大蜡螟幼虫的致病能力,其中D19A-R21A、K26A 和K36A 导致烟曲霉对大蜡螟幼虫的致病力显著增强,产生这种现象的原因是这些点突变可能提高了烟曲霉中线粒体的功能活性。其依据为酿酒酵母Cofilin 中的双突变D18A-R20A(对应于烟曲霉Cofilin 的D19A-R21A)可导致线粒体功能异常活跃,而有研究证实,在格特隐球菌中,线粒体活性升高增强真菌的致病力[19]。此外,本研究同时发现,E56A 和R59A-R61A 引起烟曲霉对大蜡螟幼虫的致病力减弱,这一现象可能与这些菌株的菌丝生长能力减弱有关。
综上所述,本研究通过点突变库揭示了Cofilin及其氨基酸位点在烟曲霉抗逆性和致病力等方面具有关键作用,其相关的分子机制应是进一步研究的重点。随着研究的不断深入,Cofilin 作为治疗烟曲霉及其他致病真菌感染的潜在药物靶点将具有广阔的发展前景。
猜你喜欢
中国农业科学(2022年16期)2022-09-19
中国农业科学(2022年15期)2022-08-09
江苏农业科学(2021年8期)2021-05-26
电脑报(2020年40期)2020-11-06
三农资讯半月报(2020年18期)2020-10-14
农民致富之友(2019年31期)2019-11-23
电脑知识与技术(2018年19期)2018-11-01
现代交际(2016年20期)2017-02-15
饮食科学(2016年9期)2016-11-18
