
苹果砧木对低氮胁迫的响应及适应性评价
2022-07-18 08:59安秀红陈艳辉程存刚赵德英康国栋周江涛张艳珍
植物营养与肥料学报 2022年6期
解 斌,安秀红,陈艳辉*,程存刚*,赵德英,康国栋,周江涛,李 壮,张艳珍,杨 安,李 鑫
(1 中国农业科学院果树研究所/农业农村部园艺作物种质资源利用重点实验室/辽宁省落叶果树矿质营养与肥料高效利用重点实验室,辽宁兴城 125100;2 河北省国家北方山区农业工程技术研究中心/河北农业大学山区研究所,河北保定 071001)
氮是植物生长发育的必需营养元素之一,在氨基酸、蛋白质、核酸和激素等植物大分子物质的代谢途径中起关键作用,是植物生长发育的重要调控因子[1–3],且在植物适应逆境过程中发挥积极作用。苹果是重要的温带落叶果树之一,在其生长发育过程中对氮素有较大需求,土壤氮素亏缺会造成植物叶片黄化,叶绿素含量降低,光合能力减弱,阻碍植物的生长发育;此外,植物氮吸收量减少,会降解体内蛋白质以重新利用氮素,往往会导致叶片加速衰老,生物量积累降低[4]。生产中多因氮肥施用不当如施肥量不合理、施肥时期不科学等,造成氮肥利用率低、资源浪费及环境污染等问题[5]。因此,利用植物自身遗传特性,挖掘其自身养分利用潜力,在适度减氮条件下筛选耐低氮和氮高效吸收利用的砧木对苹果产业可持续发展具有重要意义。研究表明,与铵态氮(NH4+)、尿素等形态相比,硝态氮(NO3–)在促进果树营养生长中的作用更为显著[6]。NO3–不易被土壤胶体吸附,移动性较高,可被植株通过根的表皮和皮层细胞从土壤中迅速吸收利用[7]。因此,根系的形态可塑性反应影响植株对氮的吸收、积累与分配[8–9],而土壤氮素含量对根系构型也具有显著影响。在对玉米[10]、油菜[11]、糜子[12]等的研究中发现,低氮处理使得植株总根长、根表面积及根体积等均显著高于正常供氮处理,根冠比显著升高,且不同品种或基因型对低氮的响应存在差异[13–14]。此外,依靠硝酸还原酶(NR)等的作用实现NO3–在植物体内的高效转运和利用,以供植物生长发育所需,因此,NR活性能反映植株的营养状况以及NO3–同化水平[15]。砧木作为果树的重要组成部分,为其提供根系,直接影响果树对养分的吸收及对逆境的适应能力。有研究报道,柑橘不同砧木间以及嫁接接穗后树体养分吸收及抗逆性均存在较大差异[16–17]。康晓育[18]比较了山定子、八棱海棠、平邑甜茶等5种苹果乔化砧后发现,低氮条件下山定子的根冠比增幅较大,而平邑甜茶和八棱海棠的氮利用效率较高。近年来利用从国外引进的T337、B9、Pajam2等矮化自根砧栽培模式在各苹果主产区逐步推广,该模式相较于过去国内常用的乔化砧、矮化中间砧栽培模式,具有矮化效果显著、苗木整齐度高、节省劳动力、早果丰产性好等显著优势,然而国外引进砧木在国内的适应能力有待考究[19–20],同时国内自育的部分矮化砧木对低氮胁迫的响应尚不明确。植物对低氮胁迫的响应往往从植物生长发育、生理生化过程等多方面进行。适宜的多指标综合评价方法可全面反映作物不同品种耐低氮胁迫的能力。目前,这类报道主要集中在小麦、玉米、水稻等大田作物上,有关苹果砧木特别是矮化砧对低氮胁迫的适应性评价则少有研究[21–23]。本研究以7种苹果砧木为材料,通过盆栽沙培试验的方法,研究在正常供氮和低氮胁迫条件下树体生长、根系形态及其对氮素的累积及代谢的特征,利用隶属函数综合评价方法比较砧木间适应能力的差异,以期为耐低氮苹果砧木的选育及苹果高效吸收利用氮素生理机制的研究提供理论支撑。
1 材料和方法
1.1 试验材料
供试一年生苹果砧木共7种,其中,矮化砧包括T337、Nic29、Pajam2、B9、71-3-150,半矮化砧为青砧2号(Qingzhen 2)和乔化砧为山定子(Malus baccataL. Borkh.)。2019 年秋季定植于上口径 36 cm、下口径32 cm、高29 cm的黑色加仑盆中,栽培基质为洗净的河沙(25 kg),放置在辽宁省兴城市中国农业科学院果树研究所温泉试验基地苹果栽培示范园避雨棚内,定期浇灌改良1/2 Hoagland营养液以确保植株正常生长。
1.2 试验设计
试验于2020年秋梢生长期(7月1日)开始,每种砧木选择40株长势基本一致的健康植株进行试验处理,每个处理20株,单株重复。处理采用浇灌改良1/2 Hoagland营养液的方法进行,正常供氮处理(NN,5 mmol/L NO3–)的营养成分为 Ca (NO3)2·4H2O 2 mmol/L、KNO31 mmol/L;低氮胁迫处理 (LN,0.5 mmol/L NO3–)的营养成分为 KNO30.5 mmol/L。其他中量元素 (MgSO4·7H2O 2 mmol/L),微量元素(EDTA-Fe 0.05 mmol/L、MnSO4·H2O 0.5 mmol/L、CuSO4·5H2O 0.0005 mmol/L、ZnSO4·7H2O 0.15 mmol/L、H3BO30.5 mmol/L、Na2MoO4·4H2O 0.005 mmol/L),调节pH为5.9~6.5。低氮处理下,其他成分不变,缺少的Ca2+用CaCl2补足,缺少的K+用KCl补足。每隔 15 天浇透水一次 (1.5 L/盆),持续处理60 天。常规管理,保证其他试验条件一致。
1.3 样品采集及测定
1.3.1 植株生长势和生物量积累 在试验处理前期(ti),在供试植株中部选择3个长势中庸、无分枝的当年生新稍作为观测枝,标记后使用卷尺(精确度0.1 cm)测定长度(Hi),在处理60 天(tf)后再次测量长度(Hf),分别计算新稍生长速率(Ah)=(Hf–Hi) /(tf–ti),式中,Hf代表处理结束时新稍长度,Hi代表处理前新稍长度,(tf–ti)表示处理时间间隔[24]。之后,在不同供氮条件下各砧木分别选择5株具有代表性的植株进行株解,用去离子水反复冲洗干净后用吸水纸吸干表面水分,使用千分之一天平分别测定植株叶片、茎干及根系鲜重。
1.3.2 根系形态分析 待根系鲜重测定后,用EPSON扫描仪及WinRHIZO根系图像分析软件(Regent Instruments Inc.,加拿大)获取根系形态的相关指标,如根系总表面积、总根长、根系总体积、根尖数、根系平均直径和根系分枝数等。
1.3.3 氮含量测定 植株形态指标测定后将植株于105℃杀青30 min,80℃烘干至恒重,用万分之一天平测定植株各器官干重用于计算根冠比,之后用不锈钢磨样机将样品粉碎,过0.18 mm筛后保存备用。取烘干过筛后的样品0.3 g经H2O2‒H2SO4法消煮后,使用K9860全自动凯氏定氮仪(海能,中国)测定植株氮含量;以各器官氮含量与干重的乘积计算各器官氮累积量;以单株总干物质量与氮总累积量的比值衡量氮利用效率[25]。
1.3.4 耐低氮能力评价

不同砧木各指标的隶属函数值计算:
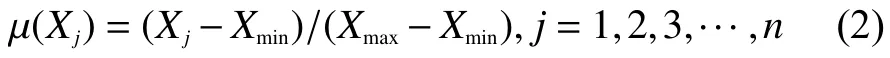
式中,μ(Xj)表示第j个指标的隶属函数值,Xj表示第j个指标的耐低氮胁迫指数;Xmax表示第j个指标的耐低氮胁迫指数的最大值,Xmin表示第j个指标的耐低氮胁迫指数的最小值。
利用熵值法计算各指标权重系数:
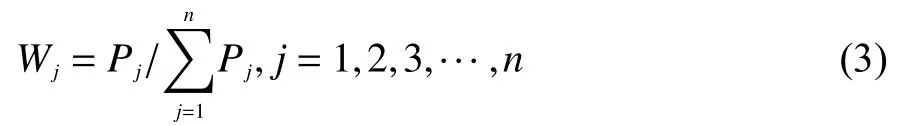
式中,Wj表示第j个指标在指标中的重要程度,即权重;Pj为各品种第j个综合指标的方差贡献率。
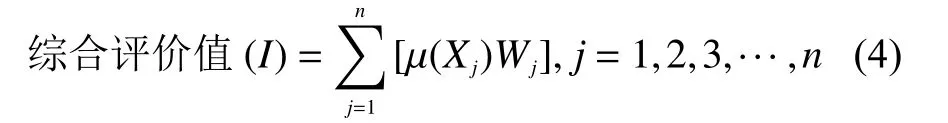
式中,I表示在低氮条件下各砧木耐低氮能力的综合评价值[26],Xj、Wj同上。
1.4 数据处理与分析
用Excel 2010和SAS 9.0进行数据处理分析及图表制作。使用单因素方差分析(One-way ANOVA)比较样本间的差异,对差异显著者采用Duncan’s多重检验(P<0.05为显著)比较,使用类平均法进行聚类分析。
2 结果与分析
2.1 不同供氮条件对苹果砧木生长的影响
在正常供氮条件下,7种苹果砧木的新梢生长速率大小依次为Nic29>T337>71-3-150>B9>山定子>Pajam2>青砧2号(图1-A),由此可知,Nic29的新梢长势强于山定子、青砧2号和其余矮化砧。与正常供氮相比,低氮条件下,T337、Nic29和山定子的新稍生长均受到显著抑制;B9、Pajam2和青砧2号的新稍生长未受显著影响;而71-3-150的新稍生长速率增幅最大,其新稍生长速率显著高于其余砧木。由图1-B可知,在正常供氮条件下,7种苹果砧木中Pajam2的平均叶面积最大,而71-3-150最小。与正常供氮相比,低氮条件下,Pajam2和山定子的叶面积显著减小,而T337、B9、71-3-150显著增大。
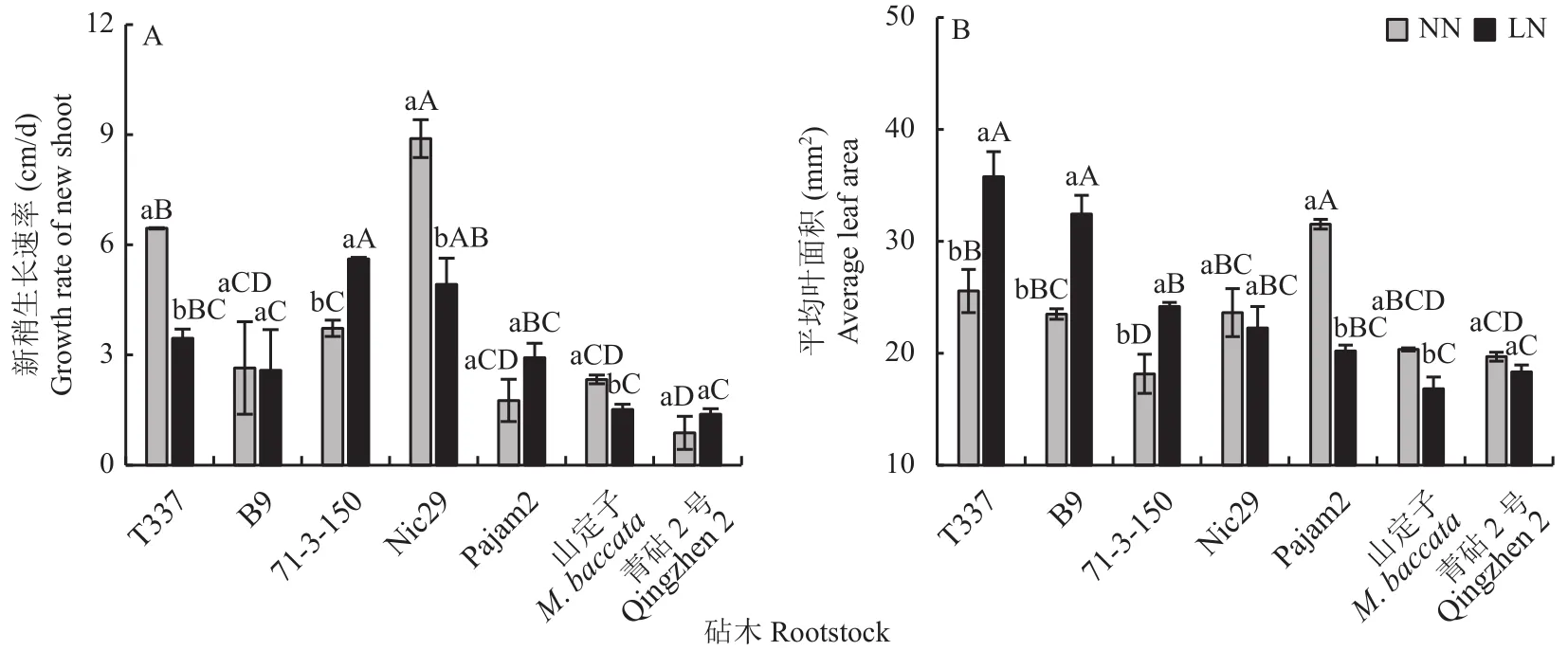
图1 不同供氮条件下苹果砧木新梢生长和平均叶面积的差异Fig.1 Shoot growth and leaf area of apple rootstocks under different nitrogen supply conditions
2.2 不同供氮条件对苹果砧木物质积累和根冠比的影响
由图2-A可知,在正常供氮条件下,乔化砧山定子和半矮化砧青砧2号的植株总干重和根冠比显著高于5种矮化砧;矮化砧中Pajam2的干物质积累量最大,B9的根冠比最高。与正常供氮相比,低氮条件下,T337和B9的植株总干重显著降低,其中T337的植株叶片、茎干和根系的干重均显著降低(图2-C),根冠比亦显著减小;其余砧木根冠比均呈上升趋势,其中B9、71-3-150和Nic29的增幅显著,三者的根冠比分别增长为对照的1.64、2.59和1.34 倍 (图2-B)。
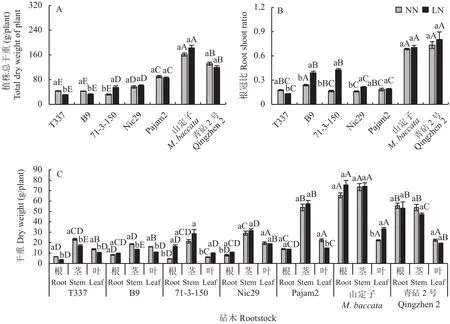
图2 不同供氮条件下苹果砧木干重及根冠比Fig.2 Dry weight and ratio of root/shoot of apple rootstocks under different nitrogen supply conditions
2.3 不同供氮条件下苹果砧木根系形态变化
在正常供氮条件下,乔化砧山定子的根尖数和根系分枝数均显著高于青砧2号和5种矮化砧,T337的根系总表面积和总根长显著高于其余矮化砧(B9、71-3-150、Nic29和Pajam2)。低氮条件下,不同苹果砧木根系形态的变化趋势存在显著差异。如表1所示,与正常供氮相比,低氮条件下,71-3-150和Nic29的根系总表面积均显著升高,此外,71-3-150的总根长、根系总体积、根尖数和根系分枝数等增幅显著。与正常供氮相比,低氮处理使得T337、B9、Pajam2和山定子根系总表面积和总根长均显著降低,T337的降幅最大,其根系总表面积和总根长分别降低了86.04%和88.57%。
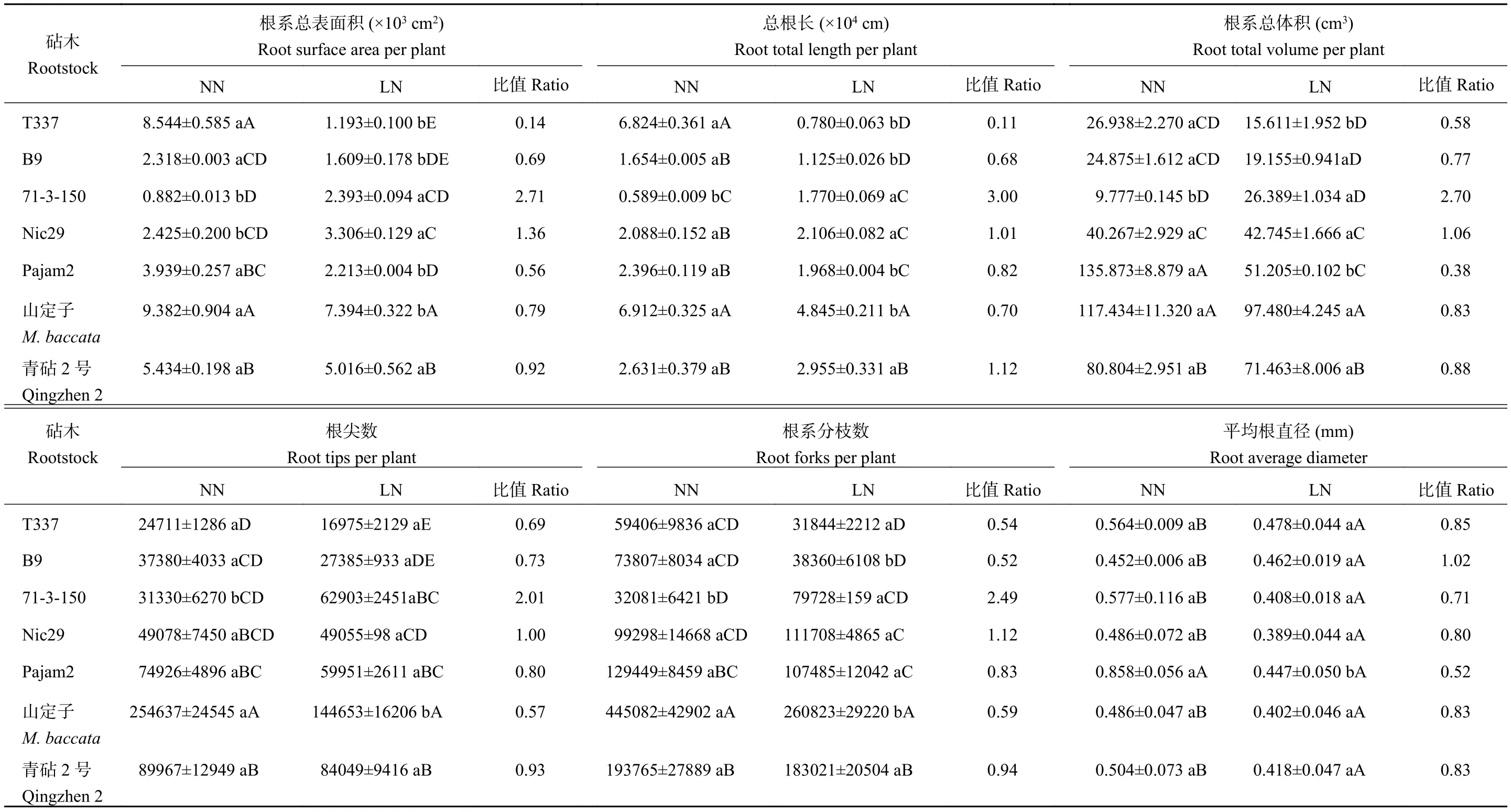
表1 苹果砧木根系形态及低氮处理相较于正常氮处理的比值Table 1 Root morphological characteristics and the ratio of apple rootstocks under low N supply relative to normal N supply condition
2.4 不同供氮条件对苹果砧木硝酸还原酶活性的影响
由图3-A可知,在正常供氮条件下,7种苹果砧木的叶片硝酸还原酶(NR)活性大小依次为Nic29>Pajam2> B9、山定子>71-3-150>青砧 2 号>T337;由图3-B可知,青砧2号的根系NR活性显著高于其余6种砧木,而矮化砧中,T337和71-3-150的根系NR活性显著高于B9、Nic29和Pajam2。与正常供氮相比,低氮条件下,7种苹果砧木叶片及根系中NR活性均发生显著改变。与正常供氮相比,低氮条件下7种砧木叶片NR活性均显著降低,其中,Pajam2和山定子降幅较大,分别降低了67.21%和64.52%。低氮胁迫对T337和71-3-150根系NR活性的影响不明显,却使青砧2号的根系NR活性降低了74.60%;低氮条件下,B9、Nic29、Pajam2和山定子根系NR活性分别较正常供氮提高了3.70、5.16、2.85和5.14倍。
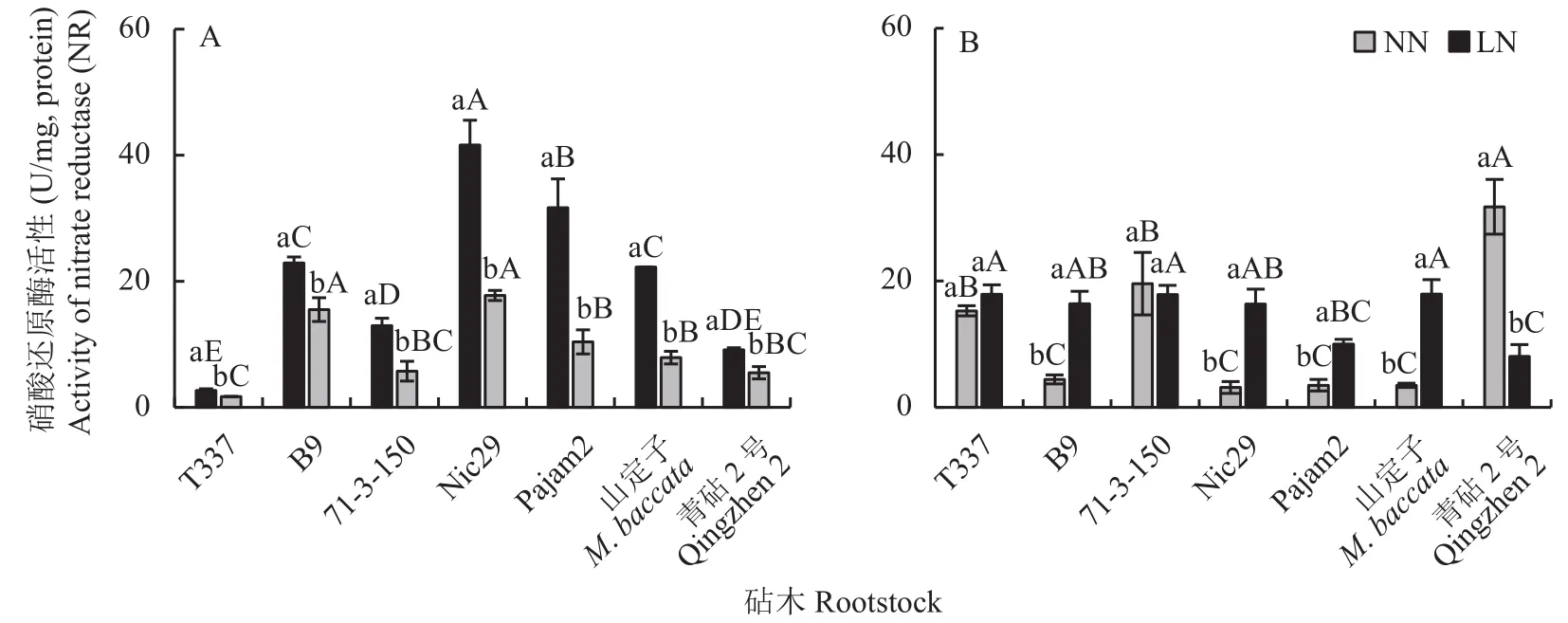
图3 不同供氮条件下苹果砧木叶片(A)及根系(B)硝酸还原酶(NR)活性Fig.3 Activities of nitrate reductase (NR) in leaves (A) and roots (B) of apple rootstocks under different nitrogen supply conditions
2.5 不同供氮条件对苹果砧木氮吸收利用的影响
由图4-A可知,在正常供氮条件下,青砧2号的植株总氮累积量显著高于山定子和矮化砧。与正常供氮相比,低氮条件下7种苹果砧木植株氮累积量均发生变化,71-3-150的植株氮累积量显著升高,增幅为66.26%;低氮胁迫使T337、B9、Nic29、Pajam2、山定子和青砧2号的植株氮累积量降低,青砧2号的植株氮累积量降幅最大。由图4-B可知,在正常供氮条件下,山定子的氮利用效率高于其余砧木,71-3-150和Pajam2的氮利用效率显著高于青砧2号、T337、B9和Nic29。与正常供氮相比,低氮条件下,B9、Nic29、Pajam2和青砧2号的氮利用效率均显著提高,青砧2号增幅最大。此外,低氮条件下,山定子和Pajam2的氮利用效率显著高于其余5种砧木。分析图4-C可知,与正常供氮相比,低氮条件下T337、B9和Pajam2的叶片氮累积量显著降低,T337和Pajam2的根系氮累积量显著降低,B9、Nic29和青砧2号的茎干氮累积量也显著降低,而71-3-150的根系和叶片的氮累积量显著升高。
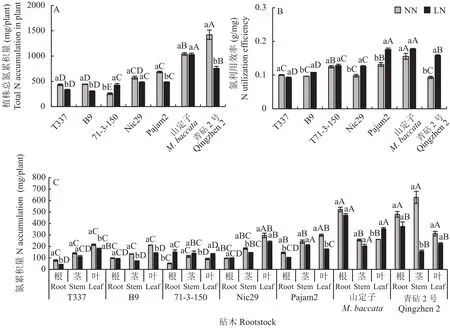
图4 不同供氮条件下苹果砧木氮累积量和氮利用效率Fig.4 Nitrogen accumulation and nitrogen use efficiency in apple rootstocks under different nitrogen supply conditions
2.6 苹果砧木耐低氮能力综合评价
以上述各指标的相对值为变量,利用模糊隶属函数对7种苹果砧木的耐低氮能力进行综合评价,结果表明,7种苹果砧木耐低氮胁迫能力依次为71-3-150>Nic29>山定子>B9>青砧2号>Pajam2>T337(表2)。以各砧木的耐低氮综合评价值为指标,采用类平均距离法可将7种苹果砧木聚类为3个类群,第Ⅰ类为耐性强的砧木,包括71-3-150;第Ⅱ类为耐性较弱的砧木,包括Nic29、山定子、B9和青砧2号;第Ⅲ类为耐性最弱的砧木,包括Pajam2和T337。
3 讨论
3.1 低氮胁迫对不同苹果砧木生长的影响
氮作为植物生长发育过程中的必需营养元素,是苹果器官分化、物质代谢及果实品质形成的重要物质基础。氮素亏缺条件下,树体会快速响应,植株生长和物质积累量等均发生显著变化,但受影响程度存在品种特异性[15,27]。在苹果栽培过程中,砧木作为树体的地下部分,负责吸收和转运土壤中的养分。优良砧木不仅是保障果树产量品质的基础,还能有效提高树体对多种生物及非生物胁迫的适应能力。通常认为在苹果栽培中,乔化砧树体高大且根系发达,对土壤适应能力强,而矮化砧木树体矮小且抗性较弱[28–29],这与本研究结果不一致。本研究对7种苹果砧木在低氮条件下的生长和生理生化指标进行分析,结合隶属函数法综合评价可知,71-3-150耐低氮胁迫能力最强,生产中常用的矮化砧木T337适应能力最弱,尽管在正常供氮条件下,乔化砧山定子在植株总干重、根系参数和植株氮累积量等方面明显高于矮化砧,但其耐低氮能力弱于71-3-150、Nic29等矮化砧(表2,图5)。程丽丽等[15]研究发现,低氮胁迫下光皮桦的根冠比增加,而低氮敏感型光皮桦植株生长和生物积累量等降幅大于耐受型。陈凌等[21]研究也发现,低氮胁迫下,耐低氮能力强的糜子品种表型性状变化幅度较小,且更容易调节自身生理特性,适应低氮环境。本研究与上述研究结果一致,与正常供氮相比,低氮胁迫下T337、Nic29和山定子的新稍生长受到显著抑制,Pajam2和山定子的叶片伸展减弱,植株地上部和根系干重降低,而71-3-150的新稍生长速率和地上部干重受影响较小,叶面积显著增大,且根系中干物质积累量增加,植株根冠比随之升高(图1,图2),表现出较强的低氮适应能力。韩清泉等[30]研究发现,低氮条件下仅耐低氮能力强的川滇柳根冠比升高,其余品种根冠比无显著变化甚至降低,这与本研究结果一致,除71-3-150、B9和Nic29外,Pajam2、山定子和青砧2号的根冠比变化不显著,而T337则显著降低。
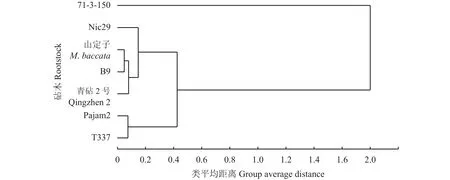
图5 苹果砧木的耐低氮胁迫能力聚类分析Fig.5 Cluster analysis of apple rootstocks tolerance to low N stress
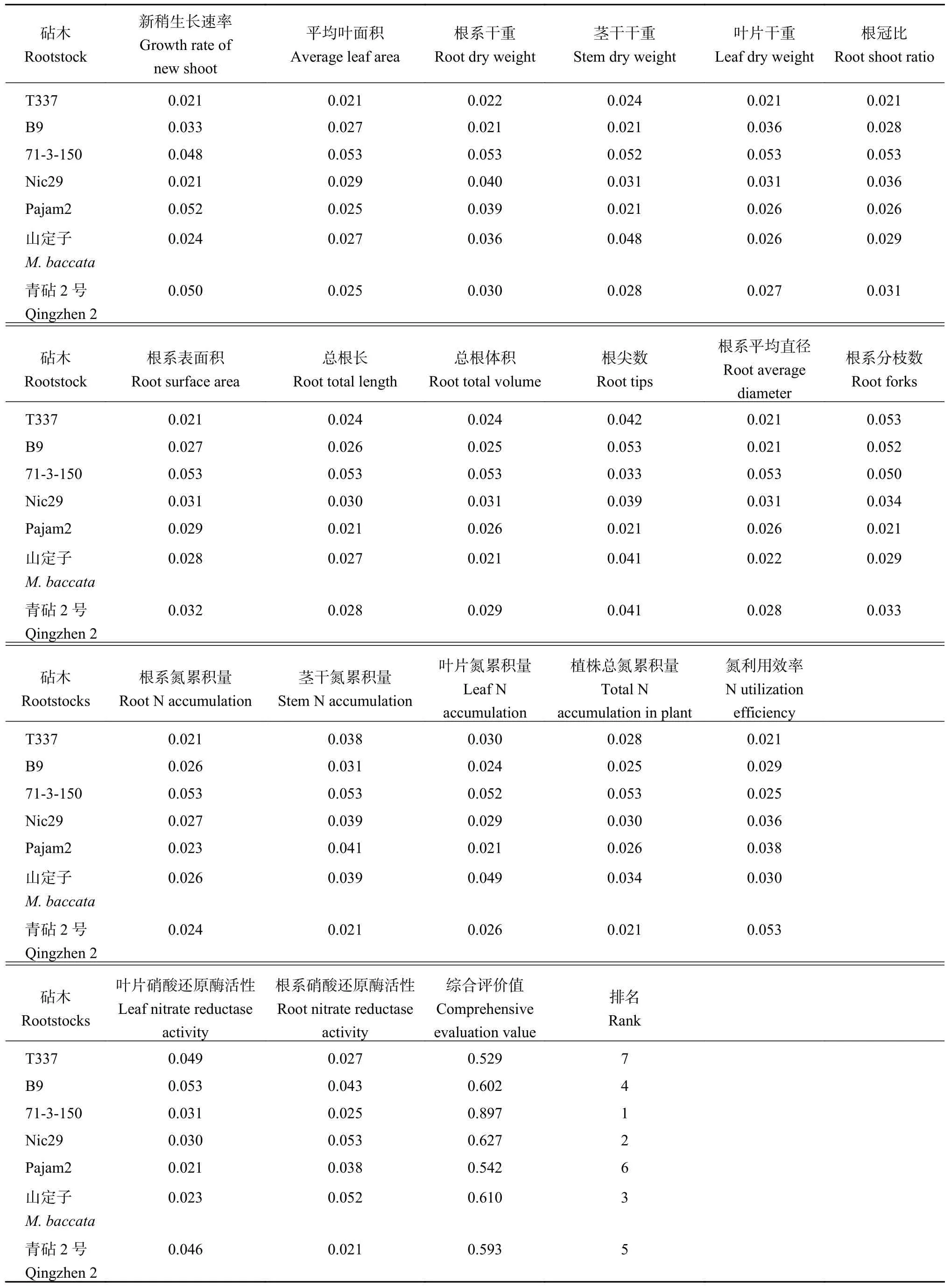
表2 苹果砧木对低氮胁迫耐受能力的模糊数学函数隶属法评价Table 2 Evaluation of resistance to low-N stress of apple rootstocks using the method of fuzzy membership function
3.2 低氮胁迫对不同苹果砧木根系形态的影响
根系是植物感知根际环境的器官,具有极强的可塑性以适应环境变化[25]。赵泽群等[10]在玉米的研究中发现,低氮条件下,玉米根干重的增加与总根长、根表面积、根体积和侧根数的增加密切相关,这与本研究结果一致。本研究发现,低氮胁迫下71-3-150的根系干物质积累量增加,根系总表面积、总根长、根系总体积、根尖数等根系性状增幅显著(表1),表明耐低氮能力强的砧木在低氮胁迫下能够促进根系生长,增大根系与土壤的接触面积,有利于植株对养分的吸收,并将更多的同化产物分配至根部,提高植株根冠比,这是砧木为适应氮素亏缺条件,捕获养分而产生的补偿机制之一[31]。不同苹果砧木根系对低氮胁迫的响应存在明显的品种特异性,低氮处理使得T337、B9、Pajam2和山定子根系总表面积和总根长均显著降低,耐低氮能力弱的砧木根系参数的降幅更大,而Nic29的根系总表面积、总根长、根体积和根系分枝数升高,但Nic29的根尖数减少,这可能与持续氮素亏缺过程中根基部细胞发生程序性死亡机制有关,而这一机制有待进一步研究[32]。
3.3 低氮胁迫对不同苹果砧木氮吸收利用的影响
关于植物氮高效吸收利用的研究主要集中在耐低氮性和氮效率两方面,前者关注植物在低氮条件下保持正常生长的能力,而后者是指在特定供氮条件下作物氮吸收利用状况[33]。根系吸收的氮素必须经同化后才能被植物利用,因此,氮代谢是植物生长发育过程中最重要的生理生化过程之一。硝酸还原酶(NR)作为一种诱导酶,是氮代谢过程中的关键酶,同时也是氮吸收的重要限制因子。正常供氮条件下,青砧2号的根系NR活性显著高于其他砧木(图3),有利于根系和植株的氮累积(图4)。李强等[34]研究发现,低氮胁迫下,不同氮效率的玉米叶片中NR活性均大幅下降,氮高效品种降幅小于氮低效品种,这与本研究结果一致,低氮胁迫使得不同苹果砧木叶片NR显著降低,Pajam2的降幅最大(图3-A)。Nic29、Pajam2 和 T337 均为 M9 (英国东茂林实验站收集和育成)中筛选到的优系,Pajam2和T337对低氮胁迫适应能力均低于71-3-150、B9等其他矮化砧,而Nic29表现出一定的优势。秦永梅等[35]研究发现,氮肥用量的增加可提高玉米叶片NR活性,但高水平的NO3–却使得根系NR降低,即叶片和根系表现出相反的变化趋势,这可能与植物体内氮的转运分配有关。彭玲等[1]在对不同稳定性供氮条件下T337的NR活性研究后发现,由于根系和叶片的所处环境不同,二者NR活性变化趋势存在差异。本研究也获得相似结果,低氮条件下B9、Nic29、Pajam2和山定子叶片NR活性降低,根系中NR活性显著提升。
NO3–被植物体吸收后,既可以在根系中同化利用,也可以转移到根系液泡中储存起来,被吸收的NO3–还可以通过木质部运往植物的地上部分[11]。发达的根系是氮高效吸收利用的基础,正常供氮条件下,山定子和青砧2号的植株氮累积量明显高于矮化砧;此外,多数研究认为,低氮条件下植株总氮累积量与植株根系形态密切相关[36–37],这与本研究结果一致。低氮胁迫使T337、B9、Pajam2和青砧2号等苹果砧木根系发育受阻,植株及各器官的氮累积量亦呈降低趋势,而71-3-150的根系干物质量和根系参数均显著提高,植株及各器官的氮累积量增加。但钟思荣等[38]研究发现,低氮条件下,烟草根系体积增大,但茎叶氮累积量减小,表明氮累积量不仅依赖于根系,也与植物自身的氮素吸收能力有关。这与本研究结果一致,低氮条件下,Nic29的根系总表面积和总体积高于正常供氮,根系中氮累积量未有明显变化,但叶片和茎干中氮累积量却呈降低趋势。除了氮吸收能力,秦璐等[25]研究发现,低氮胁迫会使油菜的氮利用效率显著增加,这与苹果砧木的变化规律一致,这是植物为适应低氮环境而产生的重要生理应答机制之一。
4 结论
在正常供氮条件下,乔化砧木山定子和半矮化砧青砧2号在植株生长、根系发育和养分吸收利用等方面均强于矮化砧;低氮条件下,各砧木均发生显著变化且变化趋势存在差异。基于各指标的耐低氮指数,利用隶属函数法和聚类分析可将砧木分为3种耐性类型:第Ⅰ类为耐性强的砧木(71-3-150);第Ⅱ类为耐性较弱的砧木(Nic29、山定子、B9和青砧2号);第Ⅲ类为耐性最弱的砧木(Pajam2和T337)。低氮条件下,苹果砧木通过提高氮利用效率适应养分亏缺,其中,耐性强的砧木植株生长受抑制程度较小,并通过调节自身生理特性,增加根系中的物质和养分积累,提高植株根冠比,以适应低氮环境。
猜你喜欢
中国果树(2022年5期)2022-06-09
中国特种设备安全(2022年1期)2022-04-26
草业科学(2022年2期)2022-03-21
防爆电机(2021年6期)2022-01-17
防爆电机(2021年3期)2021-07-21
防爆电机(2021年1期)2021-03-29
落叶果树(2021年6期)2021-02-12
中国科技纵横(2018年14期)2018-08-30
现代农业科技(2017年16期)2017-09-22
新农业(2017年3期)2017-05-17
