
种子散布位置对格氏栲幼苗根系形态及生长特征的影响
2022-06-28 03:11:40刘金福王雪琳何中声施友文沈彩霞
生态学报 2022年10期
朱 静,刘金福,邢 聪,江 蓝,王雪琳,何中声,*,施友文,沈彩霞
1 福建农林大学林学院,福州 350002 2 福建农林大学海峡自然保护区研究中心,福州 350002 3 生态与资源统计福建省高校重点实验室,福州 350002 4 福建省林业调查规划院,福州 350003 5 格氏栲自然保护区管理站,三明 365000
森林天然更新过程中,种子(种子雨和种子库)与幼苗(定居)阶段是森林更新的决定性环节[1]。种子从形成种子雨到种子库中,靠自身重力及外力作用从植物母株上掉落,凋落物是最初接触的物理环境。受凋落物厚度或所处凋落物位置影响,面临着不同的散布结果[2],(1)种子雨输入时,受林下地被层或地表凋落物的机械阻碍滞留于凋落物表层[3];(2)形成土壤种子库过程中,种子可能被动物搬运埋藏在土壤表层;(3)散落于枯枝落叶或土壤表层的种子受外界干扰,被凋落物所覆盖[4—5],其厚度变化显著影响幼苗出苗。不同种子散布位置影响着种子萌发及根系生长,且凋落物覆盖方式和厚度在其中扮演着重要角色,并可能对种子萌发和生长产生物理阻隔或保护作用[6],在一定程度上决定森林树种组成和森林更新。
根系作为植物吸收水分及养分的主要器官,其形态构型及生长状况反映了植物在逆境胁迫中的根系形态稳定性及资源利用效率[7]。种子从母株掉落于凋落物上层,林下地被层或地表凋落物产生物理阻隔作用,表层种子萌发后根系无法进入土壤吸收充足的水分及养分导致枯萎或死亡[8]。凋落物中下层分解速率快,养分资源充足且保水能力强,凋落物上层种子萌生根系一旦扎根于凋落物向下生长,根系从凋落物层获取养分效率高于土壤层,导致凋落物层中根产量显著提高[7]。凋落物表层的种子萌生根系难以进入土壤中导致死亡,而一旦进入凋落物层促进了根系性状的表达。
凋落物表层部分种子被动物搬运或重力作用下散布于土壤表层,种子萌发进入土壤容易,而萌发后根系生长过程受较强太阳照射,土壤表面易形成干旱的环境条件导致失水或新生根系被灼伤,一定程度上对根系生长起抑制作用[9]。土壤表层空气日温差变化幅度大,影响根系吸水能力和酶活性等生理生化进程,而种子吸水能力降低导致体内贮存的养分无法转变成根系能够利用的可溶性状态,降低根系生长速率[10]。种子散布在土壤表层,表层水分及光照等条件的改变导致根系在生长过程中出现枯萎或死亡,抑制根系生长。
当种子雨后发生凋落物输入时,林下种子被凋落物所覆盖,适宜凋落物通过改善土壤理化性质,增加土壤孔隙度及渗水性能,降低土壤酸化程度等途径,为根系呼吸及养分吸收提供优越的环境条件,促进根系伸展及扩张[11—12]。种子上层覆盖的凋落物经分解释放大量矿质养分归还给土壤,调节和促进生态系统中养分固持和周转,为根系生长提供充足养分来源[13]。凋落物覆盖也为根系生长提供荫蔽条件,降低强光对根直接照射,一定程度上促进或改善幼苗根系生长[8]。然而,根系生长对上层凋落物厚度的承受能力有一定范围,当厚度较深时对幼苗向上生长产生较强的机械阻碍力,幼苗为穿过厚的凋落物层降低对根系投入,以减少消耗自身大量营养物质[14],抑制了幼苗根系生长。此外,凋落物消光作用下幼苗为满足自身对光照的需求,将营养物质分配给地上组织进行光合作用,减弱根系生长[11]。可见,根系在种子不同散布位置时通过调整自身形态特征以维持植物生长需求,而关于不同散布位置种子根系的响应机制未见报道,限制了对幼苗根系在不同散布位置适应策略的理解。
格氏栲(Castanopsiskawakamii),是中国中亚热带南缘特有的壳斗科常绿阔叶高大珍稀濒危乔木,幼苗主根发达[15]。受自然及人为干扰等原因,年龄结构呈“倒金字塔型”[16]。格氏栲林内凋落物层厚,种子从植物母株掉落于凋落物上层,林下枯枝落叶层与草本层等形成厚的隔离带,导致滞留于上层种子约2/3,种子萌发率显著降低[17];或种子萌发后,胚根无法与土壤接触造成枯萎或死亡;一旦根系与土壤接触,生长速度加快[6]。土壤表层的种子受光照和水分等资源限制,萌发速度慢;当水分或光照胁迫解除时,种子恢复萌发,萌发持续时间长,萌发率高于其它散布;而胚根在两种胁迫下,造成不可逆的枯萎或死亡,降低了幼苗定居机会[6]。凋落物覆盖降低种子萌发所需的光照,萌发受到抑制;而覆盖产生的荫庇环境促使胚根在全黑暗条件下生长加快[6]。综上所述,格氏栲林内落种更新的实生幼苗更新困难是否与种子不同散布位置的根系生长状况有关值得深入探究。目前,关于3种散布位置下格氏栲根系形态生长特征的研究尚未报道,限制了对格氏栲天然更新能力的深入了解。
因此,通过室内模拟3种种子散布位置,探讨不同散布对格氏栲幼苗根系形态及生长特征的影响,旨在解决:(1)不同散布位置下幼苗根系性状表达是否存在差异;(2)基于根系性状的综合分析,不同散布位置是抑制还是促进了根系生长,未来如何调控林下凋落物层厚度以促进森林更新?即通过研究不同种子散布位置下格氏栲幼苗根系形态及生长特征,探寻适宜根系生长的凋落物厚度,为格氏栲林天然更新及可持续发展提供理论依据。
1 材料和方法
1.1 研究区概况
三明格氏栲自然保护区位于福建省三明市莘口镇,地处26°07′—26°12′ N、117°24′—117°29′ E,面积近700 hm2,海拔180—640 m。气候属中亚热带季风性气候,年平均温度19.5℃,年平均降雨量为1500 mm,年平均相对湿度为79%。土壤为酸性铁铝土,凋落物层较厚,土层腐殖质丰富[18]。保护区植物种类丰富,乔木层主要树种有格氏栲、桂北木姜子(Litseasubcoriacea)、木荷(Schimasuperba)、马尾松(Pinusmassoniana)等,灌木层主要有狗骨柴(Diplosporadubia)、山黄皮(Clausenadentata)和杜茎山(Measajaponica)等;草本植物主要有狗脊蕨(Woodwardiajaponica)、华山姜(Alpiniajaponica)和芒萁(Dicranopterisdichotoma)等[19]。
1.2 供试材料收集
2018年3月份于格氏栲自然保护区设置3个大林窗(≥200 m2)、3个中林窗(50—100 m2)、3个小林窗(≤50 m2)和1个非林窗(400 m2)。为减少因凋落物类型造成的实验误差,在每个林窗和非林窗东、南、西、北和中心5个方位,距离地面1 m处各设置1个1 m×1 m的凋落物收集框,每月月初收集框内样品带回实验室混合,于65℃烘箱中烘干至恒重。在种子结实丰年的掉落高峰期,保护区内收集大小一致的格氏栲种子(约2 g)带回实验室洗净,置于蒸馏水中静置3 h,去除漂浮、劣质的种子,将沉于水下的饱满种子捞出备用[6]。
1.3 试验设计
室内试验于2018年12月在福建农林大学林学院实验大棚内开始,选用38.5 cm×27.5 cm×14 cm规格的萌发盆,依据萌发盆体积,计算相应加入的凋落物重量。格氏栲种子成熟从植物母株掉落于地面,受凋落物影响散布在凋落物上层、土壤表层及凋落物下层。依据3种散布结果,采用完全随机设计,设置种子底层铺垫凋落物处理(U)、土壤表层处理(CK)及种子上层覆盖凋落物处理(D)等3种试验方案。种子底层铺垫凋落物设置2个试验梯度,即先铺2(U2,约40 g/m2),4(U4,约80 g/m2)cm厚凋落物再播种;种子在土壤表层及上层覆盖凋落物处理设置种子与土壤有机质直接接触,上层覆盖0(CK),2(D2,约40 g/m2),4(D4,约80 g/m2),6(D6,约120 g/m2),8(D8,约160 g/m2)cm厚凋落物[5]等5个试验梯度。试验共设计7个凋落物厚度梯度,每个处理设置3个重复,每个重复采用均匀撒播方式播种50粒颗粒饱满、大小基本一致的格氏栲种子(约2 g)。每周轮流调换萌发盆位置,以保证生长期间受光条件基本一致[14],每隔2天每次浇水200 mL。土壤选择、种子处理等具体试验方法参考文献[6]。
1.4 样品采集
以幼苗露出两片真叶为出苗标准[20],待出苗试验结束统计出苗率(16%—54.7%);继续培养幼苗6个月收获幼苗,每个重复选取3株地上长势基本一致,无病虫害的格氏栲幼苗。取样时,先将地上部分及根系用剪刀分开,小心抖落土壤,避免伤害根系组织及构型。在实验大棚内快速分为根、茎和叶,用清水清洗各器官附着的泥土等杂物,并尽量保证根系完整性;将上述各器官做好标记放入自封袋,置于事先准备好的冰盒,迅速带回实验室。使用低温去离子水将根系清洗干净,用滤纸吸干表面水分,置于4℃冰箱保存。3株幼苗测量数据的平均值作为一次有效值。
1.5 形态生长指标测定
1.5.1根系指标测定
将置于4℃冰箱内根系采用EPSON根系扫描仪(EPSON Expression ll000XL, SEIKO EPSON CORP., Japan)获取根系形态图像,利用根系图像系统分析软件(Win-RHIZO Pro 2007, Regent Instruments Inc., Quebec, Canada)扫描和定量分析根系形态图像,记录总根长(RL,root length)、根表面积(SA,root surface area)、根系体积(RV,root volume)、根系平均直径(RAD,root average diameter)、根尖数(RTN,root tips number)、分枝数(RFN,root forks number)等指标。将扫描后的根系装入信封,做好标记,置于80℃烘箱内烘干至恒重(48 h),利用电子天平(精度0.0001 g)称根系干物质质量(RDM,root dry mass)。
1.5.2比根长及比根表面积测定
比根长(SRL,cm/g)=RL/RDM;
比表面积(SRA,cm2/g)=SA/RDM[21];
1.6 数据统计
采用Excel绘制图表;利用单因素方差分析种子不同散布位置对格氏栲幼苗根干物质质量、根长、根系平均直径、根表面积、根体积、根尖数、分枝数、比根长及比表面积的影响,以新复极差法(Ducan)进行同组内差异比较(α=0.05)。利用皮尔逊(Pearson)相关分析9个幼苗根系指标的相关关系。利用主成分分析方法(Principal Component Analysis)对9个根系指标提取主成分后,按其亲疏相似程度将7个厚度梯度进行层次聚类分析(Hierarchical Clustering Analysis)。数据分析均在SPSS 21.0及R 3.6.3中实现。
2 结果分析
2.1 种子散布位置与幼苗根干物质质量

图1 不同散布位置对幼苗根干物质质量的影响 Fig.1 Effects of different dispersal positions on root dry mass of seedlingsU2:种子底层铺垫2 cm凋落物;U4:种子底层铺垫4 cm凋落物;CK:种子上层无凋落物覆盖;D2:种子上层覆盖2 cm凋落物;D4:种子上层覆盖4 cm凋落物;D6:种子上层覆盖6 cm凋落物;D8:种子上层覆盖8 cm凋落物;图中不同小写字母表示不同处理间差异显著(P<0.05)
格氏栲幼苗根干物质质量在同一凋落物厚度下(图1),凋落物上层(U处理)根干物质质量低于凋落物覆盖(D处理);除土壤表层(CK处理)外,根干物质质量随凋落物覆盖厚度增加呈下降趋势,凋落物浅层覆盖(D2处理)对幼苗根干物质质量累积的促进作用最为显著,凋落物深层覆盖(D6及D8处理)及CK处理起抑制作用。
2.2 种子散布位置与幼苗总根长及根系平均直径
种子不同散布下总根长表现为同一凋落物厚度下,凋落物上层总根长低于凋落物覆盖(图2)。与土壤表层相比,凋落物覆盖的总根长随厚度增加呈先下降后上升趋势,D2处理显著高于其它处理(P<0.05),凋落物深层覆盖(D6及D8处理)对总根长起抑制作用。
格氏栲幼苗根系在同一凋落物厚度下,散布在凋落物上层的幼苗根系平均直径高于凋落物覆盖。凋落物覆盖下根系平均直径随覆盖厚度增加呈先升后降趋势,D6处理达最大值,D2处理抑制根系平均直径加粗。与土壤表层相比,各处理均与其无显著差异性(P>0.05)。

图2 不同散布位置对幼苗根长及根系平均直径的影响Fig.2 Effects of different dispersal positions on root length and root average diameter of seedling
2.3 种子散布位置与幼苗根表面积及根体积
不同种子散布位置的根表面积表现为(图3),同一凋落物厚度下,凋落物上层根表面积低于凋落物覆盖。与土壤表层相比,凋落物覆盖下根表面积随覆盖厚度增加呈下降趋势,D2处理的促进作用最为显著。与其它散布位置相比,土壤表层及凋落物深层覆盖(D6及D8处理)均对根表面积起抑制作用,且三者间无显著差异性(P>0.05)。
格氏栲幼苗根体积在3种散布位置的变化为同一凋落物厚度下,凋落物上层根体积高于凋落物覆盖。凋落物覆盖的幼苗根体积随覆盖厚度增加而下降,D2处理达最大值。与土壤表层相比,除D8处理外,凋落物上层与凋落物覆盖两种散布位置均促进幼苗根系伸展及扩张,提高幼苗根体积。

图3 不同散布位置对幼苗根表面积及根体积的影响Fig.3 Effects of different dispersal positions on root surface area and root volume of seedling
2.4 种子散布位置与幼苗根尖数及分枝数
3种不同种子散布位置对格氏栲幼苗根尖数的影响表现为(图4),同一凋落物厚度下,凋落物上层根尖数高于凋落物覆盖。与土壤表层相比,凋落物覆盖的幼苗根尖数随厚度增加而下降,D2处理起显著促进作用(P<0.05);凋落物深层覆盖(D6及D8处理)与土壤表层均抑制根尖数,且三者间无显著差异性(P>0.05)。
格氏栲幼苗根分枝数在同一凋落物厚度下,凋落物上层分枝数低于凋落物覆盖。凋落物覆盖下根分枝数随厚度增加呈先降后升趋势,D2处理显著高于其它处理,其它各处理间无显著差异性(P>0.05)。综上,凋落物浅层覆盖(D2处理)对根分枝数的促进作用最为显著。

图4 不同散布位置对幼苗根尖数及分枝数的影响Fig.4 Effects of different dispersal positions on root tips number and root forks number of seedling
2.5 种子散布位置与幼苗比根长及比表面积
凋落物上层各处理幼苗根系比根长间无显著差异性(图5)。凋落物覆盖下,D2处理起显著促进作用,幼苗比根长显著高于其它处理(P<0.05)。深层凋落物覆盖及土壤表层间幼苗比根长无显著差异性(P>0.05),均降低了幼苗比根长。
种子散布位置对幼苗根系比表面积的影响为同一凋落物厚度下,凋落物上层比表面积高于凋落物覆盖。D2处理的幼苗根系比表面积高于其它覆盖厚度。与凋落物深层覆盖及土壤表层相比,凋落物上层(U处理)及凋落物浅层覆盖(D2处理)提高了幼苗根系比表面积(P<0.05)。
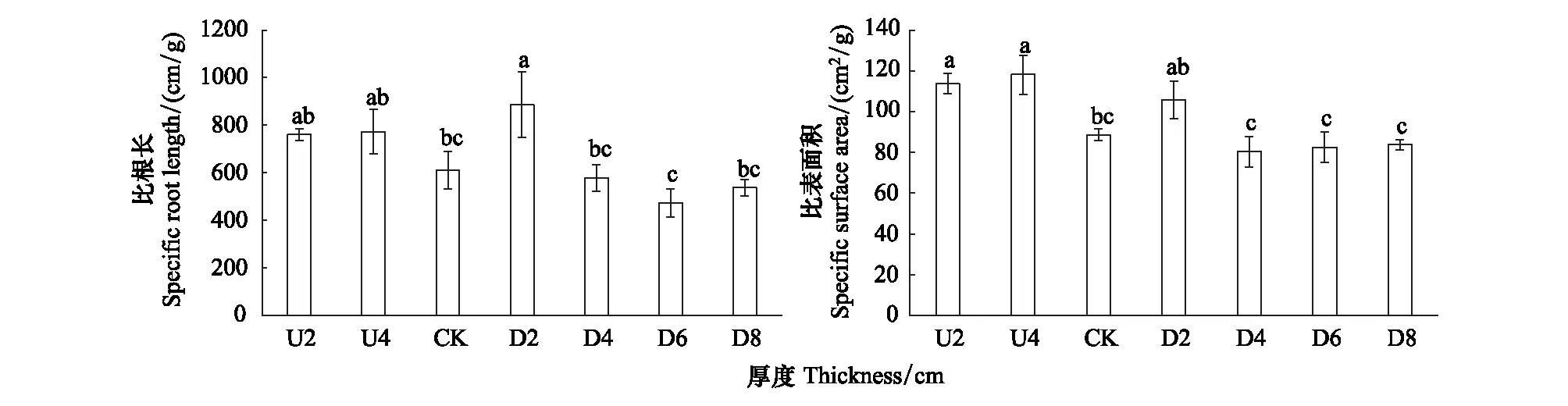
图5 不同散布位置对幼苗比根长及比表面积的影响Fig.5 Effects of different dispersal positions on specific root length and specific surface area of seedling
2.6 格氏栲幼苗根系性状主成分及相关性分析
利用主成分分析方法对3种不同散布位置下幼苗9个根系生长指标提取主成分(表1),前3个主成分累计贡献率达99.637%。第1主成分与总根长、根表面积、根尖数及分枝数呈高度正相关,提取主成分达92%以上,总根长对第1主成分贡献率最高。第2主成分与根干物质质量呈正相关,与比表面积呈负相关。根体积对第3主成分贡献率最大,呈高度正相关。

表1 根系生长指标的主成分分析
3种种子散布位置下格氏栲幼苗根系生长指标间存在显著相关关系(图6)。根干物质质量与总根长、根表面积、根体积、根尖数和分枝数呈极显著正相关。根长与其它生长指标间均存在显著相关性。根系平均直径与根长、根表面积、根尖数、分枝数及比根长呈显著负相关关系。根表面积与根体积、根尖数、分枝数、比根长及比表面积具有正相关关系。

图6 格氏栲幼苗根系特性的相关性分析Fig.6 Correlation analysis of root characteristics of C. kawakamii seedlingsRDM:根干物质质量 root dry mass;RL:根长 root length;RAD:根系平均直径 root average diameter;SA:根表面积 root surface area;RV:根体积 root volume;RTN:根尖数 root tips number;RFN:分枝数 root forks number;SRL:比根长 specific root length;SRA:比表面积 specific surface area;*:显著相关,P<0.05;**:极显著相关,P<0.01;***:极显著相关,P<0.001;红色圆圈表示正相关,蓝色表示负相关
2.7 格氏栲幼苗根系指标的聚类分析

图7 根系生长指标聚类分析Fig.7 Cluster analysis of root growth indices
以欧氏距离作为衡量3种散布位置下根系生长指标的差异大小,采用最短距离法将7个凋落物厚度梯度按9个根系指标的亲疏相似程度进行聚类分析,相似性最大的优先聚合在一起(图7)。9个指标中根长、根表面积、根尖数及分枝数提取主成分量高于92%(表1),将其聚类后7个梯度划分为4个类群。凋落物浅层覆盖(D2处理)的根长、根表面积、根尖数及分枝数显著高于其它散布,单独划分一类;种子在凋落物上层(U处理)对根系生长的促进作用次于D2处理,且9个根系指标中两者相似性最大。D4处理的幼苗根长等指标处于D2处理与凋落物深层覆盖(D6及D8处理)的过渡阶段,单独划分一类。凋落物深层覆盖和土壤表层(CK)根长、根表面积、根尖数、分枝数指标较低,且处理间无显著差异性,划分为同一类。综上所述,聚类分析与主成分分析评价结果较一致,表明了不同散布位置下根系在7个凋落物厚度梯度的生长状况。
3 讨论
3.1 种子不同散布位置对格氏栲幼苗根干物质质量的影响
植物干物质质量累积受光照、水分及养分等资源限制,并通过分配维持自身功能平衡[14]。种子不同散布位置显著影响格氏栲幼苗根系干物质质量累积,同一凋落物厚度下,凋落物覆盖的根干物质质量高于凋落物上层(U处理)及土壤表层(CK处理),在于凋落物覆盖影响了土壤水分和光照条件,而促进根干物质质量累积。凋落物上层或土壤表层散布的种子萌发后,表层水分蒸发速率快,幼苗光合作用弱,同化产物合成减少[22],同一凋落物厚度下,萌生根系因缺水失去活力,导致根干物质质量累积降低。凋落物覆盖可减缓水分散失,维持根系生长所需的水分,提高了幼苗存活率和根干物质质量。Montgomery[23]研究表明光照决定植物干物质质量累积,光照不足时植物为了获取更高的光合效率,常采取增大获取性资源的器官分配,减少非限制性获取资源器官分配的策略以提高光能利用率[24]。种子在凋落物上层及土壤表层的根系暴露于空气中,直接接收太阳照射,新生根系可能被强光灼伤而降低根系干物质质量累积。覆盖适宜凋落物降低了光照对根系直射,为根系生长提供遮荫条件,促进根系快速生长。然而,覆盖较厚凋落物降低了幼苗生长所需可用光,幼苗为了快速出苗并恢复正常光合作用,低光环境下采取缓慢的资源获取和消耗策略来维持正常生长[25],从而降低根干物质质量。凋落物深层覆盖(D6及D8 处理)的根干物质质量降低证实了深荫环境使到达根系的光强和光质减弱,幼苗通过减弱根干物质质量分配来适应弱光环境,以增强植物对光的竞争能力,满足生长[26]。3种种子散布位置表明,适宜凋落物覆盖既可改善土壤理化性质,又为根系生长提供遮荫条件,保证了幼苗生长初期对光照和水分需求,一定程度上促进根干物质质量累积[27]。
3.2 种子不同散布位置对格氏栲幼苗根系形态指标的影响
根系是幼苗与土壤接触媒介,其生长状况直接影响幼苗对水分和养分资源的吸收能力[28—29]。种子不同散布位置的萌生根形态特征表明,同一凋落物厚度下,凋落物覆盖的种子萌生根根长、根表面积、根尖数及分枝数高于凋落物上层,在于种子下层凋落物过厚或密度太大致使根系无法突破凋落物的机械阻碍而抑制生长[13];而上层根系一旦扎根于凋落物,从凋落物层汲取充足养分及水分资源,根系生长速度快且优于土壤表层,与朱静等[6]结果一致。根系形态可塑性调整也可能在于凋落物对土壤理化性质的影响。Boonman等[30]研究表明根系生长的养分通量主要发生在土壤表面,通过凋落物循环利用改变土壤养分含量,影响幼苗生长速度。种子在凋落物上层或土壤表层其体内吸水速率低于蒸腾速率,导致根系协调自身生长能力遭到破坏[31],幼苗常降低根长、根表面积、根尖数及分枝数来减少自身能量的消耗。土壤表层较低水分也会导致氧气浓度升高,加快微生物对土壤有机物质矿化,降低土壤中有机质数量[32];同时减少了地表微生物可利用碳源,降低土壤呼吸速率,对根系生长起显著抑制作用[33]。覆盖适宜凋落物对土壤表层起到保温及保湿作用,增强根系对高温和干旱的抵抗能力;当根系逐渐向深层土壤生长时,适宜的水分将土壤中营养物质转化成根系可利用状态,增加幼苗对养分资源的吸收而促进生长[34]。适宜凋落物覆盖通过植物体内的糖、淀粉等非结构性碳刺激微生物群落扩张,加速土壤有机物质分解[32],增加有机质与土壤微生物可利用碳源,提高土壤呼吸速率[33]等促进根系生长。凋落物存在(D2、D4及U2处理)的根长高于土壤表层,也证实了适宜凋落物覆盖经分解释放大量矿物质养分归还土壤[7],增加了凋落物层淋溶的可溶性有机质浓度,为根系生长提供充足养分来源。然而,根系生长对凋落物的承受能力具有一定范围,当超出可承受范围后,幼苗无法突破凋落物产生的机械阻碍以维持自身向上生长[14]。深层凋落物覆盖起抑制作用,在于深层凋落物覆盖降低了幼苗生长所需光照,植物光合作用弱,幼苗为了减少根部碳和其它能量消耗,将更多资源分配给地上部分以提高幼苗对弱光环境的耐受力和生存适合度[35]。凋落物深层覆盖减少了表层土壤日温差,阻碍了根系与外界环境的气体交换[36],降低了土壤通气性,导致根系泌氧较少,根系细胞呼吸代谢方式从有氧呼吸转变为无氧呼吸[37],造成根系活力下降而出现枯萎或死亡。D6及D8处理的幼苗根系生长速度缓慢也证实了凋落物深层覆盖降低了幼苗根系活力及资源吸收能力,导致根系发育受阻,初生根受损死亡,幼苗根系生长缓慢。
根系平均直径是反映植物根系形态特征的重要参数,影响植物的吸收和运输功能[38]。研究表明同一凋落物厚度下,种子在凋落物上层提高了幼苗根系平均直径,可能是种子散布在凋落物上层或土壤表层的萌生根系接收光持续时间长,表层水分蒸发迅速,在水分亏损情况下,幼苗从根部调取养分来维持地上组织的水分需求,同时储存淀粉以供再生[30];当水分胁迫解除后,根系快速扩张吸收水分,减少对根系平均直径的消耗,导致凋落物上层及土壤表层的根系平均直径较高。格氏栲属于外生菌根树种[39],通过外延菌丝扩张增加根系吸收面积,以保证植物高效获取土壤养分和水分资源[40]。深层凋落物覆盖下,幼苗为了抵御较厚凋落物覆盖带来的遮荫效果,通过增加根系平均直径,提高根系对资源的运输能力,以保证地上光合作用产生的养分能够用于植物生长。D6和D8处理的根系平均直径高于其它覆盖厚度,也证实了幼苗为弥补自身向上生长所消耗的营养物质,通过加粗根系平均直径储藏碳水化合物,为幼苗在短期胁迫环境下提供养分[41]。凋落物浅层覆盖(D2处理)通过将根系变得纤细来扩展整体的吸收和运输范围,提高根系对土壤水分及养分资源的获取能力,降低了根系平均直径。可见,3种散布位置下格氏栲幼苗通过根系形态可塑性调整及生态适应性策略,增加对光照、水分及养分资源的吸收与利用,从而提高幼苗建成与定居的机会。
3.3 格氏栲种群更新的启示
格氏栲属于耐荫性物种,主根发达,幼苗生长初期需要相对郁闭的环境条件。聚类分析将7个厚度梯度划分为4个类群(图7),且类群之间存在差异性。种子在凋落物上层,其萌生根根系受凋落物物理阻隔作用影响,生长速度较慢;上层根系一旦与土壤接触后,通过根系向四周扩张,大范围地吸收与利用土壤中水分与养分资源,以促进根系生长。土壤表层的根系暴露于空气中,幼根易因缺水或被强光灼伤失去活力,导致根系生长指标较低于其它覆盖厚度,降低了格氏栲幼苗建成几率。相关分析可得(图6),幼苗根干物质质量和根长等指标与根系平均直径具有显著负相关关系。凋落物浅层覆盖(D2处理)既保证植物的光照需求,也为其生长提供良好的遮荫效果,满足幼苗初期需耐荫而生长到一定阶段需足够光照的特性。格氏栲幼苗采取快速的资源占有策略,通过增加根长、根表面积、根尖数、分枝数和比根长等降低构建和维持根系单位长度所需的碳水化合物,扩大根系对土壤空间占有量,增强根系资源利用效率和养分循环效率以提高幼苗定居的机会。凋落物深层覆盖(D6和D8处理)产生较强的消光和物理阻隔效应,幼苗为获取更多有效光进行光合作用,倾向于增加根系平均直径来提高对资源的运输能力,降低根系生长对养分的消耗,以弥补短期胁迫带来的生长压力,从而提高格氏栲幼苗光竞争力和生存适合度。3种不同散布位置下,凋落物浅层覆盖(D2处理)对根系生长的促进作用最为显著,较厚凋落物覆盖起抑制作用。因此,在未来森林经营管理过程中,通过人为适当干扰,合理调控森林凋落物厚度,为根系提供良好的生长环境。综上,研究不同散布位置对格氏栲幼苗根系形态及生长特征的影响,不仅反映格氏栲幼苗在不同环境中的生长状况,也为格氏栲天然林恢复和发展提供科学依据。
4 结论
种子从植物母株掉落于地面,受凋落物厚度及其在凋落物中位置影响,格氏栲幼苗根系具有不同的生长特性。凋落物浅层覆盖(D2处理)通过提高幼苗根干物质质量、根长、根表面积、根尖数及分枝数,降低根系平均直径等促进植物高效获取土壤养分及水分资源,从而促进格氏栲幼苗快速生长;凋落物上层的促进作用次之;聚类分析将土壤表层与凋落物深层覆盖(D6及D8处理)划分同一类,通过加粗根系平均直径,降低根系生长指标等储藏营养物质以促进植物对养分资源的吸收及利用。综上所述,凋落物覆盖2 cm是最适宜格氏栲幼苗根系生长的覆盖厚度,其它散布位置下幼苗通过自身较强的形态可塑性及多元适应策略,弥补有限资源(光照及水分等)环境下的生长。将来,在种子掉落高峰期来临前或幼苗出苗阶段及时清理森林凋落物,或在土壤表层萌发种子上层覆盖2 cm凋落物,以促进格氏栲林天然再生。
猜你喜欢
发明与创新(2023年30期)2023-10-11 01:37:12
小学生学习指导(高年级)(2023年3期)2023-03-31 06:03:22
小学生学习指导(高年级)(2022年3期)2022-03-29 07:49:16
海洋石油(2021年3期)2021-11-05 07:43:10
小小说月刊·下半月(2021年8期)2021-09-05 05:49:31
河北环境工程学院学报(2021年1期)2021-03-19 08:43:00
动漫界·幼教365(大班)(2018年3期)2018-05-14 11:50:46
小学生导刊(高年级)(2017年2期)2017-06-10 02:40:42
山东农业科学(2017年4期)2017-05-18 17:06:13
焊接(2015年6期)2015-07-18 11:02:25
