
柑橘次级代谢产物β-蒎烯对柑橘溃疡病菌的生长抑制作用与机理初探
2022-07-01 15:30叶子怡范晓静邹华松
武夷科学 2022年1期
叶子怡, 范晓静, 卓 涛, 邹华松, 户 勋
(福建农林大学植物保护学院, 福建福州 350002)
柑橘溃疡病是严重的常发柑橘病害,主要危害叶、枝、果实,不仅会使柑橘树落果,还会导致柑橘树枯死(Bruningset al, 2010)。 目前,大多数种植区都是以化学防治为主,但是该防治方式使柑橘的食用安全风险增大,病原菌的抗药性增强。 因此,使用植物内生菌、壳聚糖等进行生物防治成为了近几年的研究热点(张勇祥, 2022)。
传统的松针保鲜法证明松针能有效控制柑橘果实的腐烂,尤其对柑橘青、绿霉病的防治效果显著(杨书珍等,2016)。 松针精油的主要成分β-蒎烯,对意大利青霉、白色念珠菌、大肠杆菌等表现出强烈的抑菌活性(张美红等,2018)。
本实验室在前期工作中发现,β-蒎烯作为柑橘自身就能产生的一种次级代谢产物,对柑橘溃疡病菌 (Xanthomonas citrisubsp.citri,Xcc) 的hrpG启动子有诱导作用。hrp基因(hypersensitive and pathogenicity)是Xcc编码蛋白以组成Ⅲ型分泌系统(type Ⅲ secretion system,T3SS)为害柑橘的关键致病基因。 在为害植物的过程中,Xcc 的Ⅲ型分泌系统所形成的针状结构将效应因子分泌到寄主细胞内(孙思等, 2017),导致病害发生。 β-蒎烯既可抑制Xcc 的生长繁殖,又能诱导Ⅲ型分泌系统相关基因的表达。 这是否是细菌在与植物互相对抗过程中的一个进化表现,本试验将进行初步探索。
1 材料与方法
1.1 试验材料
本试验所用菌株为柑橘黄单胞菌柑橘亚种Xcc 29-1,菌株由本实验室保存。
NB 培养基:5 g·L-1多聚蛋白胨、10 g·L-1蔗糖、2 g·L-1酵母提取物、3 g·L-1NaCl、6 g·L-1牛肉浸膏,去离子水配制,调pH 至7.0~7.2,另加琼脂粉15 g·L-1为固体培养基。 121 ℃高压蒸汽灭菌20 min 后于室温储存。
M63 培 养 基:100 mmol·L-1KH2PO4、 75 mmol·L-1KOH、 15 mmol·L-1(NH4)2SO4、1 mmol·L-1MgSO4、3.9 μmol·L-1FeSO4、22 mmol·L-1葡萄糖。 用 10 mol·L-1NaOH 调节 pH 至7.0,加去离子水定容到1 L,分装后121 ℃灭菌20 min。 加琼脂粉15 g·L-1为固体培养基。
本试验所用壮观霉素(spectinomycin,Spe)终浓度为 20 μg·mL-1。质粒:pRG960,Sper抗性,本实验室保存。
接种试验所用植物为感病葡萄柚品种Duncan。
其他试剂:β-蒎烯(6,6-二甲基-2-亚甲基-二环[3.1.1]-庚烷)母液用乙醇配制成浓度为0.0、0.1、0.2、0.4、0.6、0.8 和 1.0 μg·mL-1的 溶 液, 低 温 避 光 保 存。 β-葡 萄 糖 苷 酸 酶(β-glucuronidase,GUS)的底物 5-溴-4-氯-3-吲哚葡萄糖苷(X-gluc)使用浓度为 20 mg·mL-1。
1.2 不同浓度β-蒎烯对Xcc 29-1 体外生长的影响
制作 OD600=0.03 的 Xcc 29-1 野生型菌悬液,分别加入最终浓度为 0.0、0.1、0.2、0.4、0.6、0.8和 1.0 μg·mL-1的 β-蒎烯,于 28 ℃摇床孵育,每 6 h 测一次 OD600值。 所有试验每个处理设3 个平行,试验重复3 次。
1.3 不同浓度β-蒎烯对Xcc 29-1 体内生长和柑橘溃疡病症状形成的影响
制作 OD600=0.3 的 Xcc 29-1 野生型菌悬液,分别加入最终浓度为 0.0、0.1、0.2 μg·mL-1的β-蒎烯,混匀后,接种于葡萄柚叶片上。 取其中接种2、4 和6 d 的叶片,梯度稀释组织混合液,涂抹平板,培养,计算单菌落数量。 同时,5 d 后观察余下的葡萄柚叶片,查看溃疡症状形成的快慢和严重程度。
1.4 筛选hrpG 启动子与GUS 报告基因融合表达的菌株
为了测试β-蒎烯对Xcc 29-1 致病基因hrpG的启动子表达的影响,本试验制作了hrpG启动子与GUS 报告基因融合表达的菌株。 以Xcc 29-1 菌株的基因组DNA 为模板,用引物扩增hrpG基因启动子序列。 正反向引物序列为:5′-TCCTGCAGGATCATGCCGGTCTCTCT-3′和5′-TTCCCGGGTTAGGCGGCCTTCGCGTGT-3′。
PCR 扩增程序为:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 1 min,30 个循环;72 ℃ 10 min。PstⅠ-XmaⅠ双酶切后回收的产物连接到pRG960 载体上,构建的hrpG启动子-GUS 融合载体pRG960:PhrpG电转进入Xcc 29-1,在NB+Spe 平板上筛选获得菌株Xcc 29-1/pRG960:PhrpG。
1.5 hrpG 启动子活性检测
为了测试报告系统,将Xcc 29-1/pRG960:PhrpG分别接种于已涂抹X-gluc 的NB 和M63固体培养基上,用Xcc 29-1 和Xcc 29-1/pRG960 做对照。
1.6 不同浓度β-蒎烯对hrpG 启动子的体外诱导
在 OD600=0.3 的 Xcc 29-1/pRG960:PhrpG菌悬液中,分别加入最终浓度为 0.0、0.1、0.2、0.4、0.6、0.8 和 1.0 μg·mL-1的 β-蒎烯,于 28 ℃摇床孵育,6 h 后加入 20 μg·mL-1X-gluc,观察显色结果。
1.7 不同浓度β-蒎烯对hrp 基因的体外和体内诱导
OD600=0.3 的 Xcc 29-1 野生型菌悬液,分别加入最终浓度为 0.0、0.1、0.2 μg·mL-1的 β-蒎烯,于28 ℃摇床孵育,3 d 后收集菌液,提取 RNA,反转录,荧光定量 PCR 检测hrpG、hrpX、hrcV、hrpD6 基因的表达。
OD600=0.3 的 Xcc 29-1 野生型菌悬液,分别加入最终浓度为 0.0、0.1、0.2 μg·mL-1的 β-蒎烯,混匀后,接种于葡萄柚叶片上,3 d 后收集叶片,提取RNA,反转录,荧光定量PCR 检测hrpG、hrpX、hrcV和hrpD6 基因的表达。
2 结果与分析
2.1 不同浓度β-蒎烯对Xcc 29-1 体外生长的影响
如图 1(a)所示,β-蒎烯浓度为 0.0 μg·mL-1时,Xcc 29-1 在 48 h 的 OD 值达到 3.0。 随着β-蒎烯浓度的提高,当 β-蒎烯浓度为 0.2 μg·mL-1时,Xcc 29-1 的生长速度明显受到抑制。β-蒎烯最终浓度为 1.0 μg·mL-1时,Xcc 29-1 甚至不生长。 48 h 后的 Xcc 29-1 的生长情况如图1(b)所示,从左往右,β-蒎烯浓度逐渐提高,试管内培养基的浑浊程度逐渐下降。 β-蒎烯最终浓度为1.0 μg·mL-1的试管中培养基澄清,菌物几乎不生长。
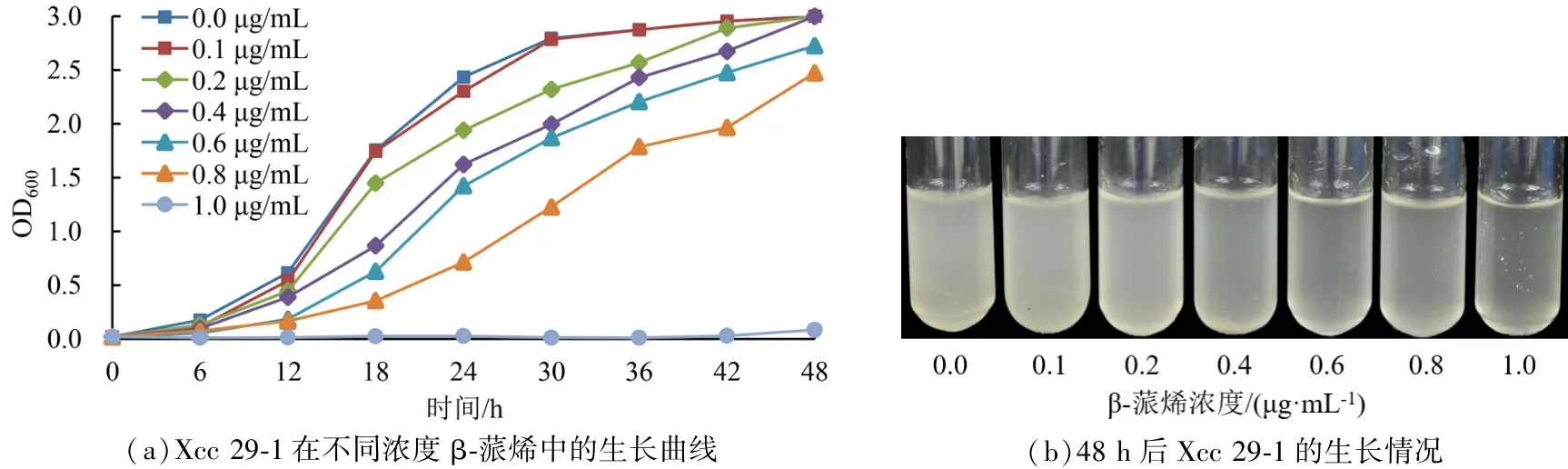
图1 液体培养基中不同浓度β-蒎烯对Xcc 29-1 生长的影响Figure 1 Effect of different concentrations of β-pinene on the growth of Xcc 29-1 in NB liquid medium
2.2 不同浓度β-蒎烯对Xcc 29-1 体内生长和柑橘溃疡病症状形成的影响
图2(a)为平板计数单菌落结果,加入β-蒎烯后,单位面积含有的细菌数量减少。 图2(b)为接种5 d 后的葡萄柚叶片,加入β-蒎烯的接种区域溃疡症状较轻。 0.2 μg·mL-1β-蒎烯的接种部位,能观察到部分坏死,这说明,0.2 ug·mL-1的β-蒎烯能快速诱导植物的抗病反应。 溃疡表型与Xcc 29-1 在葡萄柚叶片组织中的复制一致。
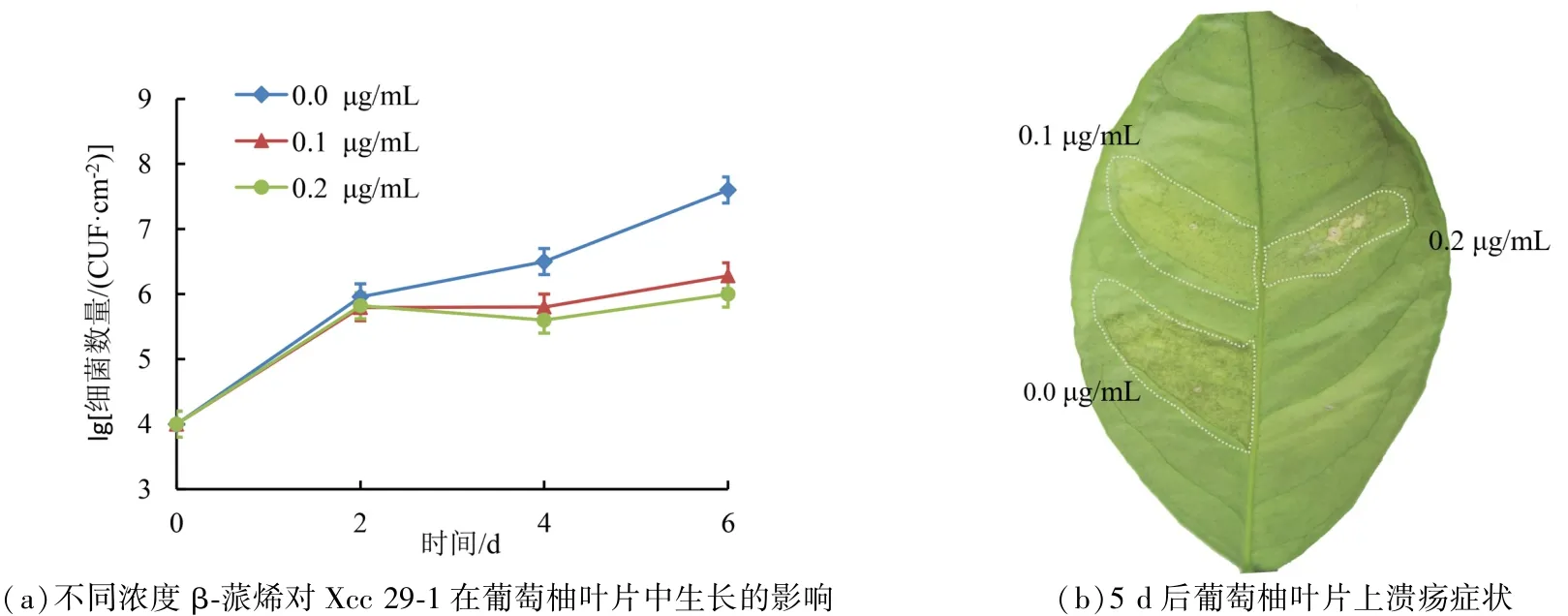
图2 植物体内不同浓度β-蒎烯对Xcc 29-1 生长的影响和溃疡症状形成情况Figure 2 Effect of different concentrations of β-pinene on the growth of Xcc 29-1 and formation of canker symptom in leaves
2.3 hrpG 启动子与GUS 的融合基因在Xcc 29-1 中的表达
图3 (a)为PCR 扩增的hrpG启动子片段的凝胶电泳结果,图3(b)为融合GUS 报告基因的菌株Xcc 29-1/pRG960:PhrpG在NA 和M63 固体培养基上的平板显色结果,培养基中加入了 20 μg·mL-1X-gluc。 对照组的 Xcc 29-1 和 Xcc 29-1/pRG960 均无显色表现,Xcc 29-1/pRG960:PhrpG在NA 和M63 固体培养基上均能显色,M63 更深,表明启动子在贫瘠培养基上更活跃。

图3 hrpG 启动子报道检测系统构建Figure 3 The construction of the hrpG promoter GUS fusion system
2.4 不同浓度β-蒎烯对hrpG 启动子的体外诱导
如图4,Xcc 29-1 加入不同浓度β-蒎烯6 h 后,随着浓度的提高,GUS 显色情况越明显,说明高浓度的β-蒎烯能高水平诱导hrpG启动子表达。 0.1 和0.2 μg·mL-1浓度也可以诱导,但水平较低。

图4 培养基中不同浓度β-蒎烯对hrpG 启动子的诱导Figure 4 The expression of the hrpG promoter in different concentrations of β-pinene in NB liquid medium
2.5 不同浓度β-蒎烯对hrp 基因的体外和体内诱导
图5(a)为体外试验结果,对照空白组,0.1 μg·mL-1β-蒎烯能诱导4 个代表性hrp基因的表达,其中hrcV的表达增强了 25 倍。 0.2 μg·mL-1β-蒎烯诱导 4 个hrp基因表达的增强都超过了4 倍。 图5(b)为体内试验结果,发现混合β-蒎烯接种后,hrp基因的表达都呈增强趋势。
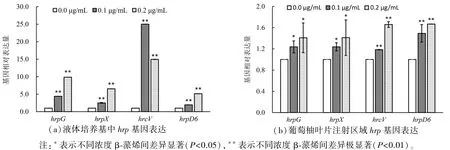
图5 不同浓度β-蒎烯对hrp 基因的诱导Figure 5 The hrp genes expression in different concentrations of β-pinene
3 结论与讨论
β-蒎烯作为柑橘的次级代谢化合物,其合成调控机制尚不清楚,但是在柑橘不同品种的精油成分中占有相当大的比例,对微生物具有直接的抑菌活性,推测可以抵御外来病原物的侵染(Tsaiet al,2017;Adokohet al,2019)。
植物合成的次级代谢产物对其与病原菌的互作有重要作用,其中包含相当一部分的植保素物质,对病原菌有直接的抑制效果,还有一部分次级代谢产物诱导寄主植物自身的抗病信号通路,增强对病原物侵染的抗性(胡洪涛等,2004)。 次级代谢物抑制病原菌生长的主要原因有:对细菌、真菌、寄生性种子植物等具有直接的杀灭效果,成为这些病原生物的拮抗物质;协助金属离子运输;刺激菌物孢子的产生,影响胞子萌发,在逆境中使芽孢暂时休眠,暂缓生长,并保护休眠中的孢子;维护生长发育环境(Demainet al,2000;廖圣良等,2016;尚春雨,2017)。
柑橘溃疡症状的形成需要 Xcc 中hrp基因的表达(Guoet al,2011;Songet al,2015),而且这种表达需要寄主植物的诱导,即与柑橘植物互作后,hrp基因才能被高水平诱导表达,这与其它植物病原黄单胞菌的hrp基因表达一致(Alfanoet al,1997;杨军等,2005)。
在前人研究的基础上,本研究发现β-蒎烯作为次级代谢产物是柑橘抑制Xcc、阻止病害发生的一个化合物,同时又可以被病原物Xcc 所利用,作为植物来源的一个信号分子,诱导hrp基因的表达。 这为更全面认识β-蒎烯在柑橘与Xcc 互作中功能提供了很好的数据支撑。 同时,还发现β-蒎烯对Xcc 的生长的抑制需要达到一定的浓度阈值。 因此,可以推测,β-蒎烯合成的差异可能也是品种抗性水平变化的一个因素。
猜你喜欢
草地学报(2022年3期)2022-03-28
今日农业(2021年11期)2021-11-27
中国食用菌(2020年11期)2021-01-18
自我保健(2020年2期)2020-12-04
幼儿教育·父母孩子版(2020年6期)2020-07-27
现代园艺(2020年3期)2020-03-05
数位时尚(幼儿教育)(2018年10期)2018-10-30
世界热带农业信息(2016年4期)2016-05-03
世界热带农业信息(2016年2期)2016-03-11
食品工业科技(2014年23期)2014-03-11
