
克隆植物对异质生境的适应对策研究进展
2022-06-28 03:12:34张晓龙吴梦迪吴秋堂王立冬张树岩韩广轩
生态学报 2022年10期
张晓龙,吴梦迪,,吴秋堂,,王立冬,张树岩,黎 磊,韩广轩,管 博,*
1 烟台大学,环境材料与工程学院, 烟台 264005 2 中国科学院海岸带环境过程与生态修复重点实验室(烟台海岸带研究所),山东省海岸带环境过程重点实验室,中国科学院烟台海岸带研究所, 烟台 264003 3 山东黄河三角洲国家级自然保护区管理委员会, 东营 257091 4 江西省流域生态演变与生物多样性重点实验室,南昌大学生命科学研究院流域生态学研究所, 南昌大学生命科学学院, 南昌 330031
生境异质性是自然生态系统的基本特征[1]。植物生长的必需资源(如光照、水分和矿质元素等)以及环境因素(如温度、湿度、盐分和病原体等)存在着复杂的时间和空间异质性[2—8]。在生态系统中,生境异质性不仅影响着生物群落中物种的多样性、群落的生长形式和结构、优势种、相对丰富度以及营养结构等,还影响着种群的生长与繁殖以及植物个体的生存适应。在不同尺度上,大到生态系统、群落,小到种群、个体,都存在生境异质性,而异质性生境在很小的时空尺度内就能对植物产生深远影响。然而,在复杂的生境条件下,植物也进化出了不同的生态适应对策[9]。克隆植物是指在自然条件下具有克隆特性的植物,即可通过与母株相连的芽、根茎、分蘖或枝条等繁殖体产生无性繁殖的植物,这些繁殖体一旦定居便可成为潜在的独立个体[10]。它能够通过其特有的生境适应对策(包括形态可塑性、克隆整合、克隆分工、觅食行为等)以最大限度地从异质性生境中获取资源和逃避胁迫,促进资源的高效利用,最终提高植物的适合度[11—12]。
克隆植物有许多不同的类型,不同的分类标准下植物类型也有所不同。根据克隆器官起源的不同,克隆植物可以分为3种类型:根源型克隆植物(root-derived clonal plant)、枝源型克隆植物(shoot-derived clonal plant),以及同时具有这两种克隆器官的克隆植物[13—14]。根源型克隆植物又可分为不同的亚类,如:根出条型、根劈裂型和块根型等[15]。枝源型克隆植物又可分为匍匐茎型、根状茎型、珠芽型、分蘖型、块茎型、球茎型、鳞茎型、压条型等以及其它亚类,包括以上类型的组合。根据克隆器官分枝类型的不同,可分为单轴型克隆植物(clonal plant with monopodial pattern)、合轴型克隆植物(clonal plant with sympodial branching pattern)和混合型克隆植物(clonal plant with mixpodial branching pattern)[16]。据同一克隆的分株离散程度的不同,可分为“游击型(guerilla)”克隆植物、“密集型(phalanx)”克隆植物以及两者的过渡类型。“游击型”克隆植物同一基株的克隆分株间距离较大,扩展速度快,分株的分布比较分散;而“密集型”克隆植物同一基株的分株间距离较小,扩展速度慢,分株分布密集[16]。在高等植物中,几乎全部的苔藓植物、绝大多数的蕨类植物和部分被子植物都属于克隆植物[17—18]。克隆植物分布范围广泛,几乎存在于所有类型的生态系统中,并通过其克隆生长在生态系统中起着不可或缺的作用[19]。
克隆生长指在自然条件下,生物自发地通过营养方式形成遗传上相同,且具有潜在独立性的新个体或分株的能力或过程[20]。利用克隆生长这一特性,克隆植物可以产生由匍匐茎或根状茎连接着的数量众多的相同基因型的分株,从而实现自身数量的增加。例如,大米草(SpartinaanglicaHubb.)利用自身的克隆生长能力,在适宜的环境下大量繁殖[21]。除克隆生长外,克隆植物另一个重要特征为双重构件性。即它既有大部分植物都有的有机构件性,又兼备克隆植物所特有的克隆构件性。克隆植物可以改变自身的性状(如间隔子长度、分枝强度和分枝角度等),在其个体周围生成大量自由分布的克隆分株[22],进而对异质生境进行适应。正是由于克隆植物拥有非克隆植物所不具有的特征,利用不同类型的克隆器官实现自身的水平或垂直方向的拓展,扩大生态幅,因此在生态系统中占据优势地位[23]。同时,克隆植物拥有持久的生命力和强大的繁殖能力,即使在某些不良或退化环境中(如高海拔、风沙、盐碱胁迫等)也能通过自身调节使环境胁迫的消极影响降到最低[24]。
克隆植物对异质生境的适应能力对生物多样性的维持和植被生态修复起到了重要作用。本文分析和梳理了近年来国内外克隆植物对于异质性生境适应性研究的主要进展并进行了系统性归纳总结,并对未来的重点研究方向提出了几点建议,以期增进更多研究者的关注,希望能够为生物多样性的维持和生态修复等相关研究的开展提供参考。
1 生境异质性的主要特征
1.1 时间异质性和空间异质性
生境异质性包括时间和空间上的异质性。衡量异质性的指标包括尺度、对比度[25]、聚集度、可预测性、空间相关性等,这些指标在时间和空间层面上均有着表现差异[26]。时间异质性表现为持续时间、相邻时间斑块的长短和频率变化以及时间变化过程中的可预测性等。例如,在光资源方面,大到年、季节尺度,小到分、秒尺度上的变化[27]。同时,在季节和昼夜的变化下,光资源的强度和质量也会随之改变。在时间上,可被植物感知的范围是由植物的诱导过程的反应时间和植物的寿命共同决定的,这两者分别是时间尺度范围的上限和下限。超过或低于这一时空尺度,异质性会随之消失[28]。
景观的生态系统组成构成了基本的空间异质性格局,异质性的分布格局在资源可利用性、环境因子的空间特征以及生物种群和群落在景观上的分布等都有所体现,在一定程度上对生物的异质性分布起到了制约的作用。空间异质性通常表现为环境的梯度变化和生态系统的斑块性。除梯度变化外,生态系统的斑块性一般表现为斑块内部资源相对统一,但斑块与斑块间能够通过植物的生长繁殖产生较大差异[4]。这种资源异质性的普遍存在大大增加了植物吸收和利用必需资源和适应方面的难度[7]。
1.2 资源异质性和环境异质性
从生境功能层面上,还可将其划分为资源和环境异质性。资源异质性是指生物所需资源(如光照、水分、养分等)在时间和空间上的不规则分布。环境异质性指环境要素(如重金属、病原菌、动物觅食和干扰等)在空间和/或时间上的非均匀变化[16],其中干扰又可以包括风蚀、沙埋等因素引起的以及由干旱、水淹、盐碱胁迫等极端环境因素引起的干扰。
资源和环境异质性在自然界是相辅相成、不可分割的。在某些情况下,同一种因子即可以作为资源,也可以成为环境胁迫因子,如:水分既是植物生长的必须资源,也能够成为影响植物生长的胁迫环境因子如干旱胁迫、水淹胁迫等。近年来,国内外学者针对克隆植物对异质生境适应性方面开展了大量的研究,其中对资源异质性的研究更为普遍,例如,薇甘菊(MikaniamicranthaKunth)在异质养分条件下,利用克隆整合提高低养分斑块分株的生物量,降低高养分斑块生物量[29];大米草在异质光照条件下,异质遮阴分株的叶片数、根长和生物量均显著高于同质条件[30];对环境异质性的研究较少,大多集中在干旱胁迫、盐碱胁迫、沙埋等方面。例如,在对油蒿(ArtemisiaordosicaKrasch.)的研究中发现,随着干旱程度加剧其相对生长率和比叶面积随之减小,根冠比随之增大[31];在对羊草(Leymuschinensis(Trin.) Tzvel.)的研究发现,盐碱胁迫经常与刈割干扰共同结合在一起对其空间拓展能力进行调节,低密度、低盐碱胁迫以及刈割生境组合下无性系空间拓展能力最强[32]。
2 克隆植物功能性状
2.1 克隆植物的繁殖方式
有性繁殖和无性繁殖是自然生态系统中存在的两种最基本的繁殖方式。对于大部分克隆植物来说,既可以运用无性繁殖的方式实现植株数量的增多,又可以通过有性繁殖方式增加植株的遗传特性(图1)。例如,在资源相对缺乏的异质生境下,克隆植物独特的克隆生长特性可以支持植株正常生长发育和繁殖,并且在一定程度上提高种间竞争优势,实现死亡风险分摊[33]。但克隆植物的这种繁殖方式也有一定的局限性:对无性繁殖投入精力较多,往往导致有性繁殖投资的减少[34]。尽管无性繁殖可以大大提高克隆植物的扩增能力,但有性繁殖对于克隆植物多样性的维持及远距离扩散起着不可或缺的作用[17]。另一方面,间隔子(如匍匐茎、根状茎、水平根等)的连接使得病原体更容易在植株间运输,加剧了某些疾病的传播。除此之外,克隆分株由于扩散效率较低通常导致大量分株在母株旁边堆积,加剧了植物自身的种内竞争,影响植株的资源获取[35]。

图1 克隆植物在异质性生境下的繁殖策略及生境适应特征Fig.1 Reproductive strategies and habitat adaptation of clonal plants in heterogeneous habitats
2.2 克隆植物的适应对策
2.2.1克隆整合
克隆整合全称克隆内生理整合,指同一基株的分株在一定时间内通过匍匐茎、根状茎和/或水平根等横生结构相互连接在一起,相连的克隆分株间存在的物质传递现象[16]。一般来说,克隆植物会发生从年老分株向年幼分株传递营养物质的向顶运输,当老年分株的吸收能力不足以提供资源运输到年轻分株时,也会发生向基运输[36]。克隆整合强度还与环境的异质程度以及资源的可利用度密不可分,异质性越大,克隆整合强度也越大。同时,克隆整合行为的发生也是有一定阈值的,在极端异质条件下,若异质性程度超出了植物所能承担的阈值,那么植物将无法发生克隆整合行为[37—39]。当距离过大导致植物无法感知生境异质性的存在时,克隆整合也将无法发生[40]。
2.2.2形态可塑性
形态可塑性是克隆植物适应异质生境的优势策略,它一般表现为克隆构型、植株个体形态结构和生物量分配等方面的变化[41]。异质生境中,克隆植物可以通过在水平方向上改变克隆构型以及在垂直方向上改变分株本身的形态结构的方式对不同的资源水平做出反应[42],这种改变自身形态对不同环境进行适应的现象称为克隆植物的形态可塑性。克隆植物在对自身有利和不利斑块中形态可塑性的表现也是不同的。在有利斑块中,形态可塑性主要表现为器官的增生,植株一般会通过加粗匍匐茎,减少节间长度和增加分枝数量的方式最大程度的对斑块资源进行利用;而在不利斑块中,植株为了尽快逃离不利生境,减少自身损失,主要表现为简化结构以及减慢自身生长,如使匍匐茎变细,增加节间长度和减少分枝数等[43]。克隆植物正是通过利用自身形态的可塑性变化,提高了获取必需资源的能力。
2.2.3克隆分工
克隆分工指相互合作的分株为了更有效地获取不同资源,伴随着克隆整合产生的形态与生理功能的协调特化[16]。克隆植物发生劳动分工的条件有4个,分别是资源的异质性分布、构件单元的专化、构件单元间的合作和分株独立生长的能力[44]。根据发生条件的不同,克隆分工可以分为环境诱导型和遗传型两种[45]。当斑块中一种资源丰富而另一种资源贫瘠时,分株发生的克隆分工就会专化吸收资源丰富斑块中的资源,并且通过相连分株运输给资源贫瘠斑块中的植株,发生这种分工的类型称为环境诱导型[46];克隆分工的另一种类型是遗传型,它只与植株自身的遗传条件有关,并且会在某一特定阶段发生形态或功能的专化。克隆植物正是利用这两种类型的克隆分工,一方面提高了自身对异质资源的获取能力,使其不再受区域资源的限制,另一方面也使自身在竞争关系中获得主动地位,提高其种间竞争力[47]。
2.2.4觅食行为
觅食行为指有机体在其生境内进行的促进对必须资源获取搜寻或分枝过程[20, 22]它包括资源共享、分株选择性放置和分株功能分工3个方面[48—49]。觅食行为一般分为强度觅食和广度觅食两种[50]。以资源在分株间的合理分配为前提,觅食行为将植物生长格局和异质性生境内必需资源的分布格局进行精细协调,使植物趋于规避资源贫瘠的斑块并占据资源丰富的斑块,将基株的死亡风险分摊到克隆分株上,通过耗—益权衡使克隆植物达到最高收益水平,并以此提高整个基株的适合度和自身的存活能力[1, 51]。
2.2.5风险分摊
一般来说,非克隆植物通过植株遗传多样性来对风险进行分摊,克隆植物则通过改变植株的生理和形态特征和子株的分布状况等途径,使分株在某些不利时空条件下适时的独立,进而避免物种的灭绝,实现风险分摊[1]。风险分摊对策分为3种:分株独立、分株形态变化和资源储藏[7]。分株独立一般通过出芽、脱落、断裂等方式将植株分离开,使其成为功能上完整并且生理上相互独立的个体[52—53]。通过这种方式基株产生独立存在的分株,基株的死亡概率就会被分摊到各个子株,植株死亡风险也因此得到降低。分株形态变化指将克隆植物表型可塑性和克隆整合性相结合,从而使各分株能够调节分株的形态和空间放置格局来适应生境异质性[54]。资源储藏是克隆植物应对生境异质性的有效手段[7]。很多克隆植物的克隆器官(如球茎、块茎、匍匐茎和根状茎等)存活时间很久,因此具有很强的储存能力和繁殖的能力[55]。资源储藏使得植株可以将资源充足时期的资源储存到克隆器官中,使其在资源匮乏的时期释放补充[1, 56]。这种行为大大缓解了资源的异质分布对植株本身生长发育所造成的压力,使植株可以从环境胁迫中尽快恢复。
3 克隆植物对异质生境的适应
3.1 克隆植物对异质资源的适应
3.1.1光照
光照异质性常常表现为光照周期、遮阴程度、光质等方面,同时光照异质性的衡量指标也包括了光照的尺度和对比度。充足的光照能够为克隆植物提供更多的能量用于生物量的累积,同时也有利于分株的生长、发育、匍匍茎节间的伸长等。然而大多研究中,克隆植物更加适合轻度的遮阴处理,植物在轻度遮荫条件下可以调节生物量的分配比例,使植物获取资源能力得到有效的提升[57]。然而中度或重度遮光条件下,植物的根冠比往往会增加,这是克隆植物“趋富特化”的典型表现,即将更多的生物量投入到吸收较丰富资源的器官或部分[20],并通过间隔子的连接和传输作用,实现资源从吸收和积累较多的分株向吸收和积累较少分株的转移[47](表1)。
3.1.2养分
土壤养分异质性广泛分布于自然界中,主要是由沉降、凋落物和植物残体等有机物的不均匀投入,在自然界长期过程中形成的[61],它表现为土壤中营养元素的斑块性分布[57]。土壤养分异质性任何一个元素(如尺度、对比度、聚集度等)的改变都会对植物的生理或形态产生不同的影响。土壤异质性程度高的区域可以在土壤内部形成小生境,生境中的物种丰富度得到提升。土壤养分异质性可以影响植物个体发育、繁殖、种群数量和形态特征、群落物种组成和种内竞争等[62],从而导致群落结构和生态系统的改变(表2)。

表1 不同克隆植物对光照异质性的适应特征
3.1.3水分
水分异质性分布在自然界中广泛存在,适当的水分含量有利于植株占据有利生境,过低或过高的水分含量则会影响植物的生长发育和繁殖能力[68—69]。水分整合在克隆植物群落适应异质生境中起着重要作用,它可以缓解环境胁迫(如干旱、水淹等)产生的消极影响[70]。一般来说,水分传输的方向为水分充足的分株向水分贫瘠的分株运输,这种情况是随着水势梯度而发生的[71—72]。另外,水分的流动很大程度上是由源—库动力驱动的,包括顶向和/或基向两种方式[73]。这大大提高了克隆植物在水分异质生境中的生长和存活能力,即使在干旱或水淹的胁迫生境中,克隆植物也可以通过子母株间的连接进行水分的交换和传递。目前,国内外对水分异质研究相对较少,一般用不同高度的淹水、不同频率水位波动以及不同的水淹持续时间来模拟异质水分生境(表3)。
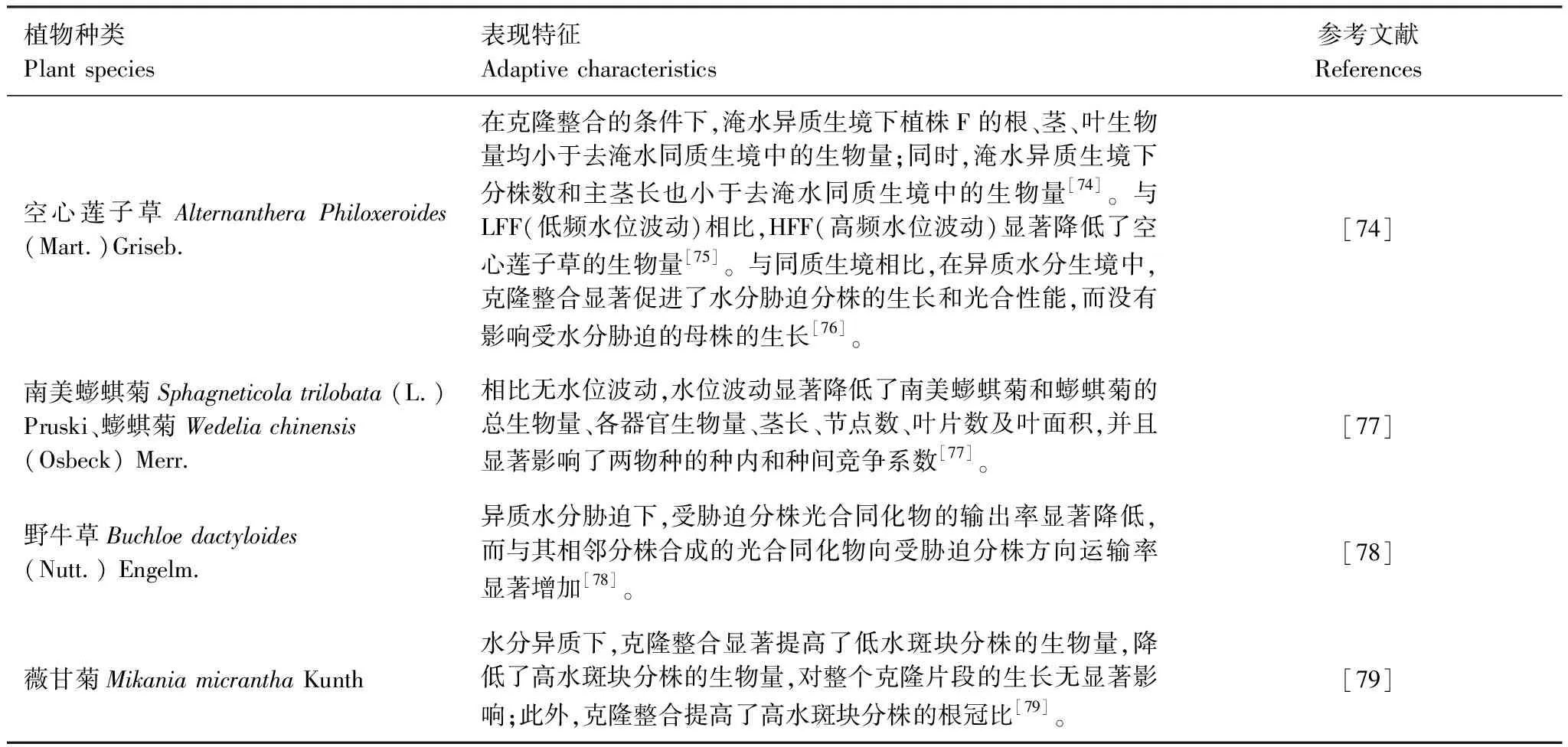
表3 不同克隆植物对水分异质性的适应特征
3.2 克隆植物对异质胁迫环境的适应
3.2.1盐碱胁迫
盐碱胁迫是自然界中威胁植物生存及生长发育最严重的非生物因素之一[80]。由于人为因素(如对自然生态系统不合理、超高强度开发利用等)和非人为因素(如海平面上升、海水入侵、降雨量分布不均等)的影响[81],全球盐碱化程度正处于日益增加的阶段。例如,目前黄河三角洲的盐碱地多达16.7万hm2[82],只有少数具有较高耐盐碱性的克隆植物能在pH为8—9.5的盐碱土里生存[83]。
克隆植物强大的表型可塑性以及克隆整合能力,使得很多克隆植物已经成为盐碱地区的先锋物种,并且在这些生境中表现出极强的抗逆性和适应性。有研究表明芦苇的耐盐范围一般在5‰—25‰之间,甚至可达40‰以上[84],因此被广泛应用于湿地的恢复与重建[81]。自我调节生物量配置模式是植物适应盐胁迫的可塑性机制之一,植物能够通过改变不同器官的生物量分配来适应不同盐分环境[85—86]。例如,在异质盐分胁迫下,蛇莓(Duchesneaindica(Andr.) Focke)单株盐胁迫比相连分株同时受盐胁迫的情况生长的更好,植株通过降低地上生物量,增加地下生物量的方式缓解盐胁迫给自身带来的消极影响[87]。同时,匍匐茎相连的蛇莓母株比断开的母株受胁迫程度较低,并且在母株受胁迫的相连分株重,未受盐胁迫的子株与对照比,其生物量、相对含水量和细胞膜透性未受到显著影响,说明克隆整合促进了植株在胁迫生境的生长[88]。异质性盐分胁迫下,克隆整合提高了积雪草(Centellaasiatica(L.) Urban)受胁迫远端分株和整个克隆片断的叶面积和生物量等生长指标; 与未遭受盐分胁迫处理相比,匍匐茎连接处理导致远端分株的根冠比显著降低[89]。
在对克隆植物群落空间分布格局进行研究时发现,盐碱胁迫下,植物会产生不同的克隆构型对消极生境进行适应。例如,盐碱胁迫干扰中的艾比湖湿地保护区的克隆植物一般表现为密集的集群状分布[90]。因为集群分布使克隆植物的生理整合能力大大增强,能够提升克隆植物的耐盐性。又如罗布麻(ApocynumvenetumL.)和花花柴(Kareliniacaspia(Pall.) Less.)等植物在盐碱胁迫的生境中,集群分布的空间分布特征通过克隆整合作用大大降低了盐碱胁迫带来的消极影响,植物因此也能够更好地生存和生长[90]。但较小尺度(0—0.5 m)下的盐碱胁迫则没有那么显著,为了避免带来更大的竞争压力,克隆植物常常表现为随机分布特征。
3.2.2重金属胁迫
进入土壤的重金属不易被分解,同时富集性强,存留时间长且不可逆,导致了土壤重金属的治理难度较大。土壤中过量的重金属不仅会对植物产生毒害,而且会通过食物链的富集威胁人类和动物的健康[91]。面对不同程度的重金属胁迫,克隆植物会采取不同的应对措施来减少其消极影响。如在对蛇莓的研究中发现,蛇莓会利用选择性放置策略,通过增长匍匐茎和产生不定根的方式进行扩张以逃避有毒生境,将自身分株更多的选择性放置在Pb污染较为轻度的地区,更多的生物量分配到地上以减少地下部分对Pb的吸收,并且子株将更多Pb积累在根部来降低对植株的伤害,同时产生更多分株分摊死亡风险[92]。还有研究表明,在中度Pb污染水平且异质性较强的生境下,蛇莓生长最为理想[93]。Roiloa 和 Retuerto在对野草莓(FragariavescaL.)进行的研究中也得到了相似的结果,他们的结果表明:在异质性铜污染土壤中,野草莓有选择地将无性系分株的定植在未污染的斑块中,受污染的斑块中则更多的是未生根的分株[94]。
然而重金属胁迫中,克隆整合并非对受胁迫分株都是有益的。例如,在对活血丹子株施加Pb胁迫时,克隆整合会以母株生长指标的降低为交换条件,来提高子株的光合能力、生物量及酶活性;当对母株施加Pb胁迫时,母株的光合指标、酶活性和生物量都显著降低,但克隆整合大幅提高了子株的生长指标[95]。总而言之,克隆整合会使整个克隆片段受益,但在遭受胁迫时,会权衡分株的成本和消耗。另外,氮素添加被证实可影响克隆植物对于重金属污染的吸收。如伍海全在研究不同形态氮素对异质性Cd胁迫下克隆植物抗逆性的影响中发现:对活血丹非胁迫端分株施加铵态氮有利于降低Cd胁迫端分株的Cd含量,而硝态氮则会显著提高胁迫端分株的Cd含量;施加铵态氮或硝态氮都能增强胁迫端分株的Cd富集能力,提高它的单株Cd富集量[96]。
3.2.3风沙胁迫
自然界中的沙埋和风蚀改变了植物一系列的生长环境,例如光照、土壤温度、土壤湿度、土壤通气状况以及植物—土壤微环境,这些是植物能否在风沙环境中成功存活的关键因素[97]。一般来说,荒漠中的植物的种子萌发和幼苗生长对沙埋胁迫都有一定的适应能力,一定程度的沙埋可以促进种子萌发和幼苗出土,同时也会刺激根系的生长[98]。但超过植物的生存阈值,也会影响植物生长甚至导致物种淘汰,沙埋因此被作为沙生植物进化的重要选择因素[99]。许多克隆植物利用克隆生长能力很好的适应了风沙环境,当植株遭受沙埋时,会通过增加分株数的方式来抵御沙埋胁迫。例如,在风沙环境中,沙鞭(Psammochloavillosa(Trin.) Bor)和无芒雀麦(BromusinermisLeyss.)利用其克隆生长的能力,从未沙埋的分株获得能量,从而提高自身在沙埋环境中的存活能力[100—101]。除此之外,克隆植物还能利用其极强的扩展能力改善沙地景观,它能比非克隆植物占据大得多的水平空间范围从而使流动沙丘的植被盖度得到一定程度的提高。如一株生长5年的蒙古岩黄芪(HedysarummongolicumTurcz.)再其周围半径3 m内可以萌发200多株根蘖苗,稳定之后可以向周围沙地扩展,是扦插造林的良好树种[102]。
沙生克隆植物利用自身的克隆整合能力,即使在风蚀生境下也有较高的存活率。例如,沙拐枣(CalligonummongolicumTurcz.)的克隆整合使得资源不仅可以在分株间进行传递,还可以在分株-母株间传递,因此,即使母株的全部根系以及风蚀水平根全部裸露在外但仍能存活[103]。除此之外,多年生的克隆植物可以固定风蚀严重的沙丘,增强了风沙景观稳定性[6]。在风蚀生境下,沙鞭可以通过在土壤较深的地方产生新的分株进行修复,赖草(Leymussecalinus(Georgi) Tzvel.)则可以通过改变间隔子长度,将更多的新生分株放置在风蚀坑内。同时,它们可以利用风蚀坑中充足的光照产生大量的分株,从而对风蚀坑进行修复[82]。基于以上特征,克隆植物已被广泛运用到沙生景观的恢复。
4 总结与展望
克隆植物以其独特的生境适应策略、多样的繁殖模式引起众多生态学者的关注。长期以来,克隆植物对生境异质性的生理生态响应一直是植物生态学领域的热门。目前,国内外研究学者对资源(光照、养分和水分)异质生境以及胁迫异质生境克隆植物的适应对策均已开展了不同程度的研究,但综合进行分析,一些方面仍有些薄弱,主要为:(1)当前研究多集中在空间异质性模拟及调查研究上,然而对时间异质性(如春旱夏涝等)条件下克隆植物的响应特征研究为数不多。另外,多数模拟控制实验周期较短,大部分只涉及植物的幼苗生长阶段,虽然该阶段植被响应特征较为明显,但植物成熟期以及繁殖生长的响应研究还较少;(2)克隆植物对生境异质性的响应研究多集中于非生物因子的模拟,而对于生物因子(如土壤动物、微生物等)的调控作用研究则非常有限;(3)如何权衡克隆植物的入侵性及其合理利用仍是研究者重点关注的科学问题;(4)筛选克隆植物修复退化生态系统的优良品种,做好生态安全风险评估工作对于生态修复也极为重要。
克隆植物在自然生态系统中扮演着重要的角色,它与生态修复、生物入侵、全球变化、生物进化等生态学前沿领域紧密相关,结合克隆植物对异质生境适应性研究热点和发展趋势,在未来研究中重点关注以下几个方面对克隆植物适应对策研究的进一步发展将有很好的推动作用:
(1)时间异质性尺度上的考量。自然界中的生境因子往往是随着时间变化而产生波动的,如土壤盐分会随着气温的升高(日波动或季节波动)呈现“表聚”的特征,而当前大部分研究是针对生境因子异质性的静态模拟。在未来的研究过程中,应考虑将生境因子随时间的变化特征与空间异质性联合起来,更全面的探索克隆植物的响应特征。另外,克隆植物不同生长时期对于生境异质性的生理生态适应特征有巨大的差异,仅从幼苗期或营养生长期的角度探讨其对生境异质性响应机制并不能完全解释其适应性进化特征,特别是克隆植物繁殖分配对生境异质性的响应机理还知之甚少。下一步有必要从野外原位控制及长时间尺度监测的角度分析克隆植物不同生长期的适应机制。
(2)异质性生境中生物因子对克隆植物适应的调控作用。以往研究多关注于克隆植物对于非生物因子异质性的生理生态响应(如养分异质性生境的根系分配策略),然而自然界中土壤生物因子(如土壤动物、土壤微生物等)同样具有重要的调控作用,甚至可能会重塑克隆植物的响应。因此生物因子在异质性生境中的调控作用应该在未来的研究中广泛关注。
(3)克隆植物的入侵性及其合理利用。近些年,以植物入侵主导的生物入侵问题已经成为全球重大环境问题之一。而克隆植物是外来入侵植物的主要类群,探索克隆植物的入侵机制及其与本地近缘种的竞争关系,分析两者在异质生境中的生理和形态差异,有利于科学的控制外来植物入侵,维持本地生态系统的平衡和稳定,该方面内容虽然在近两年已经引起了部分学者的重视[66, 104—105],但仍需深入研究。
(4)筛选性状优良的克隆植物品种用于退化生态系统修复。克隆植物多样化的生境适应对策和强大的繁殖能力,作为退化生态系统修复的关键物种具有天然的优势。然而,克隆植物的克隆性与入侵性有着密切的关系,若利用不当,会危及生态的健康稳定发展。因此,未来在修复物种选择上,注重友好型替代物种的开发,做好生态安全风险评估工作也是研究者需要重点关注的内容。
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
落叶果树(2021年6期)2021-02-12 01:29:10
今日农业(2020年24期)2020-12-15 16:16:00
兽医导刊(2016年12期)2016-05-17 03:51:50
体育科技(2016年2期)2016-02-28 17:06:07
云南师范大学学报(自然科学版)(2015年5期)2015-12-26 12:46:16
中央民族大学学报(自然科学版)(2015年2期)2015-06-09 08:45:26
物理实验(2015年10期)2015-02-28 17:36:52
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
现代检验医学杂志(2015年4期)2015-02-06 02:02:06
