
冷藏小龙虾优势腐败菌的筛选鉴定及细菌群落结构和多样性分析
2022-07-06 18:43:32孟凌云戴泽川李娇毛相朝
肉类研究 2022年6期
关键词:高通量测序
孟凌云 戴泽川 李娇 毛相朝

摘 要:以4 ℃贮藏小龙虾货架期终点筛选武汉孝感小龙虾中优势腐败菌,同时结合高通量測序方法,进一步确定小龙虾中优势腐败菌,并对小龙虾贮藏过程中菌相变化、微生物结构及其多样性进行分析。结果表明:传统培养法从腐败小龙虾中共筛选出10 株腐败菌,包括波罗的海希瓦氏菌(Shewanella baltica)、维氏气单胞菌(Aeromonas veronii)、普通变形杆菌(Proteus vulgaris)等;在属水平上,高通量测序法最终确定气单胞菌属(Aeromonas sp.)
及希瓦氏菌属(Shewanella sp.)是小龙虾腐败末期相对丰度最大的2 种优势腐败菌;同时,高通量测序结果表明,随着贮藏时间的延长,小龙虾微生物群落丰富度、多样性及均匀度不断下降,这与腐败末期优势腐败菌的大量增长及生长繁殖过程中抑制其他种类微生物有关。
关键词:冷藏小龙虾;优势腐败菌;传统培养基法;高通量测序;微生物多样性
Screening and Identification of Dominant Spoilage Organisms and Analysis of Bacterial Community Structure and Diversity in Refrigerated Crayfish
MENG Lingyun1, DAI Zechuan1, LI Jiao1,*, MAO Xiangzhao1,2
(1.College of Food Science and Engineering, Ocean University of China, Qingdao 266003, China; 2.Qingdao National Pilot Laboratory for Marine Science and Technology/Laboratory for Marine Drugs and Biologiacal Products, Qingdao 266237, China)
Abstract: At the end of the shelf life of crayfish stored at 4 ℃, the dominant spoilage organisms in crayfish from Xiaogan, Wuhan were isolated and identified by high-throughput sequencing, and the changes in the bacterial community structure and diversity in crayfish during storage were analyzed. The results showed that 10 spoilage bacterial strains were obtained by the traditional culture-dependent method, including Shewanella baltica, Aeromonas veronii and Proteus vulgaris, etc. Aeromonas sp.
and Shewanella sp. were confirmed as the dominant spoilage organisms, with the largest relative abundance at the end of crayfish spoilage. At the same time, the results of high-throughput sequencing showed that the abundance, diversity and evenness of the microbial community decreased with storage time, which was related to the rapid growth of the dominant spoilage organisms and the inhibition of other kinds of microorganisms at the end of the spoilage stage.
Keywords: refrigerated crayfish; dominant spoilage organism; traditional culture-dependent method; high-throughput sequencing; microbial diversity
DOI:10.7506/rlyj1001-8123-20220505-050
中图分类号:TS254.4 文献标志码:A 文章编号:1001-8123(2022)06-0016-07
引文格式:
孟凌云, 戴泽川, 李娇, 等. 冷藏小龙虾优势腐败菌的筛选鉴定及细菌群落结构和多样性分析[J]. 肉类研究, 2022, 36(6): 16-22. DOI:10.7506/rlyj1001-8123-20220505-050. http://www.rlyj.net.cn
MENG Lingyun, DAI Zechuan, LI Jiao, et al. Screening and identification of dominant spoilage organisms and analysis of bacterial community structure and diversity in refrigerated crayfish[J]. Meat Research, 2022, 36(6): 16-22. DOI:10.7506/rlyj1001-8123-20220505-050. http://www.rlyj.net.cnFE2EA4C7-3244-48AE-9847-AEA3170429DE
小龙虾,也称克氏原螯虾(Procambarus clarkii),属于淡水经济虾类,原产于北美洲地区,19世纪30年代引入我国[1-2]。因其风味独特、肉质鲜美,同时具有高蛋白、低热量的特性[3],受到广大消费者的喜爱,成为人们餐桌上常见的美食。水产品由于营养丰富且水分含量较高而容易引起细菌的增殖进而腐败,难以保存[4-6]。而小龙虾好温喜湿,常见于淤泥等复杂生长环境,因此其本身携带的微生物种类和数量更多。同时,小龙虾含有大量的自由氨基酸及内源氧化酶,因此相较于其他水产品而言,更容易遭到微生物的侵袭[7-8],这些都在一定程度上影响了小龙虾产业的发展。
大量研究发现,在水产品腐败过程中部分种类细菌的数量逐渐上升并占据绝对优势地位,对水产品的腐败变质起着主导作用,这些细菌被称为优势腐败菌[9],其为水产品品质下降最重要的原因。不同水产品的优势腐败菌种类会存在一些差异,于淑池等[10]发现,冷藏卵形鲳鲹中优势腐败菌为希瓦氏菌及假单胞菌,Leroi等[11]在4 ℃冷藏生鲑鱼中发现其优势腐败菌主要为假单胞菌,南美白对虾中主要腐败菌则为不动杆菌、希瓦氏菌[12],而淡水水产品,如鲢鱼和草鱼中的优势腐败菌主要为气单胞菌[13]和假单胞菌[14]。對于水产品中优势腐败菌的筛选和鉴定,在早期的研究中一般都是利用传统培养法
进行[15-16]。随着分子生物学的进步与发展,目前研究中广泛利用传统培养法结合高通量测序法对优势腐败菌进行筛选和鉴定。同时,高通量测序方法还可以对贮藏过程中水产品的微生物结构组成及其多样性进行分析,目前已在带鱼[17]、牡蛎[18]等水产品中成功应用,明确了微生物在产品贮藏过程中的变化趋势。
本研究筛选和鉴定武汉孝感小龙虾4 ℃贮藏过程中的优势腐败菌,并对小龙虾贮藏过程中菌相变化以及微生物菌群结构及其多样性进行分析。优势腐败菌是水产品品质的关键控制点,与水产品货架期密切相关,因此开展对小龙虾中优势腐败菌的研究与筛选鉴定,有利于下一步利用杀菌保鲜技术靶向杀灭优势腐败菌,提高保鲜效果,延长小龙虾货架期。
1 材料与方法
1.1 材料与试剂
小龙虾来源于武汉孝感,经加冰保活运送至实验室备用,进行实验操作前加冰猝死。
氯化钠、硼酸、氧化镁、盐酸 国药集团化学试剂有限公司;营养琼脂(nutrient agar,NA)、假单胞菌选择性琼脂(Pseudomonas CFC selective agar,CFC)培养基、三糖铁琼脂(triple sugar iron agar,TSI)培养基、BP(Baird-Parker)培养基、气单胞菌基础(Aeromonas medium base,RYAN)培养基 青岛海博生物技术有限公司。
1.2 仪器与设备
K9840自动凯氏定氮仪 海能仪器有限公司;JY2002电子天平 梅特勒-托利多仪器(上海)有限公司;
TS-2402CL恒温振荡器 上海天呈实验仪器制造有限
公司;GXM-358恒温培养箱 宁波江南仪器制造厂;GI80TW全自动高压灭菌器 南京庚辰仪器科学有限公司。
1.3 方法
1.3.1 小龙虾贮藏期间菌落总数的测定
采用活菌平板计数法,测定小龙虾中总需氧菌菌落总数,测定方法参考GB 4789.2—2016《食品安全国家标准 食品微生物学检验 菌落总数测定》。称取10 g虾肉,加入90 mL无菌生理盐水,在无菌条件下匀浆,制成10 g/100 mL样品均质液。之后用生理盐水进行10 倍梯度稀释。取稀释后的样品100 μL涂布于NA平板上,于30 ℃恒温培养箱中培养36~48 h,选取合适的平板进行菌落计数,单位为lg(CFU/g)。
1.3.2 小龙虾贮藏期间总挥发性盐基氮(total volatile basic nitrogen,TVB-N)含量的测定
每隔2 d测定小龙虾中TVB-N含量,测定方法参照GB 5009.228—2016《食品安全国家标准 食品中挥发性盐基氮的测定》中的方法二自动凯氏定氮仪法。准确称取2 g虾肉糜于蒸馏管中,之后加入75 mL纯水,振荡使样品均匀分散,并在水中浸渍30 min。测定前加入1 g MgO,将蒸馏管连接至自动凯氏定氮仪。以20 g/L硼酸为接收液,0.01 mol/L盐酸为滴定液,1 g/L甲基红-乙醇溶液和1 g/L溴甲酚绿-乙醇溶液以体积比1∶5混合作为混合指示剂。
1.3.3 腐败菌单菌落的分离及纯化
将鲜活小龙虾碎冰猝死后,置于4 ℃条件下贮藏,选择贮藏腐败终点的小龙虾样品按照1.3.1节中的方法制成菌悬液,进行10 倍梯度稀释,选择合适稀释倍数菌液均匀涂布于NA、CFC培养基、TSI培养基、BP培养基、RYAN培养基,置于30 ℃恒温培养箱中孵育48 h,在选择性培养基上挑选不同颜色、形态的菌落,挑取单菌落并进行2~3 次划线纯化,并于-80 ℃保存。
1.3.4 腐败菌菌种的鉴定
1.3.4.1 分子生物学鉴定及系统发育树的构建
腐败菌接种于营养肉汤培养基中孵育至对数稳定期,提取细菌DNA并进行16S rDNA的聚合酶链式反应(polymerase chain reaction,PCR)扩增,最终产物送往北京擎科生物科技有限公司进行测序。将测序结果在美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)与Gen Bank库中进行BLAST序列比对。选取相似性在98%以上的菌株序列,利用MEGA 7.0.26软件中的邻近法(Neighbor-Joining)构建系统发育树。
1.3.4.2 生理生化实验鉴定FE2EA4C7-3244-48AE-9847-AEA3170429DE
根据《伯杰氏细菌鉴定手册》等,应用一系列生理生化实验,将筛选得到的腐败菌菌种作进一步鉴定。
1.3.5 高通量测序法测定小龙虾中优势腐败菌
选取在4 ℃贮藏0 d(鲜样期)、8 d(腐败初期)、12 d(腐败中期)、16 d(腐败末期)的小龙虾,取虾肉进行制样,得到菌悬液。提取样品菌悬液的DNA,并进行PCR扩增,将检测合格后的DNA样品送至上海凌恩生物科技有限公司进行高通量测序。
1.4 数据处理
每组实验设置3 次平行,结果用平均值±标准差表示。利用SPSS 17.0软件对数据进行差异显著性分析,差异显著性标准为P<0.05,采用Origin 7.5软件作图。
2 结果与分析
2.1 小龙虾4 ℃贮藏期间菌落总数及TVB-N含量变化
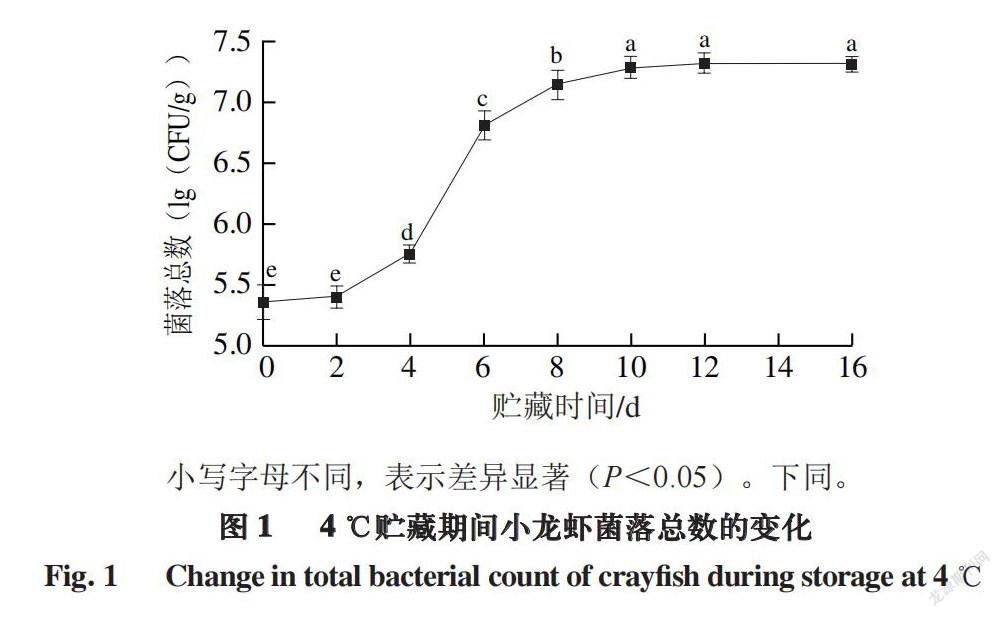
小写字母不同,表示差异显著(P<0.05)。下同。
水产品及肉类食物容易在微生物及酶的作用下发生蛋白质分解、脂质氧化等生化反应,进而引起腐败变质[19]。
因此可以通过测定小龙虾肉中的菌落总数来初步判断小龙虾被细菌污染及腐败的程度[20],反映虾的品质[21-22]。
由图1可知,小龙虾在4 ℃贮藏过程中的菌落总数随着时间的延长呈现上升的趋势,贮藏0~2 d,其增长趋势比较缓慢,说明小龙虾并没有发生明显的腐败。而贮藏2~10 d,菌落总数急剧增加,说明小龙虾在这个过程中发生严重腐败。贮藏6 d左右,小龙虾菌落总数超过无公害水产品安全限值(6.0(lg(CFU/g))),表明此时小龙虾腐败严重,到达了其货架期的终点。江杨阳等[23]
对江苏盱眙产地小龙虾研究发现,菌落总数也在6 d超过
6.0(lg(CFU/g))。除此之外,Li Yan等[24]发现,南美白对虾同样在贮藏6 d菌落总数超过了限值。本实验中的小龙虾在贮藏10 d之后,菌落总数不再明显增长,逐渐进入稳定期。
TVB-N是在微生物和酶的双重作用下分解蛋白质等含氮物质产生的生物胺、氨等挥发性碱性代谢物质的总称[25],其含量是评价水产品及肉类品质最重要的指标,是判断水产品新鲜程度及是否腐败变质的重要因素[26]。由图2可知,在整个贮藏过程中,TVB-N含量整体呈现上升趋势,前期增长比较缓慢,在贮藏后期随着菌落总数的快速增长、腐败加剧,导致了TVB-N含量增加速率加快[27]。
小龙虾TVB-N含量在貯藏6 d时已达43.28 mg/100 g,与秦求思等[28]测得的4 ℃贮藏6 d的鹰爪虾TVB-N含量相近,超过GB 10136—2015《食品安全国家标准 动物性水产制品》中所规定的淡水水产品TVB-N含量限定值30 mg/100 g,并且伴随强烈的动物腐烂的氨臭味。这与小龙虾菌落总数超过限值的贮藏时间相一致,表明TVB-N含量变化与菌落总数密切相关,结合2 个指标结果综合分析得出,小龙虾在贮藏6 d时已经腐败。
2.2 优势腐败菌单菌落的分离
选择在4 ℃贮藏至腐败的小龙虾进行平板涂布、划线,根据菌落形态及颜色差异在5 种固体培养基上分离纯化出10 株细菌,编号依次为B1~B10。10 株细菌菌落形态及颜色具体见表1。
2.3 腐败菌菌种的鉴定
2.3.1 腐败菌菌株分子生物学鉴定
对菌株B1~B10进行16S rDNA测序后,登录NCBI网站进行BLAST序列比对,并选取Max ident在98%以上的菌株,构建系统发育树,根据系统发育树(图3)及测序对比结果,将B5、B6、B7、B9及B10细菌定种,分别为中间气单胞菌(Aeromonas media)、巨型球菌(Macrococcus caseolyticus)、解淀粉芽孢杆菌(Bacillus amyloliquefaciens)、蜂房哈夫尼亚菌(Hafnia alvei)及缺陷短波单胞菌(Brevundimonas diminuta)。
2.3.2 生理生化实验鉴定
根据测序结果及系统发育树已对部分菌株定种,对仍无法定种的几种腐败菌进行生理生化实验,结果如表2所示,最终确定菌株B1~B4及B8的种类,分别为普通变形杆菌(Proteus vulgaris)、波罗的海希瓦氏菌(Shewanella baltica)、弗氏柠檬酸杆菌(Citrobacter freundii)、维氏气单胞菌(Aeromonas veronii)和嗜水气单胞菌(Aeromonas hydrophila)。
2.4 4 ℃贮藏小龙虾高通量测序结果分析
2.4.1 小龙虾4 ℃贮藏期间微生物群落的α-多样性
α-多样性分析可以反映小龙虾样品中微生物的多样性(丰富度及均匀度),由多种指数进行评价,主要包括反映微生物群落丰富度的Ace指数、Chao 1指数和Richness指数,可以体现微生物群落多样性的Shannon指数和Simpson指数,以及覆盖率[29]。
由表3可知,Ace指数、Chao 1指数及Richness指数随着贮藏时间的延长逐渐减小,这说明小龙虾样品在贮藏过程中微生物群落丰富度不断降低,微生物群落中物种种类逐渐减少,出现了优势菌属。而代表样品中微生物群落多样性的Shannon指数和Simpson指数也均降低,表示贮藏过程中小龙虾样品的微生物多样性也在逐步降低[30],物种均匀度下降。说明在小龙虾贮藏后期,出现了在数量上占绝对优势的细菌,并且这些细菌可能抑制了其他种类细菌的生长,从而降低了微生物群落的多样性。
稀释曲线体现对小龙虾样本的取样深度,可以用来评估测序量的合理性[31]。由图4A可知,随着测序深度的增加,小龙虾稀释曲线逐渐平缓,说明测序量足够且充分,测序样本能够合理反映样品中物种信息。由图4B可知,Shannon指数曲线也逐渐趋向平缓,代表微生物多样性不会再增加。同时表3中覆盖率均大于99%,因此,上述数据可以证明小龙虾样本的高通量测序数量合理,且能够正确反映出小龙虾样品中微生物的多样性及丰富度。FE2EA4C7-3244-48AE-9847-AEA3170429DE
2.4.2 小龙虾4 ℃贮藏期间细菌群落的β-多样性
对4 ℃贮藏小龙虾细菌群落进行基于操作分类单元(operational taxonomic units,OTU)的主成分分析(principal component analysis,PCA),PCA是通过降维处理展现样本特定距离的分布方法,PCA图中不同样品间距离的远近可以反映不同样本相似性的大小[32-33]。由图5可知,PC1、PC2的解释度分别为68.08%和20.03%。从样品分布可以看出,贮藏0 d样本与贮藏16 d的样本距离最远,说明其菌群组成差异最大。相对而言,贮藏8、12、16 d的样本则较为聚集,表示虽然这些样本之间存在一定的差异,但其菌群结构还是具有较高的相似度,而贮藏12 d与16 d样本距离最小,说明二者菌群结构相似度最高。小龙虾细菌菌群β-多样性的分析结果显示,小龙虾中微生物结构在贮藏过程中不断发生变化,最终造成鲜样期小龙虾与腐败末期小龙虾微生物结构之间产生较大的差异。
2.5 小龙虾贮藏期间微生物物种组成及聚类分析
对测序得到的序列进行聚类分析,以98%以上的序列聚类成OTU,并根据不同贮藏时间小龙虾样本所对应的OTU绘制维恩图。由图6可知,贮藏0、8、12、16 d样品含有的OTU数分别为2 830、1 379、1 121和977。贮藏12 d(腐败中期)与16 d(腐败末期)样品中相同的OTU数为674,高出贮藏0 d(鲜样期)与16 d(腐败末期)样品约500,因此可以说明腐败中期与腐败末期的小龙虾样品微生物物种相似度更高。
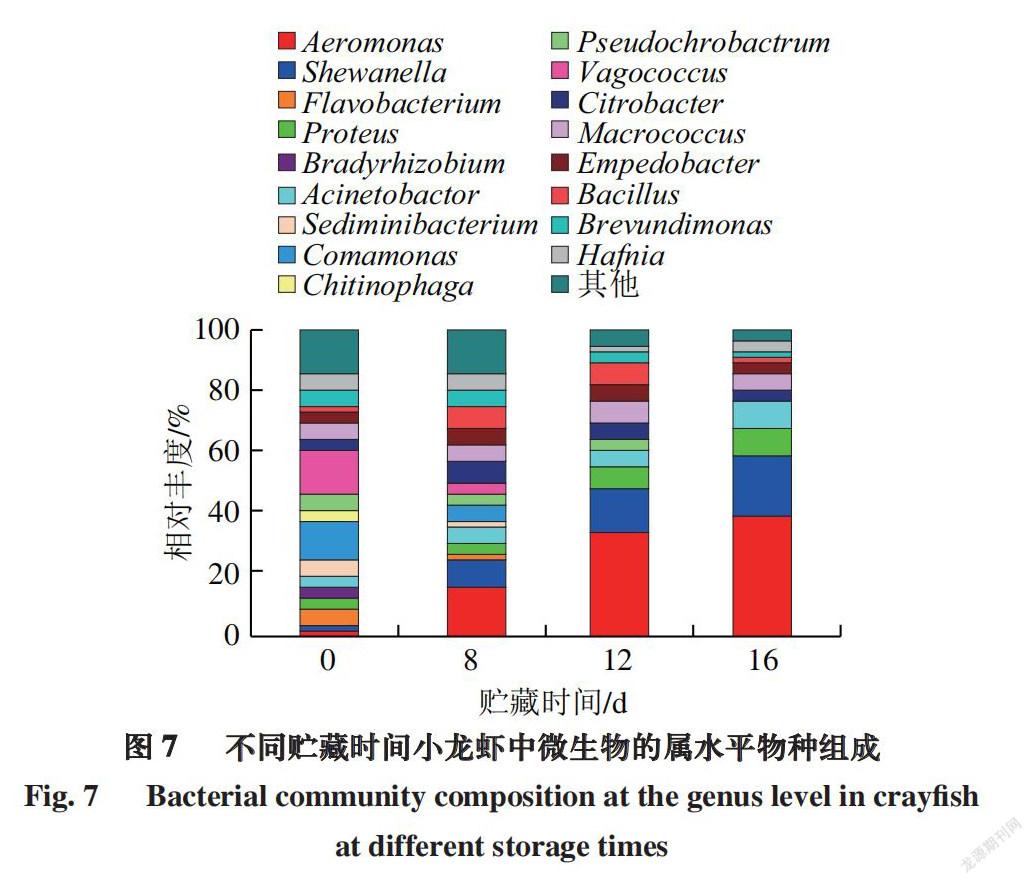
由图7可知,从整体上看,小龙虾样品中物种组成差异较大。总共涉及到17 个菌属,包括气单胞菌属(Aeromonas sp.)、希瓦氏菌属(Shewanella sp.)、黄杆菌属(Flavobacterium sp.)、变形杆菌属(Proteus sp.)、
不动杆菌属(Acinetobactor sp.)、丛毛单胞菌属(Comamonas sp.)、漫游球菌属(Vagococcus sp.)等,高通量测序法除传统培养法筛选到的10 株细菌外还鉴定出黄杆菌属、丛毛单胞菌属和漫游球菌属等7 个菌属,体现了高通量测序法的准确性及全面性。随着贮藏时间的延长,小龙虾鲜样期相对丰度较大的细菌,如丛毛单胞菌属、漫游球菌属逐渐减少,而气单胞菌属、希瓦氏菌属相对丰度则逐渐增加并成为优势菌群。在鲜样期(0 d),气单胞菌属、希瓦氏菌属相对丰度分别为1.40%、0.82%。而随着贮藏时间的延长,其相对丰度迅速增大,在贮藏末期(16 d),2 种细菌的相对丰度分别达到38.7%、18.9%。结合传统培养法在小龙虾中筛菌结果可知,本实验中小龙虾样品的优势腐败菌为气单胞菌及希瓦氏菌。黄佳奇[34]发现,冷藏小黄鱼的优势腐败菌为变形杆菌及希瓦氏菌,而朱素芹等[35]在凡纳滨对虾中筛选得到的优势腐败菌则为希瓦氏菌及不动杆菌,与本研究中产自武汉孝感的4 ℃贮藏小龙虾中筛选分离得到的优势腐败菌为希瓦氏菌及气单胞菌有所差异,可能是由于不同品种的水产品优势腐败菌有所差异。而本研究优势腐败菌种类与先前研究得出的小龙虾中优势腐败菌为希瓦氏菌、肉食杆菌[23]、阴沟肠杆菌[36]的结论也稍有不同,这说明即使是同一种水产品,其优势腐败菌也会因产地和贮藏温度的影响而发生变化。
3 结 论
本研究对产地为武汉孝感的冷藏小龙虾中优势腐败菌进行筛选鉴定,并对小龙虾贮藏过程中微生物结构变化及多样性进行分析。传统培养法从冷藏小龙虾中共筛选出维氏气单胞菌、波罗的海希瓦氏菌、普通变形杆菌等10 株细菌,结合高通量测序结果,最终确定小龙虾中主要优势腐败菌为气单胞菌属和希瓦氏菌属。同时,高通量测序结果表明,小龙虾在贮藏过程中微生物结构一直在变化。某些细菌数量逐渐减少,如在鲜样期(0 d)小龙虾中相对丰度较大的细菌丛毛单胞菌属在腐败末期小龙虾中相对丰度小于1%。而气单胞菌属、希瓦氏菌属相对丰度在贮藏过程中一直增大,并最终占据腐败小龙虾中微生物的主导地位。在腐败末期,气单胞菌属、希瓦氏菌属、变形杆菌属、不动杆菌属、柠檬酸杆菌属、巨球菌属6 种菌属的总相对含量为85%以上。α-多样性分析表明,小龙虾鲜样期OTU数量最多,菌群群落结构丰富度、多样性及均匀度最高。而随着贮藏时间的增长,多样性、丰富度及均匀度都在下降,尤其是在腐败末期到达最低,这可能与优势腐败菌的大量繁殖及繁殖过程中对其他细菌生长的抑制有关。β-多样性分析则表明,鲜样期与腐败末期菌群结构差异较大,而腐败中期与腐败末期差异较小,这与α-多样性分析結果相一致。对冷藏小龙虾优势腐败菌的筛选鉴定、细菌群落结构和多样性分析,为今后利用保鲜技术靶向杀灭腐败菌、延长货架期、降低经济损失提供了一定的理论参考,同时也为今后开发新的小龙虾品质安全检测及保鲜杀菌技术提供了思路。
参考文献:
[1] 张泽伟, 段伟文, 陈铭, 等. 过热蒸汽对熟制小龙虾优势腐败菌的杀菌动力学及其机理[J]. 广东海洋大学学报, 2020, 40(1): 87-96. DOI:10.3969/j.issn.1673-9159.2020.01.012.
[2] 王顺昌. 克氏螯虾的生物学和生态养殖模式[J]. 淡水渔业, 2003, 33(4): 59-61. DOI:10.3969/j.issn.1000-6907.2003.04.023.
[3] 吴晨燕, 王晓艳, 王洋, 等. 熟制麻辣小龙虾冷藏和冻藏条件下的品质变化[J]. 肉类研究, 2018, 32(5): 52-56. DOI:10.7506/rlyj1001-8123-201805009.
[4] 徐永霞, 白旭婷, 赵洪雷, 等. 植物乳杆菌在水产品中的应用研究进展[J]. 中国调味品, 2022, 47(2): 195-199. DOI:10.3969/j.issn.1000-9973.2022.02.040.FE2EA4C7-3244-48AE-9847-AEA3170429DE
[5] BRIONES L S, REYES J E, TABILO-MUNIZAGA G E, et al. Microbial shelf-life extension of chilled Coho salmon (Oncorhynchus kisutch) and abalone (Haliotis rufescens) by high hydrostatic pressure treatment[J]. Food Control, 2010, 21(11): 1530-1535. DOI:10.1016/j.foodcont.2010.04.027.
[6] ODUNAYO O O, SOOTTAWAT B. Natural preservatives for extending the shelf-life of seafood: a revisit[J]. Comprehensive Reviews in Food Science and Food Safety, 2018, 17(6): 1595-1612. DOI:10.1111/1541-4337.12390.
[7] LUO Zisheng, XU Yanqun, YE Qingyang. Effect of nano-SiO2-LDPE packaging on biochemical, sensory, and microbiological quality of Pacific white shrimp Penaeus vannamei during chilled storage[J]. Fisheries Science, 2015, 81(5): 983-993. DOI:10.1007/s12562-015-0914-3.
[8] HUANG Wanyou, JI Hongwu, LIU Shucheng, et al. Inactivation effects and kinetics of polyphenol oxidase from Litopenaeus vannamei by ultra-high pressure and heat[J]. Innovative Food Science and Emerging Technologies, 2014, 26: 108-115. DOI:10.1016/j.ifset.2014.10.005.
[9] GRAM L, DALGAARD P. Fish spoilage bacteria-problems and solutions: a review[J]. Current Opinion in Biotechnology, 2002, 3(13): 262-266. DOI:10.1016/S0958-1669(02)00309-9.
[10] 于淑池, 杨毅, 冯紫蓝, 等. 冷藏卵形鲳优势腐败菌的分离鉴定及致腐能力分析[J]. 食品工业科技, 2021, 42(1): 101-109. DOI:10.13386/j.issn1002-0306.2020030307.
[11] LEROI F, CORNET J, CHEVALIER F, et al. Selection of bioprotective cultures for preventing cold-smoked salmon spoilage[J]. International Journal of Food Microbiology, 2015, 213: 79-87. DOI:10.1016/j.ijfoodmicro.2015.05.005.
[12] 谢丽丹, 李蕾蕾, 王素英, 等. 低温贮藏南美白对虾特定腐败菌的分离鉴定及腐败能力分析[J]. 食品与发酵工业, 2016, 42(3): 67-72. DOI:10.13995/j.cnki.11-1802/ts.201603012.
[13] 丁婉. 半干草鱼片贮藏过程中优势腐败菌的研究[D]. 长沙: 中南林业科技大学, 2016: 50-60.
[14] 任阳阳. 鲢鱼卵类Cystatin的鉴定及其对冷藏鲢鱼肉片中假单胞菌J-4的抑制作用研究[D]. 成都: 四川农业大学, 2013: 50.
[15] 刘爱芳, 谢晶, 钱韻芳. 冷藏金枪鱼优势腐败菌致腐败能力[J]. 食品科学, 2018, 39(3): 7-14. DOI:10.7506/spkx1002-6630-201803002.
[16] OLAFSDOTTIR G, LAUZON H L, MARTINSDOTTIR E. Influence of storage temperature on microbial spoilage characteristics of haddock fillets (Melanogrammus aeglefinus) evaluated by multivariate quality prediction[J]. International Journal of Food Microbiology, 2006, 111(2): 112-125. DOI:10.1016/j.ijfoodmicro.2006.04.045.
[17] 高乾坤, 焦琳舒, 杜賀超, 等. 高通量测序分析不同产地带鱼冷藏时微生物群落多样性[J]. 食品科学, 2018, 39(18): 127-132. DOI:10.7506/spkx1002-6630-201818020.
[18] 曹荣, 刘淇, 赵玲, 等. 基于高通量测序的牡蛎冷藏过程中微生物群落分析[J]. 农业工程学报, 2016, 32(20): 275-280. DOI:10.11975/j.issn.1002-6819.2016.20.036.FE2EA4C7-3244-48AE-9847-AEA3170429DE
[19] 孔金花, 温丽敏, 诸永志, 等. 高温熟制杀菌对小龙虾品质及贮藏特性的影响[J]. 肉类研究, 2022, 36(3): 38-44. DOI:10.7506/rlyj1001-8123-20211210-236.
[20] 于晓慧, 葛孟甜, 林琳, 等. 一株常温保藏即食小龙虾优势腐败菌的分离及鉴定[J]. 肉类工业, 2018(5): 33-38.
[21] EMBORG J, LAURSEN B G, DALGAARD P. Significant histamine formation in tuna (Thunnus albacares) at 2 ℃-effect of vacuum-and modified atmosphere-packaging on psychrotolerant bacteria[J]. International Journal of Food Microbiology, 2005, 101(3): 263-279. DOI:10.1016/j.ijfoodmicro.2004.12.001.
[22] 徐麗敏, 薛长湖, 李兆杰, 等. 水溶性壳聚糖对南美白对虾品质及腐败菌相变化的影响[J]. 食品工业科技, 2008(6): 107-110. DOI:10.13386/j.issn1002-0306.2008.06.030.
[23] 江杨阳, 杨水兵, 余海霞, 等. 基于培养基法和高通量测序法分析冷藏小龙虾优势腐败菌[J]. 食品科学, 2019, 40(16): 130-136. DOI:10.7506/spkx1002-6630-20180718-229.
[24] LI Yan, LEI Yutian, TAN Yuqing, et al. Efficacy of freeze-chilled storage combined with tea polyphenol for controlling melanosis, quality deterioration, and spoilage bacterial growth of Pacific white shrimp (Litopenaeus vannamei)[J]. Food Chemistry, 2022, 370: 130924. DOI:10.1016/j.foodchem.2021.130924.
[25] 吴湛霞, 潘江球, 杨子明, 等. 混合分子质量壳聚糖结合臭氧杀菌处理对罗非鱼片的冷藏保鲜作用[J]. 广东海洋大学学报, 2016, 36(3): 71-75. DOI:10.3969/j.issn.1673-9159.2016.03.012.
[26] 许振伟, 李学英, 杨宪时, 等. 冷藏鲤鱼和罗非鱼优势腐败菌腐败能力分析[J]. 食品科学, 2012, 33(4): 243-246.
[27] ALPARSLAN Y, YAPICI H H, METIN C, et al. Quality assessment of shrimps preserved with orange leaf essential oil incorporated gelatin[J]. LWT-Food Science and Technology, 2016, 72: 457-466. DOI:10.1016/j.lwt.2016.04.066.
[28] 秦求思, 李思敏, 孟粉, 等. 基于鲜度的动力学模型预测鹰爪虾剩余货架期[J]. 中国食品学报, 2021, 21(7): 259-266. DOI:10.16429/j.1009-7848.2021.07.031.
[29] AMATO K R, YEOMAN C J, KENT A, et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes[J]. The ISME Journal, 2013, 7(7): 1344-1353. DOI:10.1038/ismej.2013.16.
[30] 徐圆程, 刘慧, 王光宇, 等. 基于高通量测序分析储藏稻谷中的真菌群落结构与优势菌属[J]. 食品科学, 2021, 42(24): 92-99. DOI:10.7506/spkx1002-6630-20201014-122.
[31] JESPERSEN L, JAKOBSEN M. Specific spoilage organisms in breweries and laboratory media for their detection[J]. International Journal of Food Microbiology, 1996, 33(1): 139-155. DOI:10.1016/0168-1605(96)01154-3.
[32] NELSON M C, MORRISON H G, BENJAMINO J, et al. Analysis, optimization and verification of Illumina-generated 16S rRNA gene amplicon surveys[J]. PLoS ONE, 2014, 9(4): e94249. DOI:10.1371/journal.pone.0094249.
[33] 庞春霞, 李艺, 虞任莹, 等. 基于Illumina Miseq技术比较不同地区传统发酵大豆制品细菌多样性[J]. 食品工业科技, 2022, 43(8): 133-140.
DOI:10.13386/j.issn1002-0306.2021070215.
[34] 黄佳奇. 小黄鱼优势腐败菌的分离鉴定及其与品质的相关性研究[D]. 杭州: 浙江大学, 2018: 24-26.
[35] 朱素芹, 张军宁, 曾惠, 等. 凡纳滨对虾优势腐败菌鉴定及其致腐能力的初步研究[J]. 食品科技, 2012, 37(1): 36-40. DOI:10.13684/j.cnki.spkj.2012.01.032.
[36] 邓灵, 赵康, 夏开, 等. 小龙虾(Procambarus clarkii)加工前后优势腐败菌的分离与鉴定[J]. 食品工业科技, 2020, 41(18): 100-104. DOI:10.13386/j.issn1002-0306.2020.18.016.
收稿日期:2022-05-05
基金项目:国家现代农业产业技术体系建设专项(CARS-48)
第一作者简介:孟凌云(1998—)(ORCID: 0000-0003-3185-2743),女,硕士研究生,研究方向为水产品加工及贮藏。
E-mail: mengly629@163.com
*通信作者简介:李娇(1992—)(ORCID: 0000-0002-6058-3725),女,副教授,博士,研究方向为水产品加工及贮藏。
E-mail: lijiao@ouc.edu.cnFE2EA4C7-3244-48AE-9847-AEA3170429DE
猜你喜欢
江苏农业科学(2017年20期)2017-11-30 14:53:29
江苏农业科学(2017年16期)2017-10-27 14:31:12
湖北农业科学(2017年13期)2017-08-08 05:09:44
中国中药杂志(2017年13期)2017-07-31 08:28:44
山东工业技术(2017年12期)2017-07-06 18:29:27
中国中药杂志(2017年7期)2017-05-26 00:05:40
中国医药导报(2017年9期)2017-05-11 18:44:13
中国中药杂志(2016年24期)2017-04-18 17:42:52
中国中药杂志(2017年4期)2017-03-28 17:06:08
中国中药杂志(2017年3期)2017-03-20 21:11:11
