
宽体金线蛭WpE基因的克隆及生物信息学分析
2022-07-14 05:41程搏幸卢昱希匡诗涛吴正关田青青邵贵燕
贵州师范学院学报 2022年6期
程搏幸,卢昱希,匡诗涛,吴正关,田青青,邵贵燕,刘 飞*
(1. 贵州师范学院生物科学学院,贵州 贵阳 550018;2. 贵州省农业科学院畜牧兽医研究所,贵州 贵阳 550002;3. 盐城工学院海洋与生物工程学院,江苏 盐城 224051)
0 引言
宽体金线蛭(Whitmaniapigra)俗称为蚂蟥,是药材水蛭基原动物之一,收录于《中国药典》(2020年版),具有活血通经的功效[1],在抗凝血、抗炎、溶解血栓等方面有显著疗效[2]。目前,从宽体金线蛭体内分离或筛选得到anti-THR、Pigrin[3]、WP-30[4]、WPI01[5]、WA3-1、Whittide[6]等具有抗凝血或溶解血栓活性的蛋白质。
Eglin C属于马铃薯抑制剂Ⅰ(Potato inhibitor Ⅰ,PI-Ⅰ)家族,是丝氨酸蛋白酶抑制剂众多家族中的一员,具有抑制胰凝乳蛋白酶的功能[7]。1977年,从欧洲医蛭(Hirudomedicinails)中分离纯化得到Eglin C,能抑制组织蛋白酶G、弹性蛋白酶、胰凝乳蛋白酶和枯草杆菌蛋白酶等的蛋白活性,具有止血、抗炎等功效[8]。
目前,关于宽体金线蛭的研究主要集中在动物养殖[9]及其抗凝等药用价值[10]方面,而对于宽体金线蛭体内具有抗炎生物活性成分的筛选及其作用机理的研究较为鲜见。从宽体金线蛭唾液腺转录组数据库(未发表)中筛选到匹配EglinC基因的转录本,克隆宽体金线蛭WpE基因,对该基因进行生物信息学分析,旨在为深入研究WpE基因功能和进一步开发利用宽体金线蛭提供科学依据。
1 材料与试剂
1.1 实验材料
所用材料来自江苏省宝应县,经作者鉴定为宽体金线蛭(W.Pigra)。选择健康、体表无伤痕的宽体金线蛭作为研究对象,冰上无菌解剖后收集其唾液腺,置于-80℃的冰箱中保存。
1.2 试剂
RNAiso Plus总RNA提取试剂、PrimeScriptTMII 1st Strand cDNA Synthesis Kit均购自大连TaKaRa有限公司,琼脂糖凝胶DNA回收试剂盒购于上海生工生物工程股份有限公司,2×Taq Master Mix购于南京诺唯赞生物科技股份有限公司,pEASY-Blunt Cloning Kit平端克隆载体试剂盒购于北京全式金生物技术有限公司,大肠杆菌DH5α感受态细胞购于北京天根生化科技有限公司,克隆引物与测序技术服务由上海生工生物工程股份有限公司提供。
2 实验方法
2.1 宽体金线蛭总RNA提取及cDNA合成
取宽体金线蛭唾液腺放入经液氮预冷的研钵中研磨,研磨期间不断补充液氮,待样品呈粉末状后转入1.5 mL离心管中,按照TaKaRa总RNA提取试剂盒的使用说明书提取宽体金线蛭唾液腺的总RNA,通过1.5%琼脂糖凝胶电泳检测RNA完整性。
以质量合格的唾液腺总RNA为模板,参照PrimeScriptTMII 1st Strand cDNA Synthesis Kit试剂盒说明书对RNA进行逆转录合成cDNA:取上一步提取的RNA 5.0 μL,按顺序加入dNTP Mixture 1.0 μL、Random 6 mers 1.0 μL、补充RNase free H2O使反应体系总体积达10.0 μL,65℃反应5 min后置于冰上冷却;再加入5×PrimeScript Ⅱ Buffer 4.0 μL、RNase Inhibitor 0.5 μL、PrimeScript II RTase PrimeScript II 1.0 μL、RNase free H2O,混匀后30℃反应10 min,95℃反应5 min,即可得到cDNA,保存于-20℃备用。
2.2 宽体金线蛭(WpE)基因的克隆
根据宽体金线蛭转录组数据库获得的WpE基因转录本序列(DN14831-c1-g1),使用Primer Premier 5.0软件设计特异性引物,上游引物WpE-F序列为5’-TTCTGAAAAATTTCATTCATTT CTC-3’,下游引物WpE-R序列为5’-ATTTTCTTCTCAGATTTCAGAAGTTC-3’,克隆产物大小为340 bp。利用2×Taq Master Mix对cDNA进行PCR扩增,扩增体系总体积50 μL,包括2×Taq Master Mix 25 μL,上游引物和下游引物各1 μL,gDNA 1 μL,补充ddH2O至50 μL。PCR反应程序为94℃预变性5 min;95℃变性15 s、50℃退火20 s、72℃延伸40 s,共40个循环;最后72℃保持5 min。使用1.5%琼脂糖凝胶电泳检测扩增产物,使用DNA凝胶回收试剂盒回收目的条带。
将胶回收纯化的目的基因按照pEASY-Blunt Cloning Kit平端克隆载体试剂盒说明书连接到pEASY-Blunt质粒上,将连接产物转入大肠杆菌DH5α感受态细胞,培养过夜后挑取阳性单克隆菌落扩大培养并进行菌液PCR,反应条件同上,使用琼脂糖凝胶电泳验证合格后送上海生工生物工程股份有限公司进行测序。
2.3 宽体金线蛭WpE基因序列的生物信息学分析
将测序得到的WpE基因序列通过DNAMAN 8.0软件翻译成氨基酸序列,使用NCBI在线网站中的ORF Finder工具(https: //www. ncbi. nlm. nih. gov/orffinder/)查找WpE基因的完整开放阅读框(Open Reading Frame)。利用SignalP 5.0在线网站对WpE蛋白进行信号肽分析。使用Cell-PLoc 2.0在线网站(http: //www. csbio. sjtu. edu. cn/bioinf/Cell-PLoc-2/)进行亚细胞定位分析。通过Expasy在线网站中的Protparam工具(https: //web. Expasy. org /protparam/)分析WpE蛋白去除信号肽后的理化性质。通过Expasy在线网站中的ProtScale工具(https: //web. Expasy. org /protscale/)进行WpE蛋白亲/疏水性分析。通过NetPhos 3.1 Server在线网站(http: //www. cbs. dtu. dk/services/NetPhos/)进行磷酸化位点分析。通过TMHMM Server 2.0在线网站(http: //www. cbs. dtu. dk/services/TMHMM-2.0/)预测跨膜结构域。通过Inter Pro Scan在线网站(https: //www. ebi. ac. uk/interpro/search/sequence/)预测保守结构域。根据NCBI BLASTP(https: //blast. ncbi. nlm. nih. gov/Blast. cgi? PROGRAM=blastp &PAGE_TYPE= BlastSearch&LINK_LOC=blasthome)的比对结果,选取同源性较高的氨基酸序列,利用Jalview 2.11.1.4软件进行氨基酸多重序列比对,使用MEME在线网站(https: //meme-suite. org/meme/tools/meme)进行氨基酸保守基序分析,最后使用MEGA 11.0.8软件采用邻接法构建系统进化树。使用SOPMA在线网站(https: //npsa-prabi. ibcp. fr/cgi-bin/npsa_automat. pl? page=npsa_sopma. html)进行二级结构预测,使用PyMOL 2.5.2软件进行三级结构建模。
3 结果与分析
3.1 宽体金线蛭WpE基因的克隆
以宽体金线蛭唾液腺cDNA为模板,利用特异性引物WpE-F、WpE-R进行PCR扩增,扩增产物经琼脂糖凝胶电泳检测,发现在目标产物大小附近出现特异性条带,测序结果与转录本碱基序列一致(图1),并将该基因命名为WpEglin(WpE,GenBank: OL598605)。
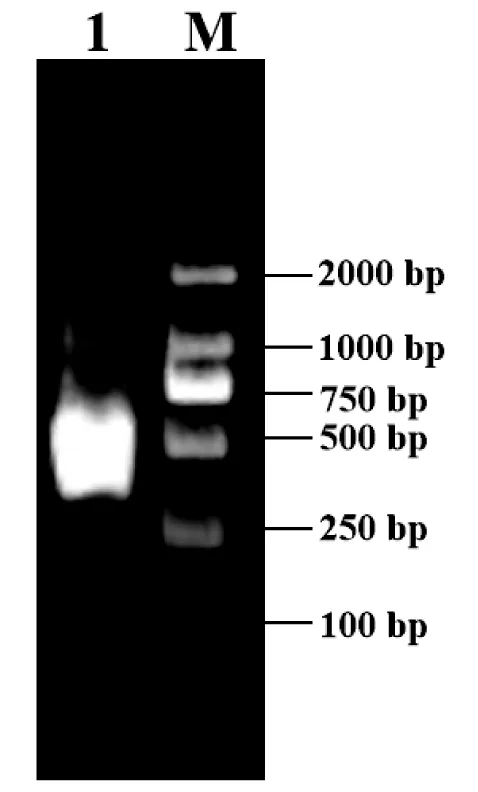
图1 宽体金线蛭WpE基因扩增结果
3.2 宽体金线蛭WpE基因核酸序列分析
WpE基因的完整开放阅读框ORF大小为258 bp,编码氨基酸85 aa,包含16个氨基酸组成的信号肽和69个氨基酸组成的成熟蛋白序列(图2),属于分泌蛋白,定位于细胞外。

图2 宽体金线蛭WpE蛋白质开放阅读框(ORF)及其氨基酸序列
3.3 宽体金线蛭WpE基因编码蛋白理化性质分析
除去WpE蛋白信号肽后,成熟WpE蛋白氨基酸序列蛋白质分子式为C374H564N100O102,原子总数为1140,相对分子质量为8.093 kDa,理论等电点(pI)为5.85。该蛋白含有8个碱性氨基酸(Arg+Lys),10个酸性氨基酸(Asp+Glu),氨基酸含量最多的是缬氨酸(Val,13个,占18.84%),其次是苯丙氨酸(Phe,7个,占10.14%)。脂肪酸系数为92.90,不稳定指数为21.05,属于稳定蛋白。成熟WpE蛋白中第65个氨基酸(Val)处疏水性最强,最高值为2.144,亲水性最强出现在第25个氨基酸(Lys)处,最低值为-1.722,亲水性(GRAVY)总平均值为-0.129,属于亲水性蛋白。成熟WpE蛋白无跨膜结构域,不属于跨膜蛋白,含有1个磷酸化位点为酪氨酸(Tyr)。
3.4 宽体金线蛭WpE蛋白氨基酸多重序列比对
将WpE蛋白氨基酸序列在NCBI的BLASTP中进行在线对比,该序列与菲牛蛭(Poecilobdellamanillensis)、欧洲医蛭(H.medicinalis)蛋白质序列的同源性较高,相似度达60%以上。选取同源性较高的菲牛蛭(P.manillensis)、欧洲医蛭(H.medicinalis)、普通小麦(Triticumaestivum)、乌拉图尔小麦(Triticumurartu)、杏(Prunusarmeniaca)、蒺藜苜蓿(Medicagotruncatula)等物种胰凝乳蛋白酶抑制剂氨基酸序列,用Jalview 2.11.1.4软件进行氨基酸多重序列比对,结果表明WpE蛋白氨基酸序列具有PI-Ⅰ家族活性位点Asp46,及能够相互作用形成氢键网络或盐桥从而稳定构象的Thr44、Asp46、Arg51、Arg53和Gly70等位点(见图3)。
利用MEME在线网站分析以上物种的氨基酸保守基序(motif),识别到3个共有的保守序列(见图4),这些保守序列可能为蛋白的活性结合位点。

图3 宽体金线蛭WpE蛋白氨基酸序列与其他物种的多重序列比对
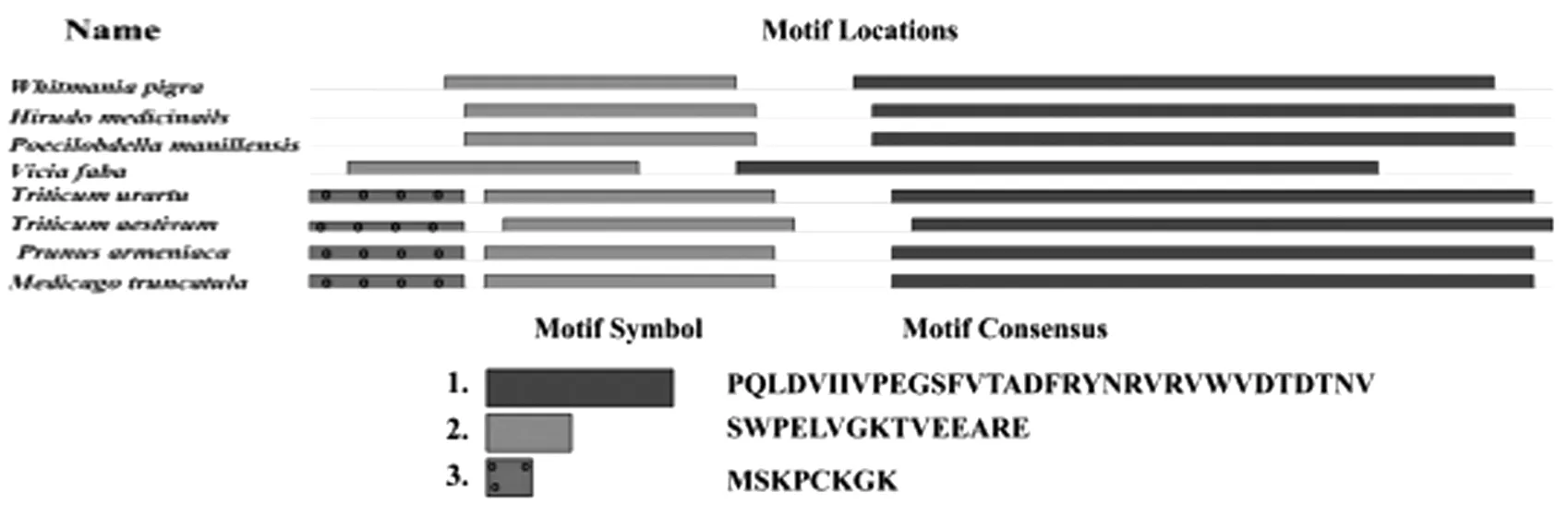
图4 宽体金线蛭WpE蛋白氨基酸序列与其他物种的保守基序分析
3.5 宽体金线蛭WpE基因系统进化树分析
在NCBI中下载已报道的菲牛蛭(P.manillensis)、欧洲医蛭(H.medicinalis)、蛭态轮虫(Rotariasordida)、虾夷扇贝(Mizuhopectenyessoensis)、欧洲扇贝(Pectenmaximus)、紫贻贝(Mytilusedulis)、普通小麦(T.aestivum)、乌拉图尔小麦(T.urartu)、枯草芽孢杆菌(Bacillussubtilis)、地衣芽孢杆菌(Bacilluslicheniformis)等物种胰凝乳蛋白酶抑制剂氨基酸序列,利用MEGA 11.0.8软件中的Neighbor-joining法构建系统进化树,BOOTSTRAP值为500,结果表明宽体金线蛭WpE蛋白氨基酸序列与菲牛蛭、欧洲医蛭亲缘关系较近,聚为一支,且遗传距离与海蠕虫(Capitellateleta)、海蜗牛(Aplysiacalifornica)等无脊椎动物较近,与传统分类学一致(图5)。
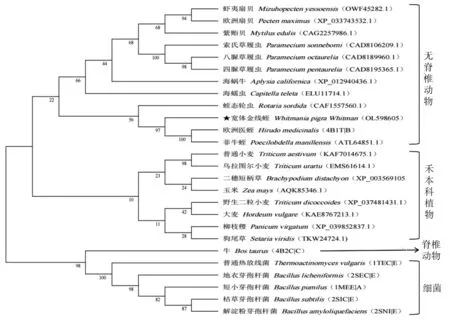
图5 宽体金线蛭WpE蛋白氨基酸序列系统进化分析
3.6 宽体金线蛭成熟WpE蛋白二级、三级结构预测
利用SOPMA在线网站对WpE编码蛋白的二级结构进行分析,结果表明WpE编码蛋白的二级结构包含24.64%α-螺旋、15.94% β-折叠、26.09%延伸链和33.33%无规则卷曲(图6)。

图6 宽体金线蛭WpE蛋白二级结构预测
采用同源建模的方法,在SWISS-MODEL在线网站中预测WpE编码蛋白的三维结构,并使用Pymol 2.5.2软件对其进行优化处理。以欧洲医蛭Eglin C蛋白(GenBank: 4B1T_B)为模板,建模覆盖范围为2~69位氨基酸残基,序列同源性为63.24%,且在WpE蛋白与欧洲医蛭Eglin C蛋白的活性位点处都具有相似的环状结构(图7)。
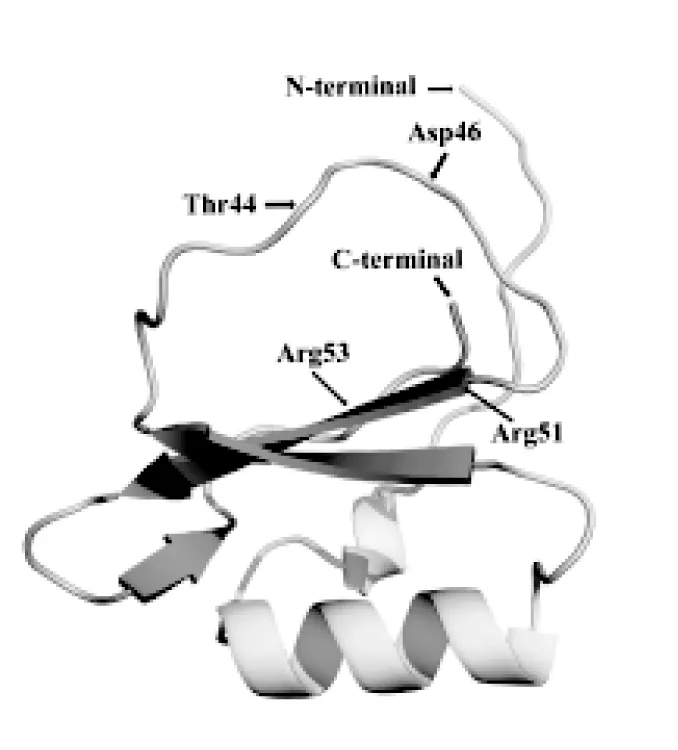
图7 宽体金线蛭WpE蛋白三级结构预测
4 讨论
本研究从宽体金线蛭唾液腺转录组数据库筛选转录本DN14831-c1-g1注释到欧洲医蛭EglinC基因,通过基因克隆的验证,且测序结果与转录本碱基序列一致。该基因编码85个氨基酸,其中N端含有16 个氨基酸组成的信号肽和69 个氨基酸组成的成熟蛋白序列。在欧洲医蛭、菲牛蛭等吸血类水蛭中已鉴定到了EglinC基因,该基因编码的蛋白质具有抑制胰凝乳蛋白酶等蛋白质水解酶的活性,具有潜在消除炎症、抑制血液凝固等功效。
欧洲医蛭Eglin C蛋白包含四个平行或反向平行的β折叠,在蛋白的侧面含有一个α螺旋,而另一面有一个延伸的环状活性位点[11],该环状结构与目标蛋白酶活性位点结合,WpE蛋白也有类似的结构(图7),推测WpE蛋白与欧洲医蛭Eglin C蛋白具有相似功能。Eglin C蛋白环状结构上的Leu45-Asp46位点直接参与蛋白酶活性抑制[12],其中Asp46相对保守,如突变后,Eglin C蛋白对丝氨酸蛋白酶的抑制作用显著下降[11],表明该位点至关重要。而Leu45相对可变,在PI-Ⅰ家族中该氨基酸位点还出现过Ala、Met等疏水性氨基酸残基(见图3),不影响蛋白抑制剂活性[13]。WpE蛋白中含有保守的Ala45-Asp46活性位点,虽然宽体金线蛭中的Ala45与吸血类的欧洲医蛭、菲牛蛭中Leu45有所不同,但其它一些物种中PI-Ⅰ家族蛋白具有Ala45,如红豆等[13],因此推测WpE蛋白该位点与其它水蛭的不同并不影响WpE蛋白活性。与其它丝氨酸蛋白酶抑制剂不同,PI-Ⅰ家族蛋白含有两个突出核心精氨酸(Arg5l,Arg53),它们可形成一种非共价相互作用,能促进蛋白中环状结合位点构象的稳定,如定点突变该两位点精氨酸,蛋白活性将受到影响[11]。WpE蛋白与Eglin C中均缺少二硫键,研究表明这并不影响Eglin C对酸变性、热变性的高度耐受[14]。根据上述分析,WpE蛋白与Eglin C蛋白结构相似,具有相似的活性位点,推测WpE蛋白具有与Eglin C类似的胰凝乳蛋白酶抑制剂活性。
Eglin C蛋白能抑制中性粒细胞组织蛋白酶G,组织蛋白酶G能够活化巨噬细胞,促进血小板聚集以加快血液凝固[15],因此,Eglin C蛋白具有抑制血液凝固的功效。此外,粒细胞蛋白酶的过量释放可能会增强炎症[16],Eglin C蛋白能通过抑制粒细胞蛋白酶活性,抑制炎症的形成。结合WpE蛋白与Eglin C蛋白结构的分析,宽体金线蛭中的WpE蛋白很可能也具有上述抗凝血和抑制炎症的功能,但两者功效的差异有待进一步研究。宽体金线蛭摄食时将具有抗凝血和抑制炎症功能的WpE蛋白注入宿主体内,可能是其顺利完成摄食的保障。
综上所述,本研究对WpE基因结构、WpE蛋白功能通过生物信息学方法进行了初步分析,为探究宽体金线蛭抗炎、抗凝血的分子机制提供理论依据,同时为今后进一步合理开发与利用宽体金线蛭奠定了理论基础。
猜你喜欢
小学生作文(低年级适用)(2022年11期)2022-12-02
中国洗涤用品工业(2019年4期)2019-05-11
文苑(2018年22期)2018-11-19
中成药(2018年10期)2018-10-26
中成药(2018年1期)2018-02-02
中成药(2017年7期)2017-11-22
快乐语文(2017年30期)2017-11-15
中成药(2017年3期)2017-05-17
中国洗涤用品工业(2017年2期)2017-04-16
中国粮油学报(2016年5期)2016-01-23
