
草地贪夜蛾生殖行为及其昼夜节律研究
2022-07-14 03:40张罗燕万小双
环境昆虫学报 2022年3期
张罗燕,汪 分,万小双,徐 进,2*,叶 辉
(1. 西南林业大学云南生物多样性研究院,昆明 650224;2. 西南林业大学西南山地森林资源保育与利用教育部重点实验室,昆明 650224;3. 云南大学生态与环境学院,昆明 650091)
草地贪夜蛾Spodopterafrugiperda(Smith),也称秋粘虫,隶属于鳞翅目Lepidoptera夜蛾科Noctuidae灰翅夜蛾属Spodoptera,原产于美洲热带和亚热带地区并在美洲大陆广泛分布,是玉米ZeamaysL.、水稻OryzasativaL.等重要粮食作物上的重大迁飞性害虫(Earlyetal., 2018; Kebede and Shimalis, 2018)。虽然草地贪夜蛾是一种长距离迁飞害虫,但在2015年之前并没有关于它在美洲以外分布的报道。自2016年开始,该害虫入侵了非洲、亚洲、大洋洲等60多个国家和地区(齐国君等, 2019)。截至目前,草地贪夜蛾已在世界113个国家和地区发生为害(齐国君等, 2020)。该害虫于2019年1月11日首次在我国云南省江城县发现,之后便开始以惊人的速度在我国蔓延(孙晓玲等, 2020)。截至2019年底,已扩散至我国 26省(自治区、直辖市)1 538个县(区),其中包括长江流域、黄河流域和东北玉米、小麦TriticumaestivumL.等重要粮食作物主产区(张智等, 2020)。在非洲,草地贪夜蛾可造成玉米年产量减少21%~53%,经济损失估计为24.81~61.87亿美元(Abrahamsetal., 2017)。在中国,草地贪夜蛾每年对玉米造成的经济损失估计为172.86~521.43亿美元(秦誉嘉等, 2020),对小麦造成的损失估计为155.71~901.43亿美元(徐艳玲等, 2020)。除了长距离迁飞能力,草地贪夜蛾还具有较强的繁殖力和抗药性(APRD, 2021),探索其绿色可持续防治方法刻不容缓。
昆虫生殖行为是性选择和进化理论的重要基础(Anholtetal., 2020),在害虫防治方面也有重要价值(徐汉虹等, 2020)。生殖行为主要包括求偶、交配和产卵等与繁殖直接或间接相关的行为过程,这些行为的发生及其特征、节律以及内外在因素是生殖行为研究的基本内容。当前,由于害虫抗药性不断增强和化学农药大量使用,造成了日趋严峻的经济损失和环境安全问题,基于昆虫信息素及生殖行为特征等建立起来的生物防治手段,例如性诱剂诱捕、交配干扰和雄性不育等,已经在一些重要农业害虫的监测和防治上成功应用并显示了巨大的潜力(徐汉虹等, 2020)。这些技术的成功运用有赖于对目标害虫的生殖行为充分的认识(Michereffetal., 2004)。草地贪夜蛾已对41种杀虫剂产生了不同程度的抗性,且其抗性仍在快速进化中(APRD, 2021)。开展草地贪夜蛾生殖行为等相关基础研究,有重要的科学意义及迫切的现实需求。
由于草地贪夜蛾的快速入侵和大面积危害,近年来对草地贪夜蛾的研究迅速增多。这些研究主要集中在其基础生物学、迁移和危害情况、防治技术的探索和建立等方面(Feldmannetal., 2019; Xiao, 2021),对于草地贪夜蛾生殖行为方面的研究报道仍然较少。其迁飞性、夜行性及生殖周期较长(10 d左右)等因素,都为草地贪夜蛾生殖行为的观测带来困难。因此,到目前为止,对于草地贪夜蛾生殖行为及节律,仍然缺乏较为全面的认识,亟需从性选择和物种进化角度进一步对其进行深入研究。基于此,本研究将理论研究和现实需求有机结合,在实验室条件下通过永久配对及每日更换配偶等处理,对草地贪夜蛾一生的生殖行为(包括雌虫召唤、雄虫求偶、雌雄交配及雌虫产卵等)和节律进行了系统观察和细致分析,并在此基础上对其性选择策略和生殖-生存权衡等进化问题进行了深入探讨,其研究结果也为草地贪夜蛾的综合治理提供了科学依据。
1 材料与方法
1.1 供试虫源
本研究所用草地贪夜蛾,其亲代于2019年7月采自云南省曲靖市沾益区田间受害玉米植株上。亲代幼虫采回后在实验室采用人工饲料进行饲养和扩繁,然后用其子代进行试验。本研究于2020年7月至9月进行,研究前该虫在实验室里繁殖约9代。人工饲料配方及配制方法同李广宏等(1998),幼虫饲养于透明塑料盒中(长23 cm×宽15 cm×高9 cm),饲养温度设为26℃±1℃,相对湿度为60%~80%,光周期为L ∶D=14 ∶10。待幼虫老熟化蛹后,将蛹放置于同样规格的装有细沙的塑料盒中直至羽化,取性成熟雌雄成虫进行配对传代以获得足够的试验用虫。
1.2 草地贪夜蛾生殖行为及其昼夜节律观察
本试验用成虫采集过程中需确保其未交配过并记录其日龄,具体方法如下:待幼虫老熟化蛹后,根据蛹腹部特征(董前进等, 2019)将雌雄蛹分开并分别放入不同的塑料盒中进行羽化。羽化多发生在灯灭前1 h和灯灭后1 h这两个小时之内,为减少试验误差,只取这两个小时之内羽化的成虫立即配对进行下述生殖行为观察。将羽化后的1个暗期和1个光期记为0日龄,再之后的1个暗期和1个光期记为1日龄,以此类推。
在上述环境条件下通过永久配对(Permanent pairing, PM)对及每日更换配偶(Change mate daily, CM)两种处理,对草地贪夜蛾成虫一生的生殖行为(包括雌虫召唤、雄虫求偶、雌雄交配及雌虫产卵)及其节律进行了观察和比较分析。PM为一雌一雄在羽化后立即放入上述透明塑料盒进行配对,然后每日进行生殖行为观察直至死亡(每对为1个重复,共20个重复,n=20)。CM为新羽化一雌一雄成虫放入塑料盒中配对后关1 d,之后交换雄性配偶,再关1 d之后又交换雄性配偶,依此每日交换1次配偶直致雌虫死亡,同一雌雄一生中只相遇1次,每日进行生殖行为观察直至死亡(每对为1个重复,共20个重复,n=20)。配偶交换处理均在天黑前20 min之内完成。永久配对也在天黑前20 min之内进行相似处理,只是将雄虫取出后再放入原雌虫盒内,并不交换配偶。在塑料盒中铺上白纸作为产卵场所,盒中放入10%蔗糖水以供成虫取食,产卵纸和糖水每日更换。
成虫的行为观察每10 min进行一次(Wangetal., 2021),每次快速扫描所有配对的成虫并记录以下行为:雄虫求偶-雄虫在雌虫周围及头顶跳跃及扇翅或伸出其外生殖器试图与雌虫外生殖器结合;雌虫召唤-雌虫将它的腹部从两翅间向上穿过且高高翘起并露出产卵器及性信息素腺;交配-雌与雄的外生殖器结合在一起。记录观察到的每头虫的每一种行为的发生时间。每天在光周期(白天)下收集当天所产的卵并置于透明塑料培养皿(8.5 cm直径×1.5 cm高)中进行孵育,之后在解剖镜下对每日产卵数和孵化数(幼虫的孵化大概在产卵3 d后)进行统计。在暗周期(夜间)下用30 W红灯照明进行行为学观测。
1.3 数据分析
1.3.1数据整理
计算雌虫召唤、雄虫求偶和雌雄交配的实时及累计百分率,每日交配百分率,交配累计百分率,每日及一生交配次数、产卵量、孵化数和孵化率,前5 d产卵速率等,主要统计方法如下:
雌虫召唤实时百分率(%)=每小时里发生了召唤行为的雌虫数/处理重复数×100
雄虫求偶实时百分率(%)=每小时里发生了求偶行为的雄虫数/处理重复数×100
交配实时百分率(%)=每小时里发生了交配行为的雌虫数/处理重复数×100
雌虫召唤累计百分率=对雌虫召唤实时百分率按小时进行累加,每日分别计算
雄虫求偶累计百分率=对雄虫求偶实时百分率按小时进行累加,每日分别计算
交配累计百分率=对交配实时百分率按小时进行累加,每日分别计算
每日交配雌虫按次分类百分率(%)=每日第几交配的雌虫数/处理重复数×100
按一生交配次数分类雌虫百分率(%)=一生交配了某一次数的雌虫数/处理重复数×100
前5 d产卵速率=前5 d产卵数/总卵数
1.3.2数据分析
对数据进行正态分布检验及方差齐性分析,然后对满足正态分布及方差齐性的处理进行单因素方差分析(ANOVA),并用LSD进行多重比较。其中关于交配次数和交配持续时间的数据经过ln(X)变换后满足正态分布。经尝试不同数据变换后仍不满足正态分布及方差齐性要求的数据则使用Kruskal-Wallis进行非参数检验。
采用线性回归分析探究了草地贪夜蛾雌蛾生殖行为(召唤时长、交配次数、总产卵数和前5 d产卵速率)与其寿命之间可能存在的相关性,以及草地贪夜蛾雄蛾生殖行为(求偶次数和交配次数)与其寿命之间是否存在相关性。
所有数据分析均使用SPSS Statistics 25.0统计软件进行。拒绝水平设为α<0.05。如无特殊说明,本研究的数值为算术平均值±标准误(means±SE)。
2 结果与分析
2.1 雌虫召唤行为节律与雄虫求偶行为节律
本研究采用永久配对(Permanent pairing, PM)和每日更换配偶(Change mate daily, CM)两种处理,对草地贪夜蛾成虫羽化后10 d(羽化当天开始直至9日龄)的生殖行为节律进行了观察和分析,结果表明雌虫召唤行为存在明显的昼夜节律,即均发生在夜间,并受到日龄和配偶的影响(图1, 图2)。从日龄上看,雌虫在羽化当日(0日龄)即可出现召唤行为,但发生频率和比率较低;1日龄召唤行为频率和比率显著提升,所有雌虫在这一日均出现了召唤行为;2~5日龄期间均保持了较高的召唤行为频率和比率,每日累计召唤发生比率在两种处理下均在80%以上;6~9日龄间召唤行为逐渐减少。从召唤行为在夜间发生的时间上来看,0日龄时主要发生在后半夜,1~5日龄多发生在前半夜,之后的6~9日龄间在夜间各时段发生频率相似。从总体上看,两种配对处理下雌虫召唤行为日节律相似,但是在CM下雌虫后期(6~9日龄)召唤行为与PM相比下降较慢。
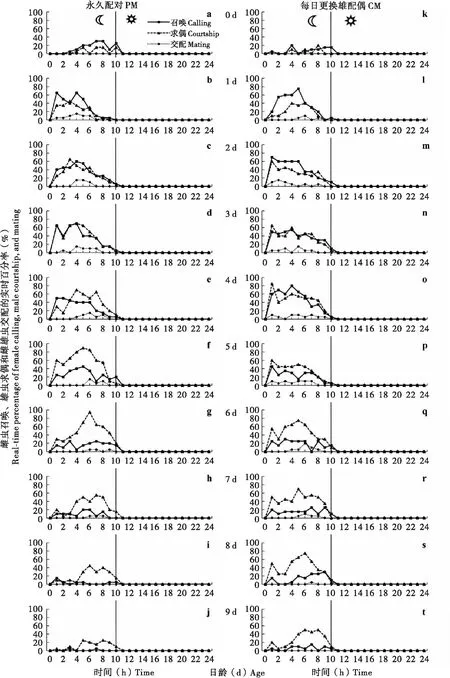
图1 草地贪夜蛾雌虫召唤、雄虫求偶和雌雄虫交配的实时百分率Fig.1 Real-time percentage of female calling, male courtship, and mating of Sopdoptera frugiperda
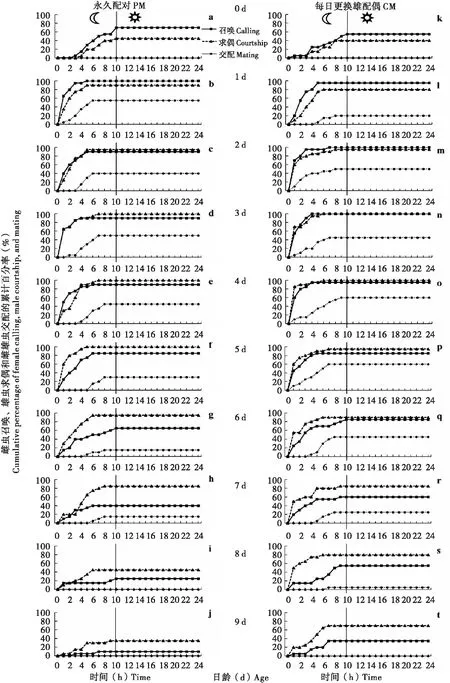
图2 草地贪夜蛾雌虫召唤、雄虫求偶和雌雄虫交配的累计百分率Fig.2 Cumulative percentage of female calling, male courtship, and mating of Sopdoptera frugiperda
同样,雄虫的求偶行为也出现了明显的昼夜节律,并因日龄和配偶的不同而不同(图1, 图2)。雄虫在羽化当日(0日龄)也出现了较低频率和比率的求偶行为;求偶行为在1日龄出现了显著提升,大部分(>80%)雄虫在这一日出现了求偶行为;从2~7日龄期间雄虫均保持了较高的求偶行为频率和比率,每日累计求偶发生率均在80%以上;在8~9日龄,PM雄虫求偶行为显著下降,CM雄虫求偶行为也有所下降,但下降幅度相对较低。从求偶行为在夜间发生的时间点上来看,两种处理间也存在明显差异,其差异主要表现在CM雄虫求偶高峰多发生在天刚黑之后,也即刚与新配偶配对之后不久。
求偶与召唤行为节律间存在较高的相似性和同步性,但也存在一定差异(图1, 图2)。从实时行为发生比率来看(图1),在0~1日龄,召唤行为明显高于求偶行为;在2~4日龄,召唤行为与求偶行为相差不大;而从5日龄开始直到9日龄为止,求偶行为明显高于召唤行为。从累计行为发生比率来看(图2),在0~1日龄,召唤行为高于求偶行为;而从2日龄开始直到9日龄,求偶行为多数情况下高于召唤行为。
雌虫召唤行为多为一个持续的过程(最高可持续数小时),而雄虫求偶行为多为偶发过程,且持续时间较短,几秒到几分钟不等。基于这些特性,对召唤时长和求偶次数进行了统计。结果表明,在两种处理下,各日龄的召唤时长均随日龄的增长呈现下降趋势(PM:F9,190=18.431,P<0.0001; CM:F9,190=14.728,P<0.0001; 图3-a),PM和CM之间只在最后2个日龄(8日龄和9日龄)存在显著差异(P<0.05)。CM雌虫一生召唤时长高于PM雌虫,但差异不显著(F1,38=1.735,P=0.1956; 图3-b)。各日龄的求偶次数均随日龄的增长呈现先上升再下降的趋势(PM:F9,190=13.625,P<0.0001; CM:F9,190=7.306,P<0.0001; 图3-c),处理之间在5日龄、8日龄和9日龄(P<0.05)存在显著差异。CM雄虫一生求偶次数高于PM雄虫,但差异不显著(F1,38=1.416,P=0.2414; 图3-d)。
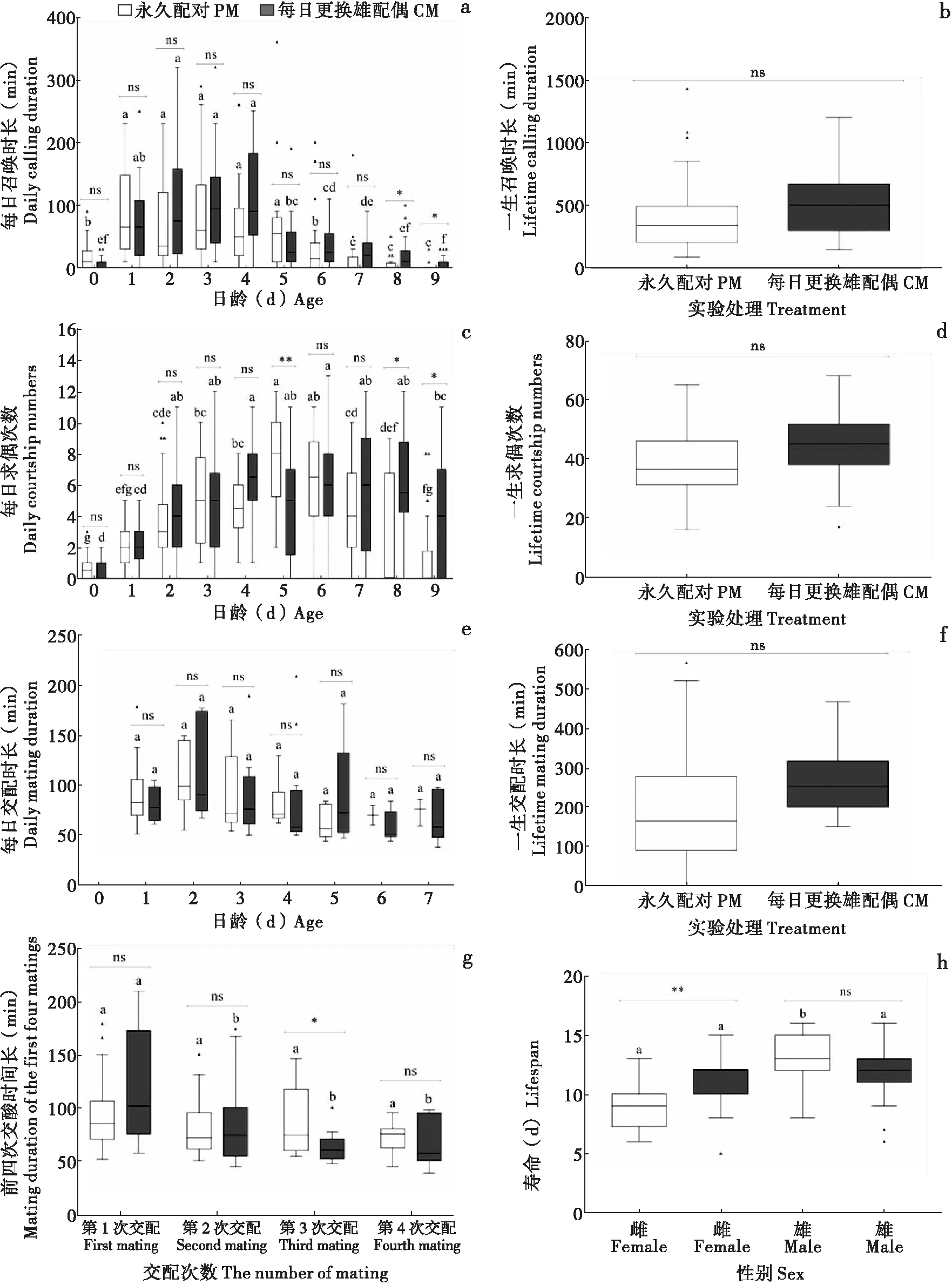
图3 草地贪夜蛾召唤时长、求偶次数、交配时间及寿命Fig.3 Calling duration, number of courtships, mating duration and lifespan of Spodoptera frugiperda注:a,每日召唤时长;b,一生召唤时长;c,每日求偶次数;d,一生求偶次数;e,每日交配时长;f,一生交配时长;g,前4次交配时长;h,雌雄寿命。Note: a, Daily calling duration; b, Lifetime calling duration; c, Daily courtship numbers; d, Lifetime courtship numbers; e, Daily mating duration; f, Lifetime mating duration; g, Mating duration of the first four matings; h, Male and female lifespan.
2.2 交配行为及其节律
草地贪夜蛾雌雄虫均可在羽化当日出现召唤或求偶行为,但在这1日未见交配现象(图1, 图2)。交配行为最早发生于1日龄成虫中。交配均发生于夜间,从交配发生的时间点来看,交配高峰多出现在召唤和求偶高峰之后。交配的发生多以雄虫主动为主,少数情况下几次求偶和尝试即可成功开始交配,多数情况下需要多次求偶和尝试才能成功。交配过程中,雌雄蛾多呈“一”字形(图4-a),少数呈“八”字形(图4-b),雌雄虫均静止不动。交配开始至10 min之内,人为地移动交尾过程中的成虫较易打断其交配;交配开始10 min之后,人为移动不易打断其交尾。交尾完成后,雌雄成虫分开,但多静止不动,活动性较低。

图4 草地贪夜蛾交配、产卵及幼虫Fig.4 Mating, egg laying and larvae of Spodoptera frugiperda注:a,雌雄蛾呈“一”字形交配过程;b,雌雄蛾呈“八”字形交配过程;c~h,初生卵,呈白色、灰色和淡紫色等;i,即将孵化的卵,棕黑色;j, k,初孵幼虫。Note:a, “-” liked mating posture; b, “v” liked mating posture; c~h, Neonatal eggs with white, gray or lavender color; i, Mature eggs, brown-black; j, k, Neonatal larvae.
交配行为节律同样与日龄和配偶密切相关。从每日交配雌虫按次分类百分率来看(图5-a, b),PM雌虫不论是第1次还是后面几次交配多表现出先高后低的趋势,而CM雌虫多表现为先低后高再降低的趋势;类似趋势也表现在每日平均交配次数(图5-c)和每日交配雌虫百分率(图5-e)上。从每日交配累计次数(图5-d)来看,1~4日龄期间PM高于CM,而5~9日龄间CM高于PM。从每日交配雌虫累计百分率(图5-f)来看,1~3日龄间PM高于CM,4~9日龄间CM高于PM。CM雌虫到5日龄时均发生了至少1次交配,即交配雌虫比率在这1日已达100%(图5-f);而PM雌虫直至7日龄才达到最高交配比率95%,之后至死依然有5%的雌虫未进行交配(图5-f)。
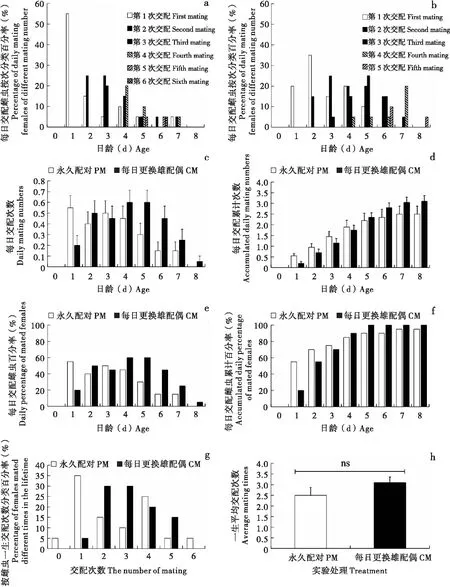
图5 草地贪夜蛾交配次数及日节律Fig.5 Mating frequency and daily rhythm of Spodoptera frugiperda注:a,永久配对的每日交配雌虫按次分类百分率;b,每日更换雄配偶的每日交配雌虫按次分类百分率;c,每日交配次数;d,每日交配累计次数;e,每日交配雌虫百分率;f,每日交配雌虫累计百分率;g,按雌虫一生交配次数分类百分率;h,一生平均交配次数。Note: a, Percentage of daily mating females of different mating number from PM; b, Percentage of daily mating females of different mating number from CM; c, Daily mating numbers; d, Accumulated daily mating numbers; e, Daily percentage of mated females; f, Accumulated daily percentage of mated females; g, Percentage of females mated different times in the lifetime; h, Average mating times.
从一生交配次数来看,处理间也出现了明显差异(图5-g)。PM雌虫有5%终生未发生交配,一生中只交配了1次的雌虫占比最高(35%),其余雌虫发生了2次及以上交配,单头雌虫最高交配了6次。CM雌虫一生中只交配了1次的只占5%,其余的雌虫发生了2次及以上交配,一生只交配了2次或3次的雌虫占比最高(均为30%),单头雌虫最高交配了5次。CM雌虫一生平均交配次数为3.1±0.261次,高于PM雌虫的2.5±0.373次(图5-h)。
从每次交配的持续时间来看,各日龄的每次交配时长均无显著差异(PM:χ2=10.511,P=0.1047;CM:χ2=11.396,P=0.0769;图3-e)。CM雌虫一生交配时长高于PM雌虫,但差异不显著(F1,38=1.206,P=0.2791;图3-f)。在PM中,雌虫第1次、第2次、第3次和第4次交配的持续时间之间没有显著差异(F3,43=1.1,P=0.3595;图3-g);而在CM中,第1次交配的持续时间明显高于第2次、第3次和第4次交配的持续时间(F3,55=6.374,P=0.0009;图3-g)。雌虫第1次、第2次和第4次交配的持续时间在处理间没有显著差异(P>0.05),仅在第3次交配上有显著差异(P<0.05;图3-g)。
2.3 产卵行为及节律
草地贪夜蛾雌虫在交配结束后当夜未观察到产卵现象,多数雌虫在交配后下一夜开始产卵。卵呈圆顶型,直径0.4 mm,高为0.3 mm,通常100~200粒卵堆积成块状,卵块上多覆盖有黄色鳞毛,初产时为浅绿、白色、灰色和淡紫色等(图4-c, h),孵化前渐变为棕黑色(图4-i),之后幼虫开始孵化(图4-j, k)。雌虫日产卵量存在明显的单峰形产卵节律(图6-a, b)。PM雌虫日产卵高峰发生在5日龄,当日产卵量为209±44粒(图6-a);CM雌虫日产卵量高峰发生于6日龄,当日产卵量为279±45粒(图6-b)。PM雌虫一生的产卵数和孵化数分别为1 079±121粒和1 033±127粒(图6-c);CM雌虫一生的产卵数和孵化数为1 214±106粒和1 128±126粒(图6-d),均高于PM,但差异不显著(产卵数:χ2=1.026,P=0.3112; 孵化数:χ2=0.360,P=0.5483)。从日孵化率来看,PM在最后2 d较低,CM在最后 1 d 较低(图6-e);PM总孵化率稍高于CM,但差异不显著(F1,38=0.29,P=0.5934,图6-f)。
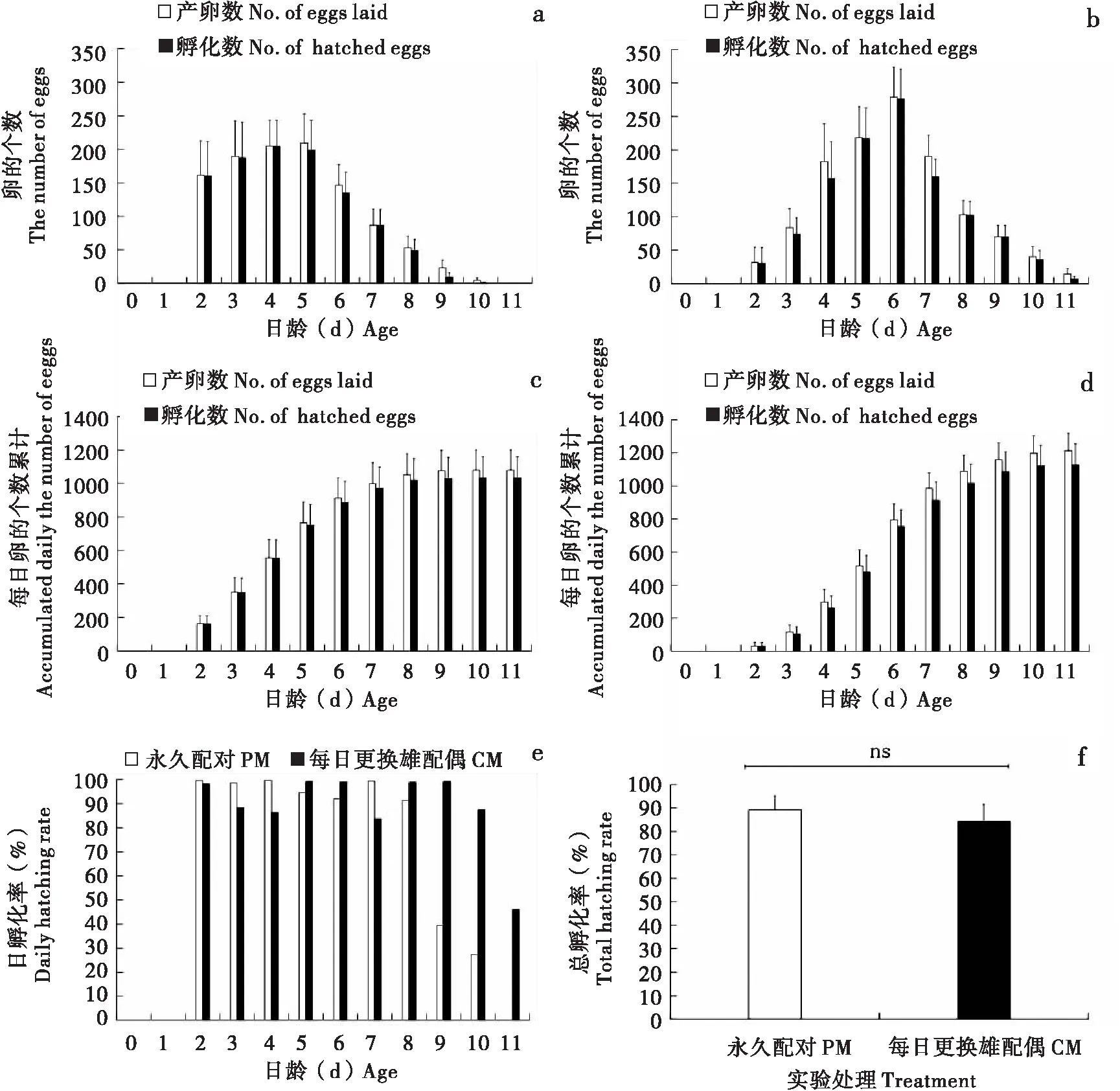
图6 草地贪夜蛾产卵量、孵化数及孵化率Fig.6 Number of eggs laid, number of hatched eggs and hatching rate of Spodoptera frugiperda注:a,永久配对的日产卵量及孵化数;b,每日更换雄配偶的日产卵量及孵化数;c,永久配对的累计产卵量和累计孵化数;d,每日更换雄配偶的累计产卵量和累计孵化数;e,永久配对和每日更换雄配偶的日孵化率;f,永久配对和每日更换雄配偶的总孵化率。Note: a, Daily eggs laid and hatched eggs of PM; b, Daily eggs laid and hatched eggs of CM; c, Accumulated daily eggs laid and hatched eggs of PM; d, Accumulated daily eggs laid and hatched eggs of CM; e, Daily hatching rate; f, Total hatching rate.
2.4 生殖行为与成虫寿命关系
雄虫平均寿命高于雌性,雌雄寿命差异在PM中显著(F1,38=37.027,P<0.0001, 图3-h),而在CM中不显著(F1,38=0.56,P=0.44, 图3-h)。CM雌蛾寿命显著高于PM雌蛾(F1,38=10.851,P=0.0027, 图3-h),而雄蛾寿命在两种处理间没有显著差异(F1,38=3.51,P=0.0518, 图3-h)。
为了进一步解析生殖与生存间可能存在的权衡关系,对成虫一生的生殖行为与寿命之间可能存在的相关性进行了线性回归分析。对于雌虫,前5 d产卵速率与寿命间在两种处理条件下均存在显著负相关性(图7);然而交配次数或召唤时长与寿命之间在两种处理下均不存在相关性(P>0.05)。对于雄虫,交配次数或求偶次数与寿命之间相关性在两种处理条件下均不显著(P>0.05)。
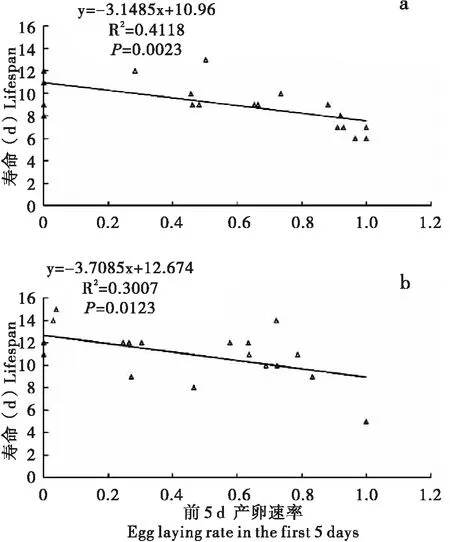
图7 草地贪夜蛾前5 d产卵速率与雌虫寿命的线性关系Fig.7 Linear relationship between the egg laying rate in the first 5 days and female lifespan of Spodoptera frugiperda注:a,永久配对处理;b,每日更换雄配偶处理。Note: a, Relationship in PM; b, Relationship in CM.
3 结论与讨论
许多鳞翅目昆虫的生殖行为和其他活动并不全天出现,其中产卵、交配、进食或飞行发生在白天或夜间的特定时期,存在明显的昼夜节律性(Lazzari and Insausti, 2008)。对于夜蛾科昆虫,生殖活动通常发生在夜间(Cardé, 1973)。本研究也显示,草地贪夜蛾的生殖行为活动均发生在夜间。昼夜节律是昆虫界最普遍的生物节律,是一种最常见的生物钟现象,而光周期是影响昆虫昼夜节律最重要的因素之一。光周期的存在及其引起的转录、翻译负反馈环路中出现的转录激活物和抑制物是昆虫生物钟形成和维持的基础(Giebultowicz, 1999)。光周期作为影响昆虫生活节律的重要因子,直接影响昆虫的发育、生理代谢以及种群季节性变动等(陈广平等, 2009)。除了受到光周期和生物钟的主调节,昆虫行为节律也受到外界环境的影响,如气温、降雨等环境变化可能影响昆虫的生殖行为(吴少会等, 2006)。晚上温度比白天低,湿度比白天高,更容易躲避天敌的捕食,这些条件可能更有利于夜蛾的羽化、展翅和寻找配偶(Pompanonetal., 1995)。
本研究在实验室模拟自然条件下可能出现的择偶现像,采用永久配对(Permanent pairing, PM)和每日更换配偶(Change mate daily, CM)两种处理,对草地贪夜蛾成虫一生(约10 d)的生殖行为和节律进行了连续观察和分析,结果表明,雄虫求偶行为节律与雌虫召唤行为节律存在明显相似性和同步性,同时交配高峰多出现在召唤和求偶行为高峰之后(图1, 图2),表明召唤和求偶行为是交配发生的重要因素。前人研究表明,雌蛾召唤(释放性信息素)等生殖活动与雄蛾飞行活动以及对雌蛾的应答能力相吻合(Prestwich and Blomquist, 1987)。雄性和雌性之间的这种性行为的协调性一方面确保了觅偶的高成功率,并有效地提高了种间的生殖隔离(Grootetal., 2015)。这些行为的发生和持续时间也受到昆虫年龄(Wangetal., 2015)、交配状况(Yangetal., 2011)以及外源(如光周期和温度)(Lietal., 2015)和内源(如荷尔蒙)(Blochetal., 2013)性因子的影响。对蛾类昆虫的这些行为节律研究,有利于有针对性的制定其精准防控策略,例如可根据目标害虫的生殖行为节律及生殖周期来确定最佳诱捕时期,或在雄性不育技术中确定不育雄性个体投放时间,以达到精准高效的防控目的(Vernonetal., 2014)。
雌虫通过召唤吸引雄虫,当雄虫找到雌虫后,雄虫一般需要进行或简单或复杂的求偶过程才能获得交配(Phelan and Baker, 1990)。蛾类交配过程中,雌虫的产卵器完全收回,雄虫外生殖器的抱器抱住雌虫的第7腹节,几丁质化的插入器插入雌虫交配囊的颈部,可以外翻的阴茎伸入到交配囊的深处,然后在那里形成精包(Norris, 1932)。本研究发现草地贪夜蛾在交配开始10 min之后,人为移动不易打断其交尾,其原因可能是精包这时已经开始形成,精包生成过程中无法在不造成损伤的情况下人为终止其交配(徐进和陈鹏, 2018)。交配完成后,精包中的精子需通过精子导管到达前庭再最终到达储精器(Friedlanderetal., 2005)。精子需在储精器中成熟后才可重新进入前庭使卵受精(Friedlanderetal., 2005)。鳞翅目昆虫精子一般需要数小时到十几小时不等才能从精包中迁移到储精器(Howell, 1991),这可能正是大部分鳞翅目昆虫(包括草地贪夜蛾;图6)一般在交配后第2天才开始产卵的主要原因。
研究表明,在许多昆虫中,由于雌虫具有较好的储精能力或专门的储精器官,一次交配足以使雌性获得足够多的精子用于使其所有卵子受精(Arnqvist and Nilsson, 2000)。尽管如此,绝大多数雌性昆虫不仅与不同的雄性而且与同一个雄性进行多次交配。昆虫的交配活动是一个非常耗时耗能的生理过程,此外,还存在身体伤害、病原菌传播和被捕食等风险(Arnqvist and Nilsson, 2000)。从进化意义上来讲,多次交配带来的好处应该高于交配可能造成的损失。性选择及进化理论研究表明多次交配有助于雌虫获得物质(营养)利益或基因(遗传)利益(胡阳等, 2010)。物质利益主要包括精子补充以及从精液中获得营养补充从而有助于提高生殖力和寿命(Arnqvist and Nilsson, 2000; Thonhauseretal., 2013)。基因利益主要包括多次交配有助于减少子代基因不相容性、提高子代基因质量或基因多样性等(Suzakietal., 2013)。为了最大限度地获取基因利益,雌性在再次交配过程中已经进化出了更喜欢新配偶的择偶策略(Zehetal., 1998)。到目前为止,已经在8种无脊椎动物中发现存在喜好新配偶的择偶现象(Bateman, 1998; Zehetal., 1998; Archer and Elgar, 1999; Eakley and Houde, 2004; Ivyetal., 2005; Xu and Wang, 2009; Tanetal., 2013; Mestreetal., 2015)。本研究结果表明,CM雌性与PM雌性相比表现出较高的生殖行为活力(图1~图3,图5),PM雌性早中期产卵量较高而CM雌性中期产卵量最高(图6)。这些结果表明,草地贪夜蛾雌性可以区分新的和以前的配偶,并表现出明显的喜新厌旧的趋势。雌虫在遇到新的配偶或与新配偶交配后,延缓产卵或下调产卵速率可能有助于新配偶的精子参与授精并提高精子竞争以获取基因利益的最大化(Archer and Elgar, 1999; Xu and Wang, 2009; Lietal., 2014; Mestreetal., 2015)。
本研究结果表明,前5 d产卵速率与雌虫寿命间存在显著负相关性(图7),这一结果暗示早期产卵量较高的雌虫寿命较短,反之亦然。前人研究表明雌虫能够通过交配从雄虫获得营养物质,如精液、附腺物质等,从而可以改善和补充生殖消耗,有助于提高雌虫生殖力和寿命(Gillott, 2003)。另一方面,精液中来自雄性附腺分泌的活性物质可以促进卵的发育和成熟,这些因子在促进雌性生殖力提升的同时可能对其生存带来负面影响(Xu and Wang, 2011; Yuetal., 2014)。生殖-生存权衡模型指出,基因在早期繁殖和后期生存之间具有拮抗作用,即早期生殖力较高的雌性寿命较短,反之亦然(Silbermann and Tatar, 2000; Okadaetal., 2014)。该模型已经在果蝇中基于热激蛋白表达获得了一些证据(Silbermann and Tatar, 2000; Okadaetal., 2014)。本研究表明草地贪夜蛾生殖与寿命之间的关系符合早期繁殖-后期存活基因拮抗假说,其相关生理和基因等内在调节机制值得进一步深入研究。
猜你喜欢
植物保护(2021年4期)2021-11-12
四川蚕业(2021年1期)2021-02-12
江苏科技报·E教中国(2020年10期)2020-12-14
江苏科技报·E教中国(2020年6期)2020-07-30
教学月刊·小学数学(2020年4期)2020-05-13
华夏地理(2019年2期)2019-07-24
小学教学参考(数学)(2017年1期)2017-03-01
理科考试研究·高中(2016年5期)2016-05-14
百科知识(2015年7期)2015-04-08
江苏农业科学(2014年3期)2014-07-16
