
延迟交配对橘小实蝇繁殖适度的影响
2022-07-14 03:41易小龙董子舒郑光楠王小云郑霞林
环境昆虫学报 2022年3期
易小龙,董子舒,郑光楠,王小云,郑霞林,陆 温
(广西大学农学院,广西农业环境与农产品安全重点实验室,南宁 530004)
橘小实蝇Bactroceradorsalis(Hendel)隶属于双翅目实蝇科,能为害250多种水果及蔬菜(Clarkeetal., 2005),还可在不同寄主间转移为害(段云博, 2018; 巫辅民, 2019; 李媛, 2020)。目前,橘小实蝇已广泛分布于广西、广东、福建、海南、湖南、贵州、云南、四川、重庆等多个省市和自治区,给我国水果产业造成了极大损失,严重制约了我国水果产业的发展(朱雁飞等, 2020)。根据橘小实蝇的发生特点,当前防治主要集中在成虫期,如雄性不育技术(梁广勤等, 2008; Sohailetal., 2019)、蛋白饵剂食诱(全金成等, 2017; 孙太华等, 2020)和性诱(薛超等, 2015; 赵菊鹏等, 2017)等。
延迟交配是指动物的交配行为在自然界中受到各种因素的影响,不能在正常时间进行交配而延迟(Chiltonetal., 1993)。延迟交配对昆虫的繁殖适度有显著影响(如交配成功率、产卵量、孵化率和寿命等),且其影响因昆虫种类不同而异。例如,延迟交配会降低斜纹夜蛾Spodopteralitura产卵量和孵化率,且显著延长其寿命(Wuetal., 2018);延迟交配会降低朱红毛斑蛾Phaudaflammans的交配率、产卵量和孵化率,但对其寿命无显著性影响(Zhengetal., 2020)。基于延迟交配能降低昆虫繁殖适度这一现象,利用性信息素在野外干扰昆虫交配已在梨小食心虫Grapholithamolesta(郑鹏华等, 2020; 郑燕等, 2020)、苹小卷叶蛾Adoxophyesoranabeijingensis(涂洪涛等, 2018)和苹果蠹蛾Cydiapomonella(张煜等, 2017; 崔笑雄等, 2020)等害虫的防治上得到应用,并取得了显著防效。
性信息素“迷向法”是指利用合成性信息素扰乱和阻断雌、雄虫之间的化学通讯(孟宪佐和魏康年, 1981)。干扰昆虫交配以延迟成虫交配日龄降低其繁殖适度,是间接控制害虫种群的一种方法。目前,延迟交配行为对蛾类害虫种群增长的负作用也已成为评价交配干扰技术有效性的指标之一,而其对实蝇类害虫的影响鲜见报道。橘小实蝇成虫的寿命相对一般蛾类较长,性成熟时间普遍在7~12日龄之间(林进添, 2004),延迟交配对橘小实蝇繁殖适度的影响,目前尚无报道。鉴于此,本文利用响应面分析法,研究了延迟交配对橘小实蝇繁殖适度的影响,以期为基于性信息素迷向法在橘小实蝇防控上的应用提供理论参考。
1 材料与方法
1.1 供试虫源
橘小实蝇初始虫源采集于广西玉林市玉州区(22.63N, 110.15E)番石榴果园,捡拾内具幼虫的番石榴虫果带回广西大学昆虫所养虫室饲养。室内饲养温度26℃±2℃,相对湿度70%±10%,光周期14 L ∶10 D,待成虫羽化后将其转移至用金属支架和尼龙网制作而成的养虫笼(长×宽×高=100 cm×100 cm×100 cm)中饲养。幼虫用香蕉Musanana饲养,成虫喂以人工饲料(酵母膏 ∶白糖=1 ∶4),并以润湿的脱脂棉供水,每隔2 d更换一次养虫笼中的饲料与水。本研究所用橘小实蝇皆为该种群的后代。
1.2 实验方法
设定羽化当天的橘小实蝇日龄为0日龄,羽化第2天的成虫日龄为1,依此类推。实验所用成虫均在3日龄之前将雌、雄分开,单性饲养,以确保其在实验之前未发生交配。实验在广西大学昆虫所养虫室进行,养虫室内温度为26℃±2℃,相对湿度为70%±10%,光周期14 L ∶10 D。
依据Expert Design 8.0中心复合设计法(Jimenez-Perez and Wang, 2003)设计了9个橘小实蝇不同成虫日龄配对组合,研究其对橘小实蝇繁殖适度的影响。具体方法是下午18时,室内灯光模拟成黄昏时的光照强度,将未交配过的雌、雄成虫各30头混合在同一养虫笼内,3次重复。然后,每20 min观察1次橘小实蝇的交配情况,如发现交配行为,待其交配稳定后,用50 mL离心管收集,并将其转移至产卵观测装置内(2 000 mL的塑料盒,内具饲料、水及用50 mL离心管制作而成的集卵器),观察时间直至当天20 ∶30,无新的交配现象出现后,关闭室内灯光。统计交配率。待橘小实蝇交配结束后,取出装置中的雄成虫,将20头交配后的雌虫单独饲养在产卵观测装置内,隔天更换水和饲料,每天更换产卵观测装置中的集卵器,在SM2800N尼康体视显微镜(尼康公司,东京,日本)下统计卵粒数量。随后将卵置于培养皿(直径9 cm)内待其孵化,3 d后记录各皿卵粒孵化数量,最后统计产卵量、孵化率和雌虫寿命。

图1 一种橘小实蝇成虫产卵量观测装置Fig.1 An observation device for fecundity of adult Bactrocera dorsarlis注:1,饲养盒;2,取虫口;3,食物盘;4,喂水槽;5,喂食槽;6,透气孔;7,集卵器;8,加厚垫;9,放虫口;10,封闭件;11,产卵孔。Note: 1, Feeding box; 2, Gaps of take insect out; 3, Feeding plate; 4, A tank for feeding water; 5, A tank for feeding food; 6, Air vent; 7, A device for collecting eggs; 8, Thickening mat; 9, Gaps of put insects in; 10, A sheet used to seal; 11, Ovipositional apertures.
1.3 数据统计
使用SPSS 22.0(IBM, 美国)进行数据统计分析。不同日龄雌、雄成虫的交配率采用单因素方差分析,并应用Tukey法进行多重比较(P<0.05)。由于雌虫的产卵量、卵的孵化率和雌虫的寿命等数据不满足正态分布,故采用非参数检验Kruskal-Wallis法进行差异显著性检验(P<0.05)。响应面实验设计与统计分析软件为Design Expert 8.0(Stat-Ease, 美国),采用Origin 8.0(Origin Lab, 美国)作图。
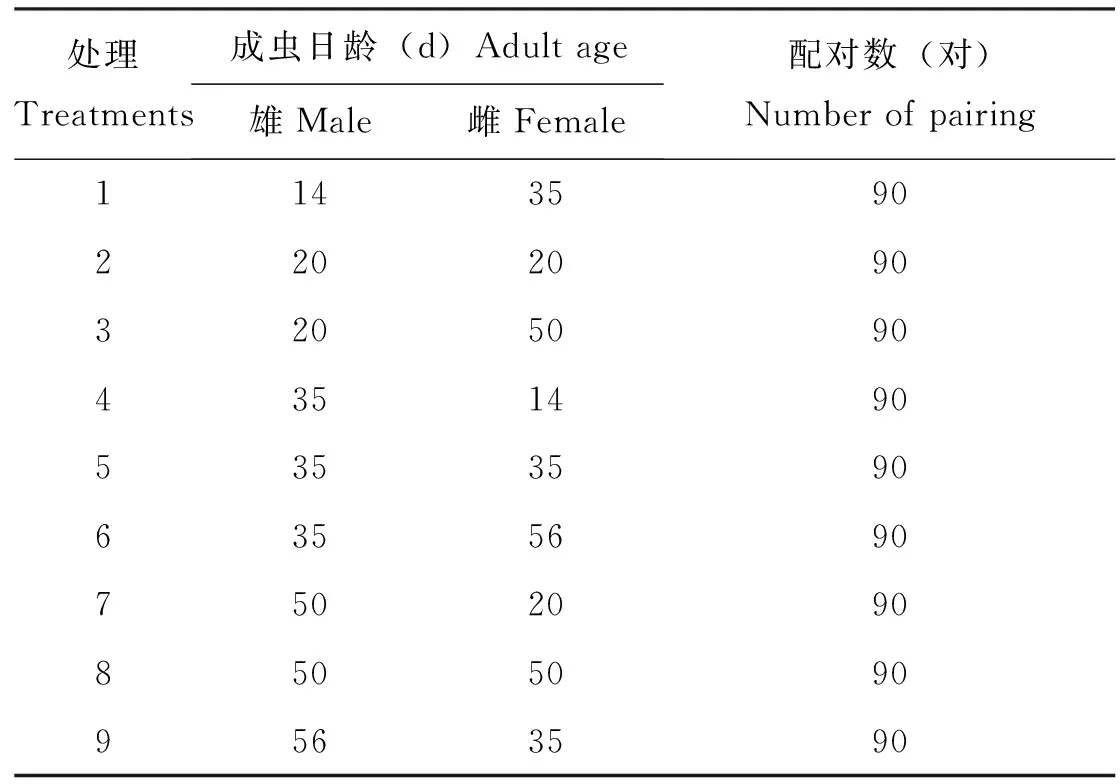
表1 橘小实蝇不同雌、雄成虫交配日龄组配(Expert Design 8.0设计)Table 1 Different mating age groups of male and female of Bactrocera dorsalis (Design by Expert Design 8.0)
2 结果与分析
2.1 延迟交配对橘小实蝇交配率的影响
交配日龄显著影响橘小实蝇的交配率(P<0.05)。35日龄雄虫与35日龄雌虫组合交配率最高,为81.00%±2.00%;35日龄雄虫与14日龄雌虫配对时,其交配率最低,为32.33%±2.33%,其余组合的交配率介于两者之间。其中,雄虫仅在14日龄未完全性成熟时,与35日龄雌虫配对的交配率显著低于35日龄雄虫与35日龄雌虫配对的交配率,其余日龄组合间无显著差异。相较而言,雌虫交配日龄的延迟对其交配率的影响更为显著,除了在未完全性成熟时会显著降低交配率外,其交配日龄的延迟也会对交配率造成显著的负面影响(表2)。
2.2 延迟交配对橘小实蝇产卵量和卵孵化率的影响
橘小实蝇雌虫的交配日龄对其产卵量存在显著影响,随雌虫交配日龄的增加,其产卵量降低;雌虫的交配日龄对卵的孵化率无显著影响。雄虫的交配日龄对卵孵化率有显著影响;随雄虫日龄的增加,卵孵化率下降。利用Design Expert软件中的中心复合设计,对不同年龄组合实验所得到的产卵量数据进行分析,结果表明,橘小实蝇雄虫交配日龄对产卵量有显著影响(P=0.023);雌虫交配日龄对产卵量的影响则极显著(P<0.001),且雌、雄虫交配日龄对雌虫产卵量不存在明显的交互作用。进一步求得产卵量(y)与雄虫日龄(x1)和雌虫日龄(x2)的回归方程为y=514.36+3.08x1-11.05x2,R2值为0.7930。综上所述,橘小实蝇雌虫产卵量的变化主要取决于雌虫交配日龄的变化,其产卵量随雌虫交配日龄的延迟而减少(表2,图2)。

表2 延迟交配对橘小实蝇繁殖适度的影响(n=20)Table 2 Effect of delayed mating on reproductive fitness in Bactrocera dorsalis

图2 交配日龄对橘小实蝇产卵量的影响Fig.2 Effect of female and male age on fecundity in Bactrocera dorsalis
2.3 延迟交配对橘小实蝇雌成虫寿命的影响
橘小实蝇雌、雄虫交配日龄的增加会显著延长雌虫的寿命。将不同年龄组合实验所得到的雌成虫寿命数据,代入中心复合设计,进一步分析了雌虫寿命与雌、雄虫交配日龄的关系。结果表明,橘小实蝇雌、雄成虫交配日龄对雌虫的寿命均有极显著的影响(雄虫,P<0.001;雌虫,P<0.001),雌成虫寿命随雌虫或雄虫交配日龄的延迟而增加,雌、雄成虫交配日龄对寿命不存在显著的交互作用。进一步求得雌成虫寿命(y)与雄虫日龄(x1)和雌虫日龄(x2)的回归方程为y=35.85+0.23x1+0.40x2,R2值为0.9069。总之,随雌、雄成虫交配日龄的增加,雌虫的寿命随之延长,其中,雌虫交配日龄的延迟,对其寿命的影响更明显(表2,图3)。

图3 交配日龄对橘小实蝇雌成虫寿命的影响Fig.3 Effect of female and male age on adult longevity of female in Bactrocera dorsalis
3 结论与讨论
本研究结果表明,随雌虫交配日龄的增加,橘小实蝇交配率先增后减,其中,35日龄雌虫与35日龄雄虫组合的交配率最高,35日龄左右可能是橘小实蝇雌虫的中龄,也是最佳交配日龄。这可能是因为雄虫更偏爱与中龄雌虫交配(Bonduriansky, 2001)或者中龄雌虫更易接受雄虫的求偶所致。例如,马尾松毛虫Dendrolimuspunctatus中龄雌虫发生交配所需的时间最短,交配持续时间最长(Liuetal., 2014)。56日龄雌虫与35日龄雄虫的交配率则显著下降,这可能是因为随雌虫年龄的增加,其减少了相关的求偶行为(Torresetal., 2002)或是部分雌虫失去交配能力(冯波等, 2019)所致。
橘小实蝇的产卵量受到雌、雄交配日龄的影响。其中,随雌虫交配日龄的延迟,其产卵量减少,与烟草甲Lasiodermaserricorne(Barbaraetal., 2019)、斜纹夜蛾(Wuetal., 2018)、朱红毛斑蛾(Zhengetal., 2020)等昆虫种类结果相一致。可能是因为延迟交配导致未交配雌虫卵母细胞的发育受阻或减弱,雌虫为了维持寿命吸收部分来自卵的营养物质,致使产卵量下降(Torresetal., 2002)。许多研究表明,雄虫交配日龄的延迟会对产卵量造成负面影响(Keietal., 2014; Yangetal., 2017),而在橘小实蝇中,随雄虫交配日龄的增加,其产卵量会显著增加,认为可能是因为交配日龄的延迟,导致雄虫对其期望达到的交配次数下降(Elzingaetal., 2011),因此,为了获得更大的繁殖利益,在交配时进行了更多的射精投入所致(Bretmanetal., 2011; Esfandietal., 2019; Liuetal., 2020)。橘小实蝇卵的孵化率仅受雄虫交配日龄的影响,雌虫与年轻雄性交配后的卵孵化率更高,可能与年轻雄性高质量的精子有关,同样的现象也出现在斑禾草螟Chilopartellus(Dhillonetal., 2019)和茶尺蠖Ectropisobliqua(Yangetal., 2017)等昆虫中。
本研究发现,随雌、雄成虫交配日龄的延迟,橘小实蝇雌虫寿命随之增加,可能是由于延迟交配降低了雌虫与繁殖相关的能量消耗或者吸收了部分来自卵的营养物质所致(Subramanyam, 2003)。在不同昆虫种类中,延迟交配对雌虫寿命的影响不同;在一些昆虫种类中,延迟交配会延长雌虫寿命(李定旭等, 2009; 赵信等, 2012; 陆鹏飞等, 2013),而在另一类昆虫中,延迟交配会缩短雌虫寿命(Wangetal., 2010; 张清泉等, 2012)或不影响雌虫寿命(Zhengetal., 2020)。
总之,本研究发现延迟交配可在不同程度上降低橘小实蝇的交配率、产卵量和孵化率。这一结果对于橘小实蝇的防治工作具有一定的指导意义,但在田间延迟交配是否能导致其种群明显下降仍需要进一步研究。
猜你喜欢
湖北植保(2022年5期)2022-10-20
中国蚕业(2022年1期)2022-03-09
今日农业(2021年6期)2021-11-27
植物保护(2021年4期)2021-11-12
福建林业科技(2020年2期)2020-06-23
农家科技中旬版(2020年5期)2020-06-15
新农村(2018年32期)2018-04-02
现代畜牧科技(2015年12期)2015-10-21
江苏农业科学(2014年3期)2014-07-16
