
干露胁迫对毛蚶稚贝生理代谢相关酶活力的影响
2022-07-04 09:20:00江天棋姜亚洲凌建忠唐保军
海洋渔业 2022年2期
张 扬,江天棋,姜亚洲,凌建忠,孙 鹏,唐保军
(上海海洋大学水产与生命学院,上海 201306;2.中国水产科学研究院东海水产研究所,上海 200090)
生活在潮间带的海水贝类易受干露、高温、低氧等多种环境因素影响[1],其中干露时间过长被视为潮间带贝类最致命的威胁[2-3]。干露胁迫下,潮间带贝类体内水分逐渐丧失[4-5],生长延缓甚至死亡[6-8]。研究发现,厚壳贻贝(Mytilus coruscus)稚贝在1~8℃下干露48 h后死亡率即达40%以上[9],硬壳蛤(Mercenaria mercenaria)稚贝干露48 h后死亡率达63.3%[6],中国蛤蜊(Mactra chinensis Philippi)稚贝在干露20 h后死亡率即达40%以上[10]。同时,因无法获得足够氧气,贝类代谢方式由有氧呼吸逐渐转为无氧呼吸,其代谢产物会对贝类造成损伤[4,11-12]。干露胁迫显著影响大竹蛏(Solen grandis)呼吸代谢酶的活力[13],并导致魁蚶(Scapharca broughtonii)幼贝出现显著的氨氮积累,显著影响其再次入水后的耗氧率和排氨率[4]。此外,有研究报道,干露胁迫下贝类体内非特异性免疫酶的活力,如酸性磷酸酶、超氧化物歧化酶等,也会发生变化[14]。因此,研究干露胁迫下潮间带贝类的生理代谢酶活力,有助于了解海水贝类的逆境适应机制。
毛蚶属于软体动物门,瓣鳃纲,翼形亚纲,蚶目,蚶科,毛蚶属,俗称毛蛤、麻蚶、瓦楞子等,在我国北起鸭绿江、南至海南均有分布,莱州湾、渤海湾、辽东湾、海州湾、象山港等区域资源尤为丰富,是重要的经济贝类。毛蚶生活史可分为3个阶段:浮游期、附着期、底栖期,当毛蚶壳长12~15 mm后开始底栖生活[15]。退潮后附着或底栖的毛蚶可能露出滩面暴露在空气中,毛蚶苗种的运输过程中也会遭受干露胁迫。本研究测定了干露胁迫下毛蚶稚贝的生理代谢相关酶活力,并以滤水率和耗氧率为指标评估干露胁迫后稚贝的恢复状况,以期为毛蚶耐干露胁迫生理指标的筛选和生理响应机制研究提供参考,并为毛蚶大规格苗种运输和增养殖提供指导。
1 材料与方法
1.1 实验材料
毛蚶稚贝采自浙江宁波象山港海域滩涂,取回实验室后挑选大小一致的个体[平均壳长(16.99±2.48)mm,平均体质量(1.98±0.46)g],清除表面附着物后放入2 000 L玻璃钢桶暂养3 d。暂养期间,海水盐度21.55±1.09,控制水温(20.0±0.5)℃,每天换水一次,投喂2次亚心形扁藻(Platymonas subcordiformis)。
1.2 失水率测定
将300只毛蚶稚贝平均分成3组,贝壳表面水分用吸水纸擦干,分别测定各组初始总质量,直接暴露在室内阴凉处空气中进行干露,空调控制室内温度为20℃,相对湿度为50%,采用自然光照。测定干露后2、4、8、12、24、36、48、72 h的总质量,用差减法计算单位个体干露失水率(如中间有死亡,记录死亡个数,去除死亡个体,继续称重,以毛蚶贝壳张开、触碰后无闭壳反应为标准认定死亡状态)[16]。
1.3 酶活测定
毛蚶稚贝干露48 h后死亡率超过33.3%,因此测定毛蚶干露前48 h的生理代谢相关酶活力。分别于干露2、6、12、24、48 h取稚贝,未干露稚贝设为对照组,在冰盘上迅速解剖取全组织,液氮冷冻后置-80℃冰箱保存。酶活力测定时,将组织和9倍体积的0.9%生理盐水放入预冷的匀浆机中,制成组织匀浆,以2 500 r·min-1离心15 min,取上清液测定丙酮酸激酶(PK)、己糖激酶(HK)、琥珀酸脱氢酶(SDH)、超氧化物歧化酶(SOD)和酸性磷酸酶(ACP)活力以及三磷酸腺苷(ATP)含量。
各酶活力均采用试剂盒(南京建成生物工程研究所)测定,具体测定方法按照试剂盒说明书。PK酶活力为1 min内1 g组织蛋白1μmol的磷酸烯醇式丙酮酸转变成丙酮酸为1个酶活力单位。HK酶活力定义为1 mg组织蛋白1 min生成1 nmol的烟酰胺腺嘌呤二核苷酸磷酸定义为1个酶活力单位。SDH酶活力定义为1 mg蛋白1 min使反应体系的吸光度降低0.01为1个比活力单位。SOD酶活力定义为1 mg组织蛋白在1 mL反应液中SOD抑制率达50%时所对应的SOD量为1个活力单位。ACP酶活力定义为30 min内产生1 mg酚为1个活力单位。采用考马斯亮蓝法测定上清液蛋白浓度。
1.4 耗氧率和滤水率测定
为研究干露胁迫后毛蚶稚贝呼吸代谢的恢复情况,取20℃下干露48 h的200只毛蚶稚贝放回水温20℃的海水中,测定滤水率和耗氧率。
滤水率实验在3 L广口瓶中进行,每瓶加亚心形扁藻10 mL,使藻细胞浓度约为1.0×104个·mL-1,每个处理组设置3个平行,每瓶放10只稚贝,未干露的稚贝为对照组,另设不放贝的空白瓶,用来校正实验瓶中藻浓度的变化。毛蚶贝壳张开后开始计时,每隔20 min取一次水样,连续取样3次,每次取水量为500 mL,以孔径为0.45μm的混合纤维素酯滤膜抽滤水样,测定叶绿素a(Chla)含量。实验结束后用游标卡尺测量实验贝生物学数据,解剖取软体部于65℃烘干24 h,称量组织干重。
滤水率按COUGHLAN[17]的方法计算,公式如下:
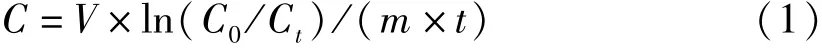
式(1)中,V为取样时瓶内海水体积(L),m为实验用贝软体部干重(g),C0和Ct分别为每两次取样间试验组瓶内Chla浓度(μg·L-1),t为取样间隔时间。
耗氧率采用500 mL玻璃呼吸瓶测定,呼吸瓶内放10只稚贝,每个时间点设3个平行,未做干露处理的毛蚶作为对照组,不放贝的呼吸瓶作为空白组。放入毛蚶5 min后开始计时,用溶解氧测定仪(雷磁JPBJ-609L)测定瓶内溶解氧浓度,读数间隔2 min,持续1 h。实验结束后用游标卡尺测量稚贝壳长,解剖取软体部于65℃烘干24 h,称量组织干重。
根据下式计算耗氧率:

式(2)中,R为单位体质量耗氧率,b为溶解氧浓度对时间线性回归的斜率绝对值(扣除空白组溶解氧浓度变化),V为呼吸瓶体积(L),m为稚贝软体部干重(g)。
1.5 数据处理
实验数据采用平均值±标准差(mean±SD)表示。利用Excel 2010软件进行数据处理、绘图,采用SPSS 19.0统计软件对数据进行单因素方差分析(one-way ANOVA),以P<0.05为差异显著。
2 结果与分析
2.1 干露胁迫对毛蚶稚贝失水率的影响
毛蚶失水率与干露时间的关系满足曲线方程y=-0.002 7x2+0.767 9x+0.379 5,R2=0.995 6(n=300,P<0.05)(图1)。失水率随着干露时间的延长而增大,干露12 h后毛蚶失水率达9.45%,48 h后失水率达32.38%。毛蚶稚贝在干露48 h后死亡率超过30%。

图1 干露胁迫对毛蚶失水率的影响Fig.1 Effect of air exposure on water loss rate of S.kagoshimensis
2.2 干露胁迫对毛蚶稚贝生理代谢相关酶活力的影响
在干露条件下,毛蚶组织PK活力在干露2 h显著上升,在6 h恢复到对照组水平,其后在12~24 h显著高于对照组,后下降到对照组水平(表1)。干露胁迫后,组织HK活力下降,处理后6、12、48 h活力显著低于对照组(P<0.05),在12 h降至最小值,2 h和24 h的活力与对照组相比差异不显著。毛蚶组织SDH活力先上升后下降。与对照组相比,处理后2~6 h SDH活力无显著变化,至12 h SDH活力显著升高(P<0.05),达到最大值,其后SDH活力下降,恢复到对照组水平,干露48 h的SDH 活力显著低于对照组(P<0.05)。
毛蚶组织ATP含量先上升后下降(表1)。干露处理后2 h,组织ATP含量与对照组差异不显著,6~12 h组织ATP含量显著高于对照组(P<0.05),在12 h达到最大值,其后开始下降,24~48 h恢复到对照组水平。

表1 干露胁迫对毛蚶代谢酶活力及ATP含量的影响Tab.1 Effect of air exposure on metabolic enzyme activity and ATP content of S.kagoshimensis
干露胁迫后,毛蚶组织SOD活力先下降后上升(表2)。干露处理后2 h,SOD活力与对照组相比无显著差异;处理后6~24 h,SOD活力显著低于对照组(P<0.05),在12 h降至最小值;在48 h SOD活力恢复到对照组水平。组织ACP活力先上升后下降。与对照组相比,干露后2 h的ACP活力无显著变化;处理后6~12 h,ACP活力显著高于对照组(P<0.05);24~48 h,ACP活力持续下降,恢复至对照组水平。

表2 干露胁迫对毛蚶免疫相关酶活力的影响Tab.2 Effect of air exposure on immune enzyme activity of S.kagoshimensis
2.3 放回水中后毛蚶稚贝滤水率和耗氧率变化
毛蚶稚贝放回水中后6~24 h,滤水率升高,但与对照组相比无显著变化,至48 h滤水率显著升高(P<0.05)(图2)。毛蚶放回水中后,耗氧率先下降后上升,最小值与最大值分别出现在12 h和24 h,各个时间点的耗氧率均与对照组差异不显著(图3)。

图2 干露48 h后放回水中毛蚶滤水率的变化Fig.2 Variation of filtration rates of S.kagoshimensis back to water after 48-hour air exposure

图3 干露48 h后放回水中毛蚶耗氧率的变化Fig.3 Variation of oxygen consumption rates of S.kagoshimensis back to water after 48-hour air exposure
3 讨论
3.1 失水率
水是生命活动的必须介质,细胞内的酶活反应以及其他化学反应都离不开水。毛蚶属于双壳贝类,在遭遇环境因子胁迫时会出现紧闭贝壳的现象[18]。本实验中,干露过程中毛蚶双壳关闭,通过这种方法来减少水分流失。不同种类贝类的闭合程度和厚薄不同,从而导致对干露的耐受能力不同[8]。本实验中,随着干露时间延长,毛蚶失水率逐渐增大,12 h的失水率为9.45%,24 h的失水率为17.88%,而干露24 h的文蛤(Meretrix meretrix)失水率仅为3%[16];这可能因为文蛤贝壳边缘光滑,闭合较紧,而毛蚶贝壳边缘呈锯齿状,易于蒸发水分。不同规格的贝类耐干露能力也不同,如海湾扇贝(Argopecten irradians)[19]的成贝耐干露能力大于稚贝和幼贝。据报道,壳长0.7~20.0 mm的毛蚶苗在20℃下干露15 h死亡率超过50%[20],壳长22.25~27.49 mm的毛蚶在18℃下干露3 d存活率仍接近100%[21],本实验中,平均壳长约17 mm的毛蚶稚贝在20℃下干露48 h死亡率超过30%,说明不同规格的毛蚶耐干露能力有差异,关于毛蚶成贝的耐干露能力需继续开展研究。
3.2 生理代谢酶活力
丙酮酸激酶是糖酵解过程中的主要限速酶之一,己糖激酶是糖酵解途径中控制葡萄糖代谢速率的第一个关键酶,在机体代谢过程中发挥着重要作用和调节功能[22]。本实验中,HK和PK两种酶的活力在12 h前后变化较大,说明随着干露时间的延长,稚贝可能通过调节两种酶的活力调控代谢方式和水平,提高葡萄糖的无氧氧化产生ATP,为机体提供能量,以抵抗干露胁迫对机体的影 响, 这 与 三 疣 梭 子 蟹 (Portunus trituberculatus)[23]、厚壳贻贝[24]的研究报道一致。
SDH负责把琥珀酸脱氢形成延胡索酸,是有氧氧化中重要的酶[22]。本实验中,SDH活力在12 h达到最大值,说明12 h是毛蚶有氧呼吸最旺盛的时间,毛蚶通过增强SDH活力来加速丙酮酸分解产生ATP,为机体抵抗干露胁迫提供能量。12 h后SDH活力逐渐下降,可能因为随着氧气摄入越来越少,毛蚶稚贝逐渐转入无氧呼吸。有研究表明,大竹蛏SDH酶活力随着干露时间的延长逐渐下降,超过24 h活力下降愈发明显[13],说明对海水贝类来说,12~24 h是干露耐受的重要节点[25]。
能量稳态对生物环境胁迫耐受性起着关键作用,能量平衡是动物胁迫适应的基本要求。能量的获取、转换与分配影响动物环境压力下的代谢适应。在不同的环境胁迫压力下,生物体往往表现出不同的能量代谢策略[3,26-27]。本实验中,在12 h之前毛蚶全组织ATP含量随干露时间延长而增加,在12 h达到最大值,与SDH活力的变化趋势一致,说明毛蚶通过提升有氧代谢水平,产生能量以应对不利环境带来的影响。12 h以后可能毛蚶体内转变为无氧代谢,有氧条件下1分子葡萄糖可产生32个ATP,而糖酵解只能产生4个ATP分子[22],导致毛蚶体内ATP含量下降。有研究报道,虾夷扇贝(Patinopecten yessoensis)在4℃干露下24 h内ATP含量变化不大,而48 h后含量急剧下降[28],也可能与此有关。
SOD是抗氧化系统的关键酶[29]。本实验中,毛蚶组织SOD活力在干露后6~24 h出现下降,表明干露胁迫对毛蚶的免疫系统产生了显著影响,可能产生的大量活性氧自由基抑制了抗氧化酶的活力。菲律宾蛤仔(Ruditapes philippinarum)在25℃下干露,SOD活力也出现显著下降[14],而虾夷扇贝各组织SOD活力在4℃干露1 d后显著上升[28,30],说明不同贝类在应对干露胁迫时的免疫反应有差异。ACP是溶酶体的重要组成部分,在体内直接参与磷酸基团的转移和代谢,并在免疫反应中发挥重要作用[31]。在本实验中,干露2 h时ACP活力变化不大,6~12 h活力显著上升,表明干露胁迫激活了毛蚶体内参与免疫的水解酶活力,以增加机体对外源异物的防御能力和代谢水平。有研究指出,干露条件下甲壳类无法获得足够的氧气,体内以无氧呼吸为主,会产生乳酸、琥珀酸等无氧代谢产物,导致甲壳类酸中毒[11-12],破坏其免疫功能[4,32-34],说明干露胁迫下甲壳类的呼吸代谢与免疫功能密切相关。
以上结果表明,干露胁迫初期,毛蚶稚贝主要通过有氧呼吸为机体提供能量;随着干露时间延长,稚贝逐渐转变为无氧代谢。转录组学分析发现,长牡蛎(Crassastrea gigas)在干露胁迫后维持基础代谢的能量成本增加,随着胁迫延续,长牡蛎通过减少体内代谢活动维持机体基础能量消耗,整体能量策略从积极的补偿策略变为保守的节能策略[27],本实验中毛蚶稚贝的代谢适应策略应与长牡蛎类似。
3.3 胁迫后恢复状况
呼吸代谢是直接反映生物体能量代谢的基本生理活动,不仅反映生物体的代谢特征和生理状态,还可用来评估环境改变对生物体的影响程度[13]。贝类滤水率受生物因素和环境因素的影响较大[35-37],是反映贝类生理状态的重要指标。耗氧率是新陈代谢的重要指标之一,也是贝类能量学研究的重要参数。环境条件不适合时,贝类通过调整自身代谢,消耗自身储存的能量来抵抗逆境,从而导致耗氧率的变化[38]。本实验中,干露毛蚶的滤水率在放回水中后,前12 h与对照组无显著差异,说明干露胁迫后毛蚶摄食能力的恢复需要一定时间,可能与毛蚶贝壳的开放程度有关。贝类在遇到不适环境时会通过关闭进出水管或贝壳来保护自己[39],进出水管上侧纤毛也会减少摆动[40-41],进出贝类体内的氧气量和水量会降低,导致耗氧率和滤水率下降。12 h后毛蚶的滤水率显著升高,说明毛蚶适应了水环境,贝壳张开,与外界水体交换量增大,这与对文蛤[16]的研究结果一致。本研究中,干露胁迫后放回水中的毛蚶耗氧率先降低后升高。贝类在经过胁迫后会出现生长抑制现象,经过一段时间的适应后,会进行代谢补偿[15],如栉江珧(Atrina pectinata)耗氧率会随着温度的升高先降低后升高[37]。本实验中,毛蚶放回水中24~48 h,耗氧率与对照组无显著差异,说明毛蚶经过48 h的干露后呼吸代谢仍然可以恢复到正常水平。
4 小结
本研究表明,干露胁迫对毛蚶稚贝的生理代谢有显著影响。干露胁迫初期,毛蚶稚贝通过提高有氧代谢水平维持基础代谢;随着干露时间延长,稚贝逐渐转为无氧代谢,以降低机体能量需求,维持机体生存需要。重新接触海水后,毛蚶稚贝通过调节代谢水平,如升高滤水率、降低耗氧率,来增加饵料摄入和机体能量水平,以尽快恢复到正常状态。本实验结果表明,毛蚶运输时干露时间应控制在24 h内,以降低无氧代谢对毛蚶机体的损伤。
猜你喜欢
淡水渔业(2023年2期)2023-03-29 04:03:30
海洋渔业(2022年6期)2022-02-11 03:33:08
养殖与饲料(2021年1期)2021-12-04 20:29:24
水产科学(2021年1期)2021-01-28 10:53:12
水产科学(2018年3期)2018-06-04 02:10:58
大连海洋大学学报(2017年3期)2017-06-19 18:56:11
河北渔业(2016年5期)2016-09-08 10:31:08
河北渔业(2015年6期)2015-06-24 11:19:13
应用海洋学学报(2014年1期)2014-11-22 07:17:50
浙江海洋大学学报(自然科学版)(2014年6期)2014-03-13 03:23:09
