
湘西地区吕氏泰勒虫的单倍型多样性及种系发育关系分析
2022-07-06 05:52李中波侯强红
河南农业科学 2022年4期
李中波,侯强红,李 晖,舒 鸣
(1. 怀化职业技术学院,湖南 怀化 418000;2. 湘西民族职业技术学院,湖南 吉首 416000)
吕氏泰勒虫(Theileria luwenshun)为一种常见且重要的蜱传性血液原虫[1],隶属于泰勒科、泰勒属[2],寄生于山羊、绵羊的红细胞、淋巴细胞和巨噬细胞内[3],可引发一种以发热、贫血、黄疸、消瘦、体表淋巴结肿大及拉血红蛋白尿为特征性临床症状的血液原虫病——羊泰勒虫病[4‐5]。吕氏泰勒虫1914 年首次被发现于埃及的患病羊只[6],而后世界上多个国家和地区(如欧洲、非洲、中东及亚洲)陆续有报道[7],尤以热带、亚热带地区流行最为普遍[8]。吕氏泰勒虫因具有较强的感染力和致病力,且分布广泛,已成为我国范围寄主的优势虫株之一[9]。该虫一旦感染羊只,可引起被感染羊只出现发热、溶血性黄疸、贫血、消瘦及拉血红蛋白尿等症状[4‐5],如救治不及时或不当可导致羊只大量死亡[10],给养殖户造成巨大的经济损失。钟维等[11]调查四川阿坝州牦牛及藏绵羊中吕氏泰勒虫的感染情况,发现其感染率高达75%。此外,多种因素影响吕氏泰勒虫病的流行,如气候环境对吕氏泰勒虫的分布影响极大[12]。地理位置的阻隔,在一定程度上也会影响吕氏泰勒虫的基因交流、种群结构和系统发育。地理位置为吕氏泰勒虫的物种多样性、遗传多样性和单倍型多样性提供先决条件。
单倍体基因型简称单倍型,是指在一染色单体上具有统计学关联意义的一类单核苷酸多态性[13]。我国地域宽广,地理、气候多样,是造就物种多样性的基本条件。同一物种为快速适应栖息地的环境和气候,往往在自身的形态及遗传结构等方面发生变化,形成单倍型多态性及个体多样性。然而,一个物种的单倍型多样性及遗传多样性又是研究物种遗传资源保护不可或缺的内容。因此,研究一个物种的单倍型多态性和种系发育关系,不仅为物种遗传资源保护提供重要的信息和理论基础,而且还有助于今后理解该物种在自然环境中的进化过程,因而这方面的研究已越来越受到人们的重视和关注。迄今为止,国内外已开展诸多关于寄生虫的遗传多样性、遗传结构、单倍型多态性及种系发育关系等方面的研究。AMZATI 等[14]基于线粒体细胞色素c氧化酶亚基Ⅰ(cox1)基因和12S rRNA序列研究具尾扇头蜱(Rhipicephalus appendiculatus)的种群结构和单倍型;MALATJI 等[15]基于cox1基因分析鸡蛔虫(Ascaridia galli)的遗传结构和单倍型。我国仅有冯雪皎[16]基于线粒体cox1、16S rDNA 和核糖体ITS-2 基因对4 个地理种群长角血蜱(Haemaphysalis longicornis)的单倍型及遗传多样性进行研究。
核糖体、线粒体基因均具有进化速度快、缺乏基因重组及母系遗传等特征[17‐19]。目前,核糖体、线粒体基因已广泛应用于种群结构、分子标记、生物进化、种系发育、物种鉴定、单倍型及进化基因组学等方面研究[20‐21]。国内学者研究集中在某一寄生虫的鉴定、归类、形态描述、遗传进化关系及其所携病原微生物检测等方面,鲜有学者就某一寄生虫的单倍型多样性进行研究。湘西地区毗邻云贵高原,为多山陵地带,辖区内含有极为丰富的蜱种,然其内沟壑纵横,气候多变,交通极为不便,给蜱虫的生殖交流造成一定程度上的地理隔离,间接为吕氏泰勒虫个体及遗传多样性的形成提供了先决条件,但关于这一地区吕氏泰勒虫单倍型多样性及种系发育关系等方面的研究甚少。因此,对该地区羊源吕氏泰勒虫的单倍型多态性及种系发育关系进行研究,不仅有助于该地区医学、公共卫生事业及畜牧业的发展,还可为今后泰勒虫病的分子流行病学调查、诊断、防控及相关药物的研发奠定基础。为此,以16株来自湘西地区的羊源性吕氏泰勒虫为样品,采用PCR 扩增其18S rRNA、cox1基因序列,分析吕氏泰勒虫的单倍型多态性及种系发育关系,为泰勒虫的种类鉴定、分类提供依据,并为该地区泰勒虫病的诊断、控制及分子流行病学调查提供参考。
1 材料和方法
1.1 样品采集
含有泰勒虫的山羊血液由湘西民族职业技术学院寄生虫实验室提供,将血液分别编号为XT1—XT16,置于-20 ℃保存,备用。
1.2 主要试剂
动物血液、组织DNA 提取试剂盒购买于北京天根有限公司;Taq高保真酶(Mix)购自宝生物工程(大连)有限公司;蛋白酶K 和DL2000 DNA Marker均购自北京全式金生物技术有限公司;电泳缓冲液购自南京诺唯赞生物科技股份有限公司。
1.3 DNA提取
根据血液、组织DNA 提取试剂盒的使用说明书,分别提取16 株吕氏泰勒虫的基因组DNA。再将提取好的DNA样品置于-20 ℃保存、备用。
1.4 目的基因的扩增、测序
根据NCBI 所公布吕氏泰勒虫的18S rRNA、cox1基因序列,设计2 对特异性引物(敏感度为0.047 ng/µL),即18S rRNA-F:5´-AACCTGGTTGAT‐CCTGCCAGTAGTCAT-3´,18S rRNA-R:5´-GATCC‐TTCTGCAGGTTCACCTAC-3´和cox1-F:5´-TGACT‐GGACTGTTTGGTGG-3´,cox1-R:5´-TGAGAAACC‐AGCGAAGTGC-3´,分别对16 株羊源吕氏泰勒虫的18S rRNA、cox1基因序列进行PCR 扩增[22]。PCR 扩增体系(总体积均为50µL):Taq高保真酶(Mix)25µL,DNA 模板1 µL,上下游引物各1 µL,dH2O 22 µL。PCR 扩增程序:95 ℃预变性5 min;95 ℃变性30 s,58 ℃/56 ℃(18S rRNA/cox1)退火30 s,72 ℃延伸1 min,共35个循环;最后72 ℃均再延伸5 min。待PCR 反应完成后,从每个反应体系取5 µL PCR产物进行1.5%琼脂糖凝胶电泳检测,观察结果并拍照。将所有阳性PCR 产物送至生工生物工程(上海)股份有限公司进行双向测序,获得吕氏泰勒虫18S rRNA、cox1基因的部分序列。
1.5 序列比对、校准和对齐
将所获得的18S rRNA、cox1基因序列分别进行BLAST 分析比对,再运用软件Clustal X 分别对所获得的18S rRNA、cox1基因序列进行校准和对齐[23]。
1.6 变异位点与单倍型多样性分析
基于所获得的18S rRNA、cox1及18S rRNA+cox1基因序列,运用软件DNAStar[24]分析基因变异位点;运用软件DnaSP 5.0[25]、Network 4.6[26]及Arlequin version 3.0[27]分析单倍型多样性。
1.7 种系发育关系分析
基于18S rRNA+cox1基因序列,以所获吕氏泰勒虫及从NCBI 中下载的吕氏泰勒虫虫株Li2(JF719831、JQ518295)、Li6(JF719833、JQ518296),尤氏泰勒虫(Theileria uilenbergi)(JF719835、JQ518295),马泰勒虫(Theileria equi)(KY111761、MF510491)及小泰勒虫(Theileria parva)(L02366、AB499089)为内群,并以双芽巴贝斯虫(Babesia bigemina)(JQ723014、AB499085)为外群,运用软件Clustal X、PhyML 3.0[28]及MEGA 5.0[29],采用最大相似法(ML)和GTR+I+G 模型(bootstrap=100)对吕氏泰勒虫的种系发育关系进行分析,再运用软件FigTree v1.3.1[30]描绘所构建的系统发育树。
2 结果与分析
2.1 湘西地区吕氏泰勒虫18S rRNA基因的PCR扩增结果
湘西地区吕氏泰勒虫18S rRNA 基因的扩增结果见图1。从图1 可以看出,16 个PCR 产物均为阳性,每个泳道只含1 个条带,无明显杂带,且大小与预期一致,均约为400 bp。
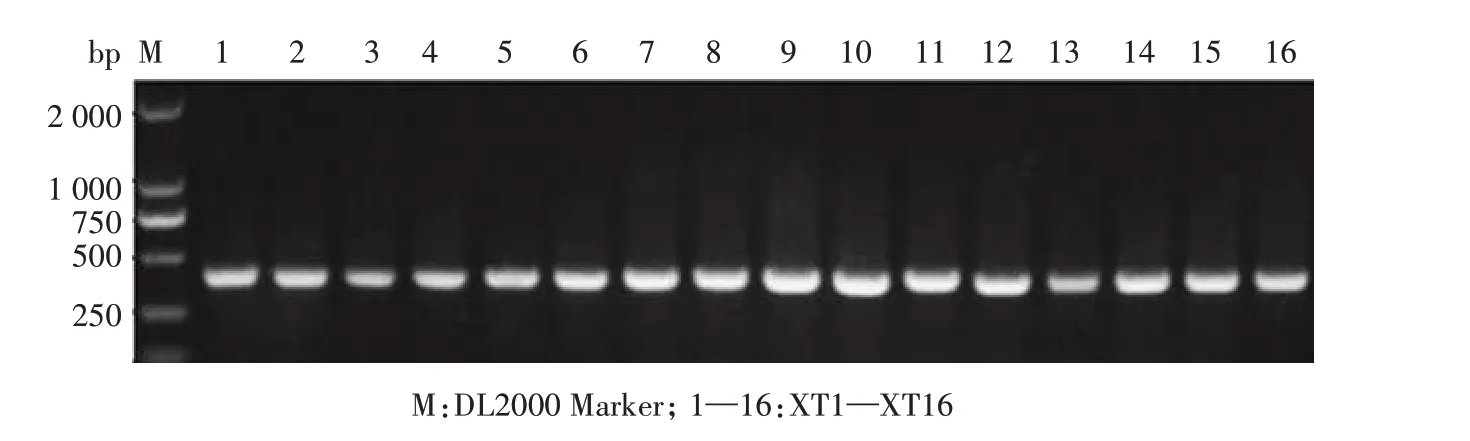
图1 湘西地区吕氏泰勒虫18S rRNA基因的PCR扩增结果Fig.1 PCR amplification of 18S rRNA gene of T.luwenshun from western Hunan
2.2 湘西地区吕氏泰勒虫cox1基因的PCR扩增结果
湘西地区吕氏泰勒虫cox1基因序列的扩增结果见图2。从图2 可以看出,16 个PCR 产物均为阳性,每个泳道只含1 个条带,无明显杂带,且大小与预期一致,均约为1 100 bp。
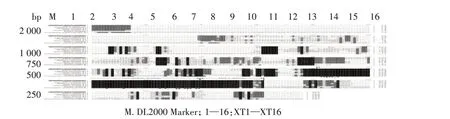
图2 湘西地区吕氏泰勒虫cox1基因的PCR扩增结果Fig.2 PCR amplification of cox1 gene of T.luwenshun from western Hunan
2.3 湘西地区吕氏泰勒虫18S rRNA、cox1基因序列的测序、校准、对齐与提交
测序结果显示,湘西地区吕氏泰勒虫18S rRNA、cox1基因序列长度分别处于387~398 bp 和1 096~1 102 bp。利用软件Clustal X 分别校准、对齐所有18S rRNA、cox1基因序列,最终得到18S rRNA基因序列的长度均为384 bp,cox1基因序列长度均为1 092 bp。将所有处理后的18S rRNA、cox1基因序列进行BLAST 比对分析,结果显示,湘西地区吕氏泰勒虫的18S rRNA、cox1基因序列与GenBank 中吕氏泰勒虫的相似度为98%~100%,从而确认所获序列均为目的基因序列。将湘西地区吕氏泰勒虫18S rRNA、cox1基因序列提交到NCBI,获得18S rRNA 基因登录号MW750496—MW750511,cox1基因登录号MW937264—MW937279。
2.4 湘西地区吕氏泰勒虫18S rRNA、cox1基因序列的变异位点分析
基于上述的18S rRNA、cox1基因序列,分别对湘西地区吕氏泰勒虫18S rRNA、cox1及18S rRNA+cox1基因序列的变异位点进行分析,结果见表1。由表1 可知,在吕氏泰勒虫18S rRNA(384 bp)基因序列中共含有9 个变异位点(转换7 个,颠换2 个);cox1(1 092 bp)基因序列中共存在2 个变异位点(转换2 个,颠换0 个);18S rRNA+cox1基因序列中,共包含11个变异位点(转换9个,颠换2个)。
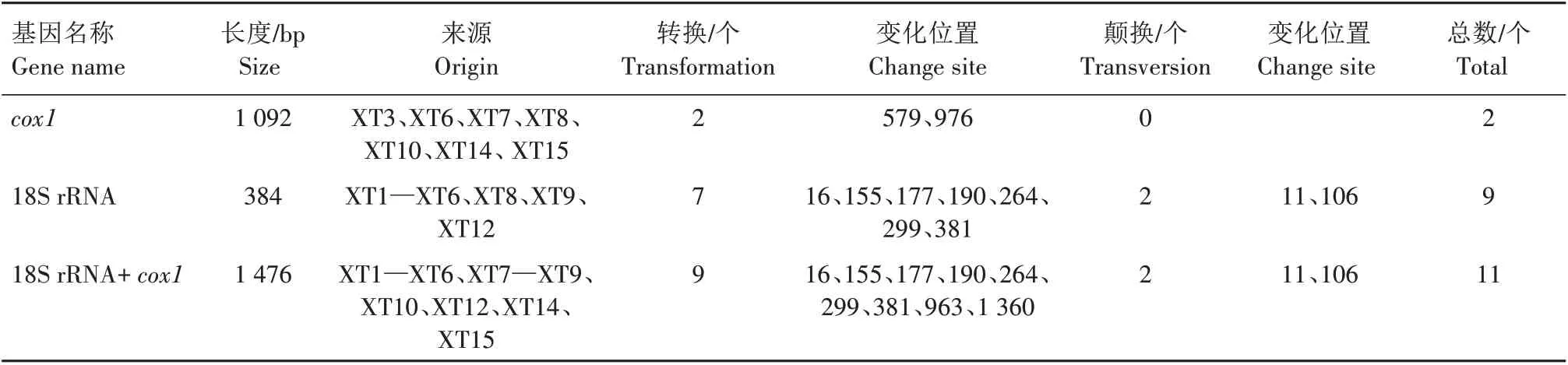
表1 湘西地区吕氏泰勒虫18S rRNA、cox1和18S rRNA+cox1变异位点Tab.1 The variable sites in 18S rRNA,cox1 and 18S rRNA+cox1 sequences of T.luwenshun from western Hunan
2.5 湘西地区吕氏泰勒虫单倍型多样性分析
运用软件DnaSP 5.0、Network 4.6 及Arlequin version 3.0 分别对湘西地区吕氏泰勒虫18S rRNA、cox1及18S rRNA+cox1基因序列的单倍型多样性进行分析(图3)。由图3 可知,湘西地区吕氏泰勒虫18S rRNA 基因序列含有7 个单倍型(h=7,A1—A7,Hd=0.775);cox1基因序列共包含4 个单倍型(h=4,B1—B4,Hd=0.625);18S rRNA+cox1基因序列共存在10个单倍型(h=10,C1—C10,Hd=0.925)。在A单倍型中,单倍型A1 共包含7 个序列,即XT7、XT10、XT11、XT13、XT14、XT15、XT16;单倍型A4包含4个序列,即XT1、XT2、XT3和XT4;而剩下的单倍型,每个单倍型只包含1 个序列。在B 单倍型中,单倍型B1 所包含的序列最多,共8 个,即XT1、XT2、XT4、XT5、XT11、XT12、XT13 和XT16;B2 包含2 个序列,即XT8 和XT10;B3 包含3 个序列(XT3、XT14、XT15);B4 也包含3 个序列,即XT6、XT7 和XT9。在C 单倍型中,单倍型C1(XT1、XT2、XT4)、C8(XT10、XT14、XT15)和C9(XT11、XT13、XT16)均含3个序列,其余的单倍型仅含1个序列。此外,在A、B 及C 单倍型中,A1、A4、B1、B3、B4、C1、C8 与C9 均为古老单倍型,均分别包含吕氏泰勒虫18S rRNA及cox1基因的古老序列。
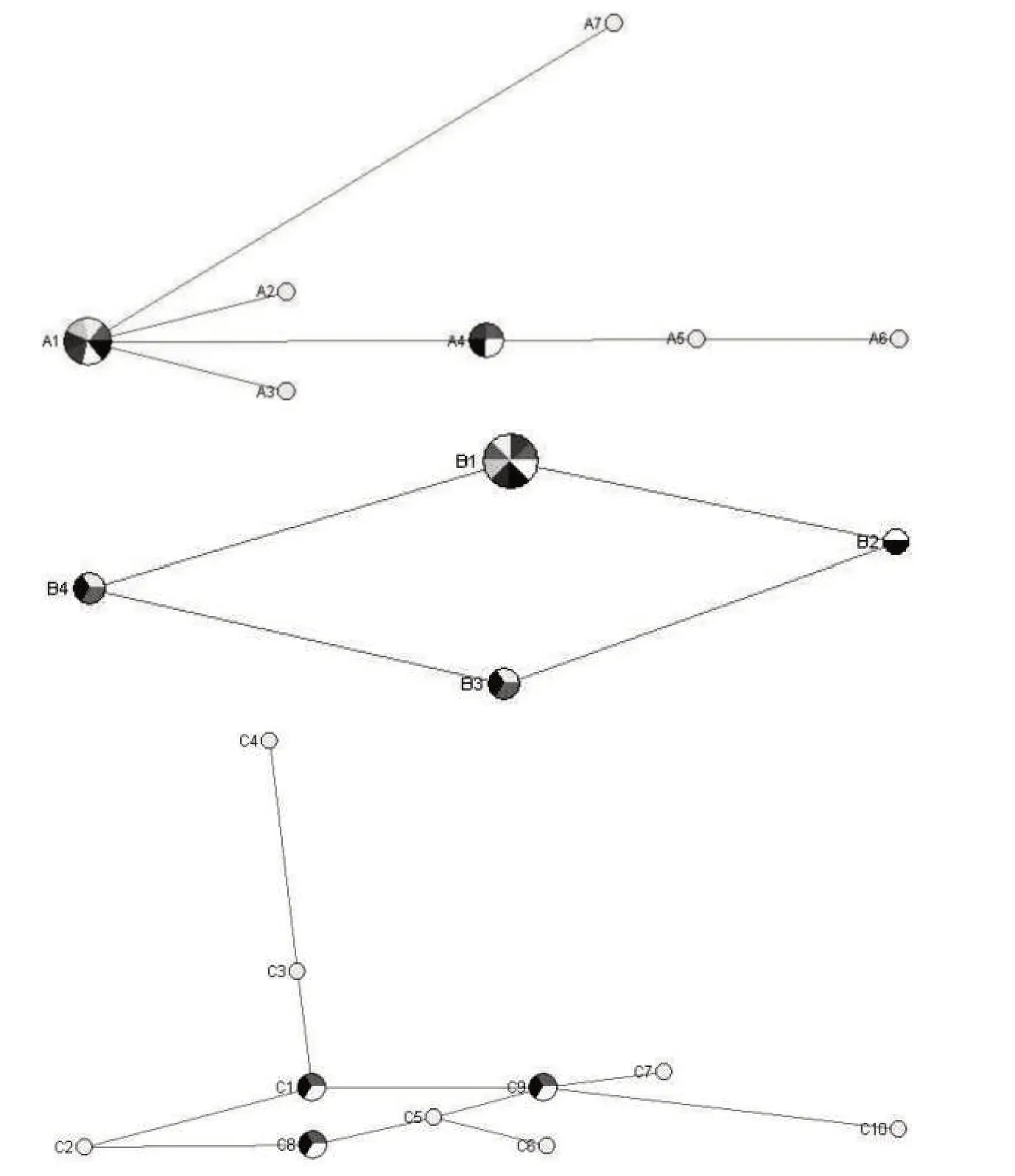
图3 湘西地区吕氏泰勒虫18S rRNA(A)、cox1(B)和18S rRNA+cox1(C)基因序列单倍型网络Fig.3 Network maps of 18S rRNA(A),cox1(B)and 18S rRNA+cox1(C)haplotypes of T.luwenshun from western Hunan
2.6 湘西地区吕氏泰勒虫的种系发育分析
湘西地区吕氏泰勒虫的种系发育分析结果见图4。从图4可以看出,该系统发育树共包含3个分支(CladeⅠ—Ⅲ),其中分枝CladeⅡ由16 株来自于湘西地区的羊源吕氏泰勒虫与来源甘肃的吕氏泰勒虫株Li2及Li6共同形成,而马泰勒虫则单独形成系统发育树的分支CladeⅠ,尤氏泰勒虫与小泰勒虫则共同形成该系统发育树的分支Clade Ⅲ。此外,CladeⅡ与Clade Ⅲ位于系统发育树的一大分支上,与CladeⅠ相分离。
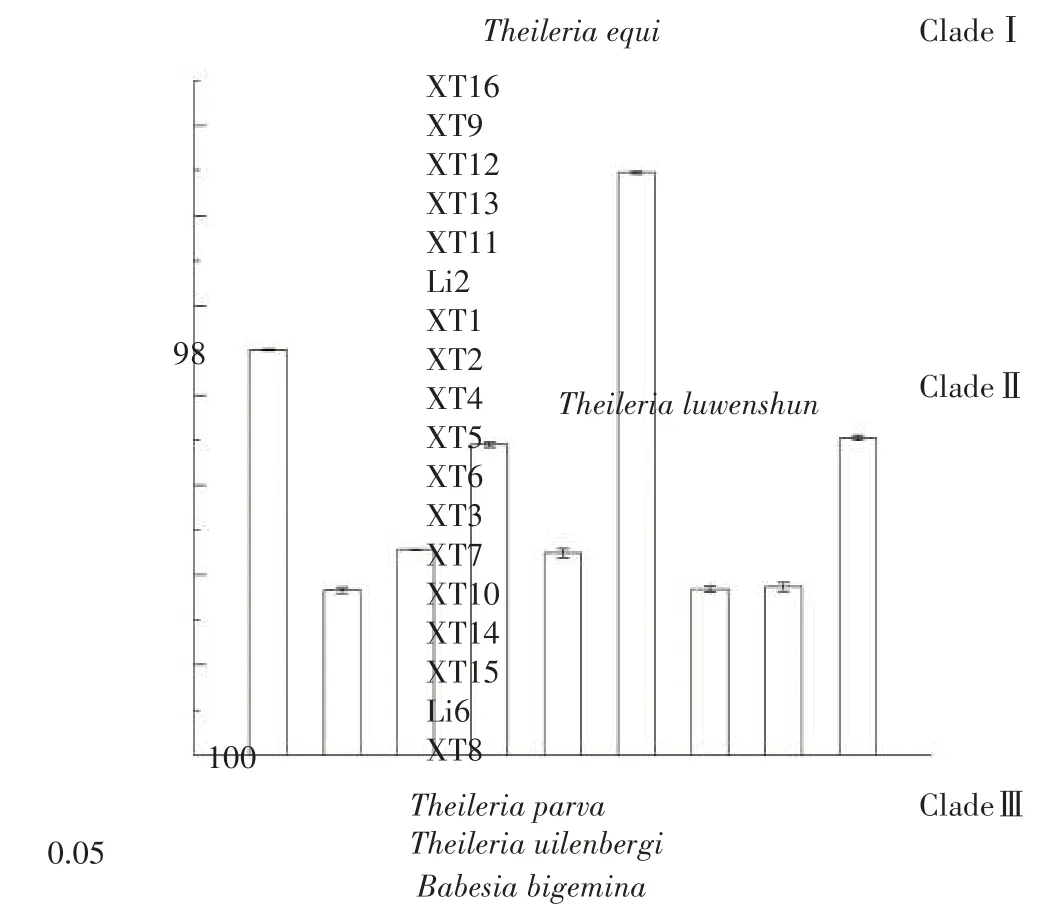
图4 湘西地区吕氏泰勒虫种系发育树Fig.4 Phylogenetic tree of T.luwenshun from western Hunan
3 结论与讨论
变异位点分析结果显示,湘西地区吕氏泰勒虫cox1基因序列(1 092 bp)共包含2个变异位点,且均为转换型,而在18S rRNA 基因序列(384 bp)中,共包含9个变异位点,7 个为转换型,2 个为颠换型。两者相比,发现cox1基因序列所包含的变异位点显著低于18S rRNA基因序列,这与前人研究结果相一致[23,31],也与核糖体基因具有高突变的特点相符合[32‐33],表明cox1基因序列的保守性高于18S rRNA基因序列,暗示cox1基因序列更适合用于物种的遗传变异分析,而18S rRNA因具有种内差异大的特点不适于遗传变异分析,这与林瑞庆等[34]认为ITS 存在种内差异大、不适合作为犬复孔绦虫种的遗传标记相吻合。
根据吕氏泰勒虫18S rRNA、cox1及18S rRNA+cox1基因序列的单倍型多样性的分析结果,发现18S rRNA、cox1及18S rRNA+cox1基因序列的单倍型均未出现聚集成群的现象和单倍型多样性的值均较低,且存在多个共同古老的单倍型(A1、A4、B1、B3、B4、C1、C8、C9),说明这16 株来源于湘西地区的羊源吕氏泰勒虫单倍型之间关系较近,并拥有共同的祖先,存在吕氏泰勒虫18S rRNA及cox1基因的古老序列,表明16株来源于湘西地区的羊源吕氏泰勒虫之间虽有一定程度的基因变异和单倍型多样性,但尚未出现遗传分化的现象。
在系统发育树中,来自湘西地区的16株羊源性吕氏泰勒虫与Li2及Li6两虫株相互掺杂在一起,共同形成该系统发育树的分枝Clade Ⅱ,这说明湘西地区的16株羊源吕氏泰勒虫之间及与Li2、Li6两虫株之间的亲缘关系较近;所选的尤氏泰勒虫及小泰勒虫则共同形成该系统发育树的Clade Ⅲ,这说明尤氏泰勒虫与小泰勒虫之间的亲缘关系近;而分支Clade Ⅱ虽与Clade Ⅲ相分离,但均位于该系统发育的一个大分支上;由马泰勒虫单独所形成的分支Clade Ⅰ却独立位于系统发育树的另一个大分支上,这表明吕氏泰勒虫与尤氏泰勒虫、小泰勒虫的亲缘关系较马泰勒虫近。以上结果表明,来自湘西地区的16 株羊源性吕氏泰勒虫之间及与外域性虫株的亲缘关系近,无遗传分化现象,与上述种内差异和单倍型多样性分析的结果相一致。此外,该结果也表明,吕氏泰勒虫与尤氏泰勒虫、小泰勒虫的亲缘关系近于马泰勒虫,且尤氏泰勒虫与小泰勒虫之间具有较近的亲缘关系。
本研究中,来自湘西地区的16株羊源吕氏泰勒虫之间虽存在一定程度的基因变异、遗传多样性及单倍型多样性,但它们个体之间、单倍型之间及与外域性吕氏泰勒虫株(Li2、Li6)之间存在较近的亲缘关系,尚未出现遗传分化的现象。
猜你喜欢
分子催化(2022年1期)2022-11-02
中国农业科学(2022年16期)2022-09-19
数理报(学习实践)(2021年5期)2021-04-07
电脑报(2020年40期)2020-11-06
金桥(2020年9期)2020-10-27
电脑知识与技术(2018年19期)2018-11-01
文艺论坛(2017年4期)2017-05-08
发明与创新·中学生(2016年8期)2016-05-14
岷峨诗稿(2014年3期)2014-11-15
英语学习·新锐空间(2013年1期)2013-05-08
