
miR-204-5p通过调控IL-11对甲状腺乳头状癌细胞恶性生物学行为的机制研究
2022-07-04 06:04王玉文周雅琪宋宗义赵永丽
医学研究杂志 2022年6期
王玉文 周雅琪 宋宗义 赵永丽
甲状腺乳头状癌(papillary thyroid carcinoma,PTC)是甲状腺恶性肿瘤中最常见的组织类型,其发生率在过去几十年中一直在稳步上升[1,2]。尽管大多数病例具有良好的预后和治疗反应,但高达30%的患者在10年内出现局部复发或远处转移[3]。微小RNA(microRNA,miRNA)是一组具有18~25个序列,没有编码能力的单链RNA,可以通过与目标mRNA的3′非翻译区(untranslated region,UTR)结合,参与转录后基因表达[4]。miRNA调节细胞周期及多种基本生理过程,包括细胞凋亡、增殖、迁移、侵袭和分化,并且miRNA表达或功能在各种恶性肿瘤(包括PTC在内的)中失调[5,6]。miR-204-5p通过靶向HMGB1抑制甲状腺乳头状癌K1细胞增殖、侵袭和迁移[7]。最近的基因芯片的测序结果显示miR-204-5p在PTC中下调[8]。然而,miR-204-5p在PTC中的生物学功能和潜在机制未知。
白细胞介素11(interleukin 11,IL-11)是一种属于多功能IL-6细胞因子家族的造血细胞因子,在肿瘤的发生、发展和转移中起重要作用[9,10]。IL-11的高表达对乳腺癌、胃癌和细胞性肾细胞癌等恶性肿瘤细胞的增殖和侵袭具有促进作用[11~13]。本研究旨在探讨miR-204-5p在PTC中抗癌角色和生物学功能。
材料与方法
1.主要试剂和仪器:DMEM培养基购自美国Thermo Scientific公司;psiCHECK-2荧光素酶载体购自美国Promega公司;miR-204-5p模拟物(miR-204-5p)、miR-204-5p阴性对照(miR-neg)、miR-204-5p抑制剂(anti-miR-204-5p)和抑制剂对照(anti-miR-neg)购自南京凯基生物科技发展有限公司;Lipofectamine 3000购自美国Invitrogen公司;聚凝胺购自美国Sigma公司;miRNeasy Mini试剂盒购自德国Qiagen公司;SuperScript Ⅲ反转录试剂盒购自美国Invitrogen公司;ABI 7900HT荧光定量PCR仪购自美国Applied Biosystems公司;SYBR Premix Ex Taq Ⅱ购自中国大连TaKaRa公司;MTT试剂盒购自美国Invitrogen公司;IX81倒置显微镜购自日本Olympus公司;FACScan流式细胞仪购自美国BD Biosciences公司;Annexin V-FITC/PI细胞凋亡检测试剂盒购自美国BD Biosciences公司;双荧光素酶报告系统购自美国Promega公司;BCA试剂盒购自北京索莱宝生物工程有限公司;白细胞介素11(interleukin 11,IL-11)、甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)购自英国Abcam公司;辣根过氧化物酶偶联的山羊抗兔IgG购自美国Santa Cruz Biotechnology公司;ECL发光系统购自美国Amersham Biosciences公司。
2.细胞培养:人PTC细胞系(TCP-1和BCPAP)、人甲状腺正常细胞NTHY-ORI 3-1和HEK293T购自美国ATCC细胞库,在含有10%胎牛血清的DMEM培养基中于37℃和5% CO2中培养。
3.载体构建:miR-204-5p、miR-neg、anti-miR-204-5p和anti-miR-neg从商业公司获得,并通过测序验证结果。根据说明书,Lipofectamine 3000用于转染。转染序列如下:miR-204-5p: 5′-UUCCCUUUGUCAUCCUAUGCCU-3′; anti-miR-204-5p: 5′-AGGCAUAGGAUGACAAAGGGAA-3′。
4.慢病毒转染:将含有pre-miR-204-5p的450bp片段克隆到pWPXL载体中以生成pWPXL-miR-204。使用Lipofectamine 3000将pWPXL或pWPXL-miR-204-5p构建体与所对应的包装质粒psPAX2或pMD2G共转染到HEK293T细胞中,48h后收集病毒颗粒。TPC-1细胞用含有8mg/ml聚凝胺的重组慢病毒转染。通过qRT-PCR检测稳定的转染基因表达。
5.qRT-PCR:根据制造商的说明,使用miRNeasy Mini试剂盒提取总RNA和miRNA。SuperScript Ⅲ反转录试剂盒用于将RNA转录成cDNA,并在ABI 7900HT荧光定量PCR仪上使用SYBR Premix Ex Taq Ⅱ进行qRT-PCR。将靶基因表达水平标准化为内部对照(U6或GAPDH)的水平,并由2-△△CT计算比较差异。引物序列如下:miR-204-5p 上游引物:5′-ACAATTGAACGTCCCTTTGCC-3′,下游引物:5′-CTCAACTGGTGTCGTGGA-3′;IL-11上游引物:5′-GGGAATTTGTCCCTCTGGCA-3′,下游引物:5′-CTGGGACTTAAGACCTCCAGC-3′;GAPDH 上游引物::5′-GCAACTAGGATGGTGTGGCT-3′,下游引物:5′-TCCCATTCCCCAGCTCTCATA-3′;U6上游引物:5′-CTAGATAATGGTGCTGATAGATGGA-3′,下游引物:5′-CGGTATAGCTGGACGCTCTG-3′。
6.细胞增殖和平板克隆试验:使用MTT法测定法评估细胞活力,具体参照文献所述[14]。简而言之,将2×103个细胞培养在96孔板中,并在24、48、72和96h用20ml MTT(0.5mg/ml)染色4h。弃上清液,每孔加入200ml DMSO溶解沉淀。在490nm处测量吸光度。对于平板克隆试验,将细胞以2×102个/孔接种在6孔板中并生长2周。形成的菌落用PBS洗涤,甲醇固定,0.5%结晶紫染色。在IX81倒置显微镜下计数菌落数。
7.细胞周期和细胞凋亡分析:对于细胞周期分析,收集2×105个转染细胞,用70%乙醇固定并在4℃下孵育过夜。洗涤后,细胞用RNase A(100mg/ml)处理30min,然后用PI(50mg/ml)在暗室中染色30min。使用FACScan流式细胞仪分析细胞周期分布。使用Annexin V-FITC/PI细胞凋亡检测试剂盒评估细胞凋亡。
8.双荧光素酶报告基因分析:HEK293T细胞在96孔板中培养,并用50ng野生型(WT)或突变(MUT)IL-11 3′-UTR构建体或50mmol/L miR-neg或miR-204-5p mimic共转染。48h后收集细胞,并用双荧光素酶报告系统测量海肾荧光素酶活性。
9.Western blot法检测:裂解细胞并使用BCA法测量蛋白质浓度。变性蛋白30mg通过12% SDS-PAGE分离并转移到PVDF膜上,用5% 脱脂牛奶在Tris缓冲盐水中室温封闭1h,并与一抗孵育IL-11和GAPDH在4℃过夜,然后在常温下孵育二抗辣根过氧化物酶偶联的山羊抗兔IgG。通过化学ECL发光显影蛋白质条带。
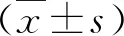
结 果
1.miR-204-5p在TPC-1和BCPAP细胞中表达下调:笔者评估了正常人甲状腺细胞NTHY-ORI 3-1和两种PTC细胞中miR-204-5p的表达水平。qRT-PCR的结果显示,与正常细胞比较,TPC-1和BCPAP细胞系中的miR-204-5p水平显著降低(P<0.05,图1)。这些数据表明miR-204-5p在PTC细胞中显著下调。

图1 miR-204-5p在TPC-1和BCPAP细胞中表达下调
2.miR-204-5p抑制TPC-1细胞增殖:由于miR-204-5p在TPC-1的表达水平更低,因此笔者选择TPC-1进行后续试验。随后笔者探索了miR-204-5p在TPC-1细胞中的生物学功能。TPC-1细胞用表达带有miR-204-5p或者miR-neg的慢病毒感染,qRT-PCR验证显示,miR-204-5p在转染细胞中的表达显著增加(P<0.05,图2)。MTT结果显示,与转染miR-neg细胞比较,细胞存活率明显降低(P<0.05,图3A),并且在平板克隆试验中也显示了转染miR-204-5p可以降低细胞增殖率(P<0.05,图3B)。

图2 TPC-1细胞过表达处理后miR-204-5p表达水平

图3 miR-204-5p抑制TPC-1细胞增殖
3.miR-204-5p促进TPC-1细胞凋亡和细胞周期阻滞:流式细胞术结果显示与miR-neg转染细胞比较,转染miR-204-5p增加了TPC-1的细胞凋亡率(P<0.05,图4A)。并上调G0/G1的比值(P<0.05)和降低了S期细胞的比例(P<0.05,图4B),表明miR-204-5p促进了TPC-1在S2期的细胞阻滞。
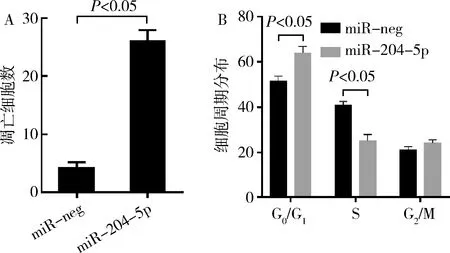
图4 miR-204-5p促进TPC-1细胞凋亡和细胞周期阻滞
4.miR-204-5p通过靶向结合3′-UTR抑制IL -11表达:TargetScanHuman 7.1预测了IL-11 3′-UTR与miR-204-5p存在高度保守的结合位点(图5A)。双荧光素酶报告分析结果显示,miR-204-5p的转染降低了野生型IL-11 3′-UTR的HEK293T细胞中的相对荧光素酶活性(P<0.05),但在转染了3′-UTR且miR-204-5p突变结合位点的细胞中,该活性基本未改变(图5B)。

图5 miR-204-5p与IL-11 3′-UTR存在靶向关系
5.miR-204-5p与IL-11存在负性调节关系:qRT-PCR的结果显示过表达miR-204-5p会降低IL-11 mRNA的水平(P<0.05),而敲减miR-204-5p则显示出相反的结果(P<0.05,图6),与mRNA的结果一致,蛋白表达也观察到了类似的结果。

图6 miR-204-5p与IL-11存在负性调节关系
讨 论
miRNA通过调节靶基因表达在癌症的发生、发展中发挥关键作用,它们可以在功能上区分为促癌因子和抑癌因子[15,16]。作为一种肿瘤抑制因子,miR-204-5p在结直肠癌中下调并与不良预后相关,过表达miR-204-5p可抑制结直肠癌细胞增殖、迁移和侵袭并诱导上皮间质转化[17]。miR-204-5p可抑制子宫内膜癌细胞的克隆生长、迁移和侵袭以及移植瘤体的生长[18]。本研究中miR-204-5p在PTC细胞中显著降低。生物学功能分析表明,miR-204-5p可以抑制细胞活力和增殖能力,阻断细胞周期进程,增强细胞凋亡。
Wang等[19]研究发现,IL-11通过NF-κB/microRNA-204/血小板生成素调节轴促进了造血干细胞移植疗法在再生障碍性贫血小鼠模型中的治疗效果。miR-204-5p可以影响食管癌细胞和小鼠模型中的增殖、侵袭、凋亡和细胞周期[20]。既往研究显示,IL-11与miR-204-5p存在负性调节关系。Tang等[20]研究表明,miR-204-5p与IL-11的表达呈负相关,而IL-11的过表达逆转了miR-204-5p在细胞系中的癌症抑制作用。Liang等[21]研究显示,IL-11是miR-204-5p的下游靶标,lncRNA脱氧鸟苷激酶反义RNA1(lncRNA deoxyguanosine kinase antisense RNA1,DGUOK-AS1)可以保护IL-11免受miR-204-5p介导的降解,从而起到促进乳腺癌的发展进程。在本研究中,笔者通过生物信息学预测和荧光素酶活性测定证实IL-11是miR-204-5p的靶标。发现通过过表达或敲减miR-204-5p的表达分别可以引起IL-11的下调或者上调,这表明了二者之前存在负性调节关系。在随后的研究中笔者还会在其他类型的PTC细胞上进行评估miR-204-5p的生物学功能并进行在体荷瘤实验。
综上所述,miR-204-5p作为肿瘤抑制因子,可能通过调控下游靶基因IL-11的表达来调控PTC细胞的生物学行为。结果表明,miR-204-5p可能是一种有用的诊断标志物和潜在的治疗分子。
猜你喜欢
中国农学通报(2022年12期)2022-06-01
作物学报(2022年4期)2022-02-10
三农资讯半月报(2021年1期)2021-01-27
中学生物学(2019年7期)2019-10-17
江苏农业科学(2019年23期)2019-03-03
中国中医药信息杂志(2018年11期)2018-01-05
中国医药导报(2016年33期)2017-03-06
三联生活周刊(2017年1期)2017-01-11
医学信息(2016年29期)2016-11-28
中学生物学(2016年10期)2016-11-19
