
高产杂交棉F1 与双亲的光合性能及相关基因表达比较
2022-06-23 02:06:50唐丽媛李兴河王海涛张素君刘存敬张建宏
河北农业大学学报 2022年3期
唐丽媛,李兴河,王海涛,张素君,蔡 肖,刘存敬,张建宏
(河北省农林科学院 棉花研究所/农业部黄淮海半干旱区棉花生物学与遗传育种重点实验室/国家棉花改良中心河北分中心,河北 石家庄 050051)
棉花是重要的经济作物,在我国国民经济中占有十分重要的地位[1]。在当前棉花种植面积持续下降的形势下[2],充分利用杂种优势[3-6]育成强优势杂交棉,可有效提高棉花产量。
棉铃发育85% 以上所需能量来自棉铃对位叶[10],是产量形成的主要源器官,而光合作用是作物产量形成的根本来源,是所有生理进程的基础[11-12],对植株产量及品质的形成至关重要[13],且能够稳定遗传[14-15],因此通过优势杂交种棉铃对位叶的光合生理特性分析可解析其产量形成,但棉花上针对棉铃对位叶进行光合生理特性的研究还较少[16]。光合作用是受多基因和蛋白精细调控的复杂性状,随着分子生物学飞速发展,目前该研究不再只局限于生理指标测定,而通过基因表达、基因组学等现代分子手段解释不同作物的杂种优势机制[17-18]。光合机构的形成由核质基因共同编码,其中核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)是决定碳同化速率的关键酶,也是光呼吸中不可缺少的加氧酶,由8 个大亚基和8 个小亚基组成[19],Rubisco 小 亚 基(Rubisco smaller subunit, RbcS)由核基因组编码,大亚基(Rubisco larger subunit,RbcL)由叶绿体基因组编码[20],在光合作用调控网络中起着重要作用,但其基因表达在棉花中的报道不多。本研究以高产杂交棉‘冀1518’[21]及双亲为试验材料,比较其棉铃发育各时期棉铃对位叶的光合性能指标差异,分析光合相关基因表达特征,从光合性能层面解析杂交棉‘冀1518’获得高产的生理机制,以期为今后更好的利用杂种优势定向创制新种质、培育高产杂交种提供理论依据。
1 材料与方法
1.1 试验材料
以高产杂交棉‘冀1518’[21]及双亲为试验材料,母本为‘冀228’[22],父本为‘冀567’,材料均由河北省农林科学院棉花研究所提供。
1.2 试验地点及试验设计
试验于2018—2019 年在河北省农林科学院棉花研究所石家庄小安舍试验站进行,供试土壤为砂壤土,地力均匀,前茬作物为棉花。2018 年4 月25日播种,5 月14 日移栽;2019 年4 月23 日播种,5月13 日移栽。试验采用随机区组设计,设置3 次重复,小区面积7.1 m×6 m,行距0.75 m,株距0.25 ~0.30 m,将每小区一半作为田间取样区,一半作为调查、测产区。小区周围设置保护行。田间管理同一般试验田管理。
1.3 取样
本研究以棉铃对位叶为研究对象,棉花盛花期对植株中部第6 ~8 果枝的1 ~2 果节在开花当天挂牌标记[10],分别取开花后0DPA(Days post anthesis)、10DPA、20DPA、30DPA、40DPA 共5个时期棉铃对位叶,每个时期取10 片,经液氮速冻保存于- 80℃冰箱待测相关酶活性及基因表达。
1.4 测定指标与方法
1.4.1 光合数据及叶绿素含量测定 对标记的棉铃对位叶自开花当日起每隔10 d 测定光合性能指标,测定时间与取样时间同步,0DPA ~40DPA 分5个时期完成测定。于晴天北京时间9:00—12:00 用Li6400 便携式光合测定仪,测定各材料对应叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),为减小环境中自然条件不稳定造成的误差,使用内置人工光源和叶室控温装置,并开启空气干燥器,光照强度设置为1 200 umol/(m2·s),叶室温度设为30 ℃固定值[16],当光合速率数值在设置的光照强度和叶室温度条件下稳定时进行测定,每个小区测5 片叶,取平均值;同时用日本产SPAD-502 型叶绿素计测定该叶片叶绿素含量,每个小区测5 片叶,取平均值。
1.4.2 水分利用效率和羧化效率 利用测定光合数据计算水分利用效率(WUE,)和羧化效率(CE),作为衡量光合特性指标。WUE=Pn/Tr[23];CE=Pn/Ci[24]。
1.4.3 超氧化物歧化酶、过氧化物酶酶活及丙二醛含量测定 分别采用苏州科铭生物技术有限公司试剂盒测定超氧化物歧化酶(SOD)、过氧化物酶(POD)酶活和丙二醛(MDA)含量。
1.4.4 叶面积测定和棉铃调查 每调查区选择连续5 株生长一致的棉株,在盛花期(7 月25 日)、结铃盛期(8 月20 日)和吐絮期(9 月20 日)对棉株果节数、脱落数进行调查,并测定各果节对位展开叶最大叶面积。
1.4.5 杂种优势计算 应用杂种优势率和超亲优势率度量杂种优势程度。杂种优势率(中亲优势率)=(杂交种性状值-父母性状本均值)/父母本性状均值×100%[16],>0 为正向优势,<0 为负向优势;超亲优势率=(杂交种性状值-高值亲本性状值)/高值亲本性状值×100%[25]。
1.4.6 产量及产量构成因素测定 于吐絮期调查各小区农艺性状,计算单株铃数;待棉铃成熟自然吐絮风干后,每小区收获10 株进行单株籽棉产量测定;收获中部50 个棉铃称重后计算单铃重;收获测产区全部籽棉,经混合取样后考种轧花计算衣分及皮棉产量。
1.4.7 叶片光合相关基因表达测定 使用普洛麦格(北京)生物技术有限公司试剂盒提取不同时期叶片样品的总RNA,利用Takara PrimeScript™ RT reagent Kit 反转录合成第1 链cDNA,以第1 链cDNA 为 模 板 在CFX96 定 量PCR 仪(Bio-Rad,USA)上进行实时荧光定量PCR(qRT-PCR)。查找光合相关基因[19,26],使用Primer Premier 5.0设计特异引物(表1),苏州金唯智生物科技有限公司合成引物。按照北京艾德莱生物科技有限公司2×Sybr Green qPCR Mix 推荐的反应体系进行PCR扩增。每个样品设置3 次生物学重复,以陆地棉组蛋白基因Histone3为内参基因,采用2-△△CT法分析基因相对表达量。

表1 光合相关基因的引物Table 1 Photosynthetic genes primer sequences
1.5 数据分析
所有数据使用Excel2010 软件进行处理,计算两年均值及标准差,使用DPS15.1 对各性状指标进行方差分析,以最小差数法(LSD 法)进行多重比较,使用SigmaPlot 14.0 及R 语言作图分析。
2 结果与分析
2.1 ‘冀1518’产量杂种优势表现
对‘冀1518’及双亲主要产量性状(表2)分析,‘冀1518’2 年产量性状均表现出了显著的杂种优势。
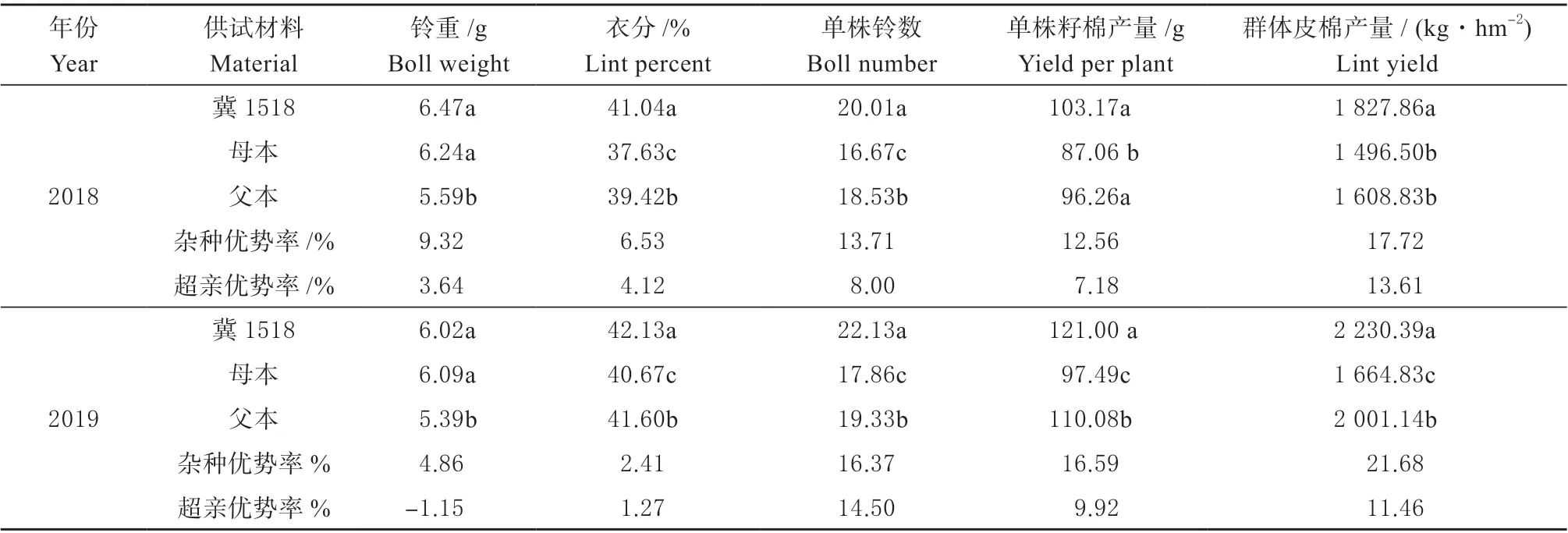
表2 ‘冀1518’及双亲主要产量性状Table 2 Main yield traits between ‘Ji1518’ and its parents
2 年平均,‘冀1518’单株籽棉产量比两亲本分别高21.46%、8.64%,群体皮棉产量比两亲本分别高28.37%、12.42%,与父母本均呈显著差异,表现出较强的超亲优势;产量构成因素中,‘冀1518’的平均铃重、衣分及单株铃数均表现出超亲优势,单株铃数(11.25%)>衣分(2.70%)>铃重(1.25%),其中衣分和单株铃数与双亲差异达显著水平。
2.2 ‘冀1518’及双亲单株棉铃对位叶相关性状表现
对‘冀1518’及双亲单株棉铃对位叶相关性状(表3)分析发现,‘冀1518’总果节数比父本少但差异不显著,而显著多于母本,其所对叶面积同样比父本小但差异不显著,而显著大于母本;‘冀1518’的成铃率、单株成铃对位叶总叶面积和平均棉铃叶面积均高于双亲,表现出4.92%~21.16%的超亲优势,说明‘冀1518’在实际成铃及有效光合产物生产方面具有超亲优势。
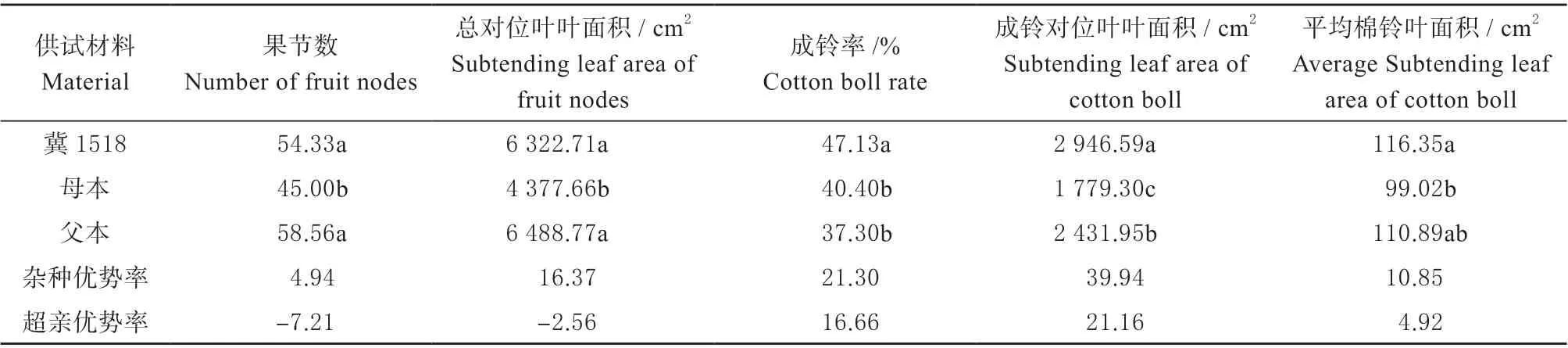
表3 ‘冀1518’及双亲单株棉铃对位叶相关性状Table 3 Subtending leaf of cotton boll related traits per plant between ‘Ji1518’ and its parents
2.3 ‘冀1518’及亲本不同时期棉铃对位叶光合性能差异
分析‘冀1518’及亲本棉铃对叶位光合性能指标的反应如图1 所示。
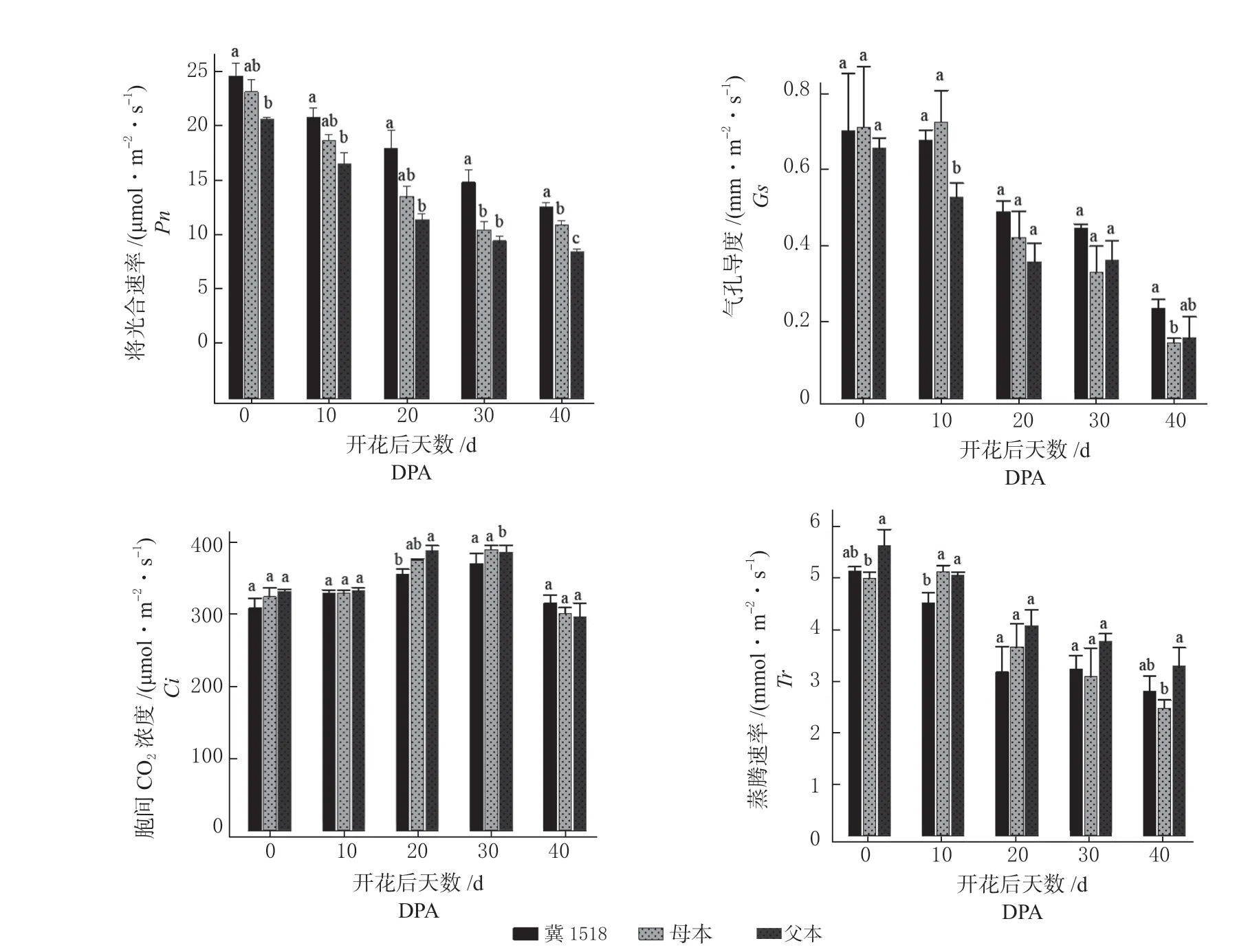
图1 ‘冀1518’及亲本棉铃对叶位光合性能指标分析Fig.1 Analysis of photosynthetic properties of subtending leaf of cotton boll between ‘Ji1518’ and its parents
Pn随棉铃增大呈逐渐降低趋势,‘冀1518’Pn整个棉铃发育阶段均高于双亲,20DPA ~40 DPA与父母本差异均达到显著水平,其中20DPA、30DPA超亲优势率均达23.53%以上;Gs随棉铃发育逐渐降低,Gs在棉铃发育前期(0DPA ~10DPA)介于母本和父本之间,自20DPA 均高于双亲,但仅在40DPA 与母本差异显著;Ci在棉铃整个发育阶段呈单峰曲线趋势,3 个材料均在30DPA 达到最高值,‘冀1518’在20DPA 低于双亲且与父本差异显著,其他时期各材料间差异均不显著;Tr在棉铃发育阶段整体呈逐渐下降趋势,‘冀1518’在10DPA 极显著低于双亲,而其他各时期值多介于双亲间且与双亲差异不显著。
2.4 ‘冀1518’及亲本不同时期SPAD、WUE 及CE 分析
SPAD在整个棉铃发育阶段呈逐渐下降趋势(图2),‘冀1518’与双亲始终无显著差异,杂种优势不明显;WUE随棉铃发育未见明显的一致性规律,其中‘冀1518’在10DPA ~30DPA 保持较高的水分利用率,显著高于双亲,超亲优势达24.25% ~42.89%,0DPA 和40DPA 与母本差异不显著且在40DPA 低于母本;CE 整体呈下降趋势,‘ 冀1518’ 整 个 时 期 高 于 双 亲, 表 现 出9.09% ~30.93%的超亲优势,与母本在20DPA、40DPA 差 异 显 著, 与 父 本 在0DPA ~20DPA、40DPA 差异显著,3 个材料在30DPA 无显著差异。
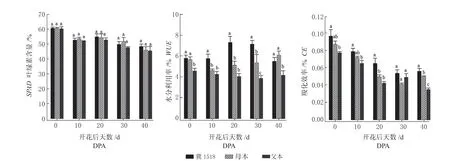
图2 ‘冀1518’及亲本棉铃对叶位叶绿素含量(SPAD)、水分利用率(WUE)及羧化效率(CE)分析Fig.2 Analysis of SPAD, WUE and CE of subtending leaf of cotton boll between ‘Ji1518’ and its parents
2.5 ‘冀1518’及亲本不同时期叶片细胞保护酶活性差异
图3 分析表明,SOD和POD活性均呈单峰曲线,SOD活性中‘冀1518’和父本在20DPA 达到峰值,‘冀1518’高于双亲并与母本差异显著,其他时期值均介于双亲值之间,‘ 冀1518’在10DPA ~40DPA 的 叶 片SOD活 性 表 现 为0.02%~12.83%的中亲优势;3 个材料的POD活性0DPA ~20DPA 均较低,在30DPA 迅速达到峰值,‘冀1518’在各时期均高于双亲值,表现为16.14% ~31.34% 的 超 亲 优 势, 在10DPA、30DPA、40DPA 与父母本均差异显著;随棉铃增大,棉铃对位叶逐渐衰老受损,‘冀1518’及其双亲MDA含量持续升高,‘冀1518’MDA含量及变化与母本相似,增长速率较慢且自30DPA 以后MDA含量显著低于父本,整个时期表现为-0.57%~-26.62%的负向优势。以上结果表明,‘冀1518’为棉铃发育提供主要营养的对位叶,在棉铃发育阶段尤其是中后期(30DPA ~40DPA),叶片细胞保护酶生理活性高而细胞损伤慢,因此叶功能期较双亲延长。
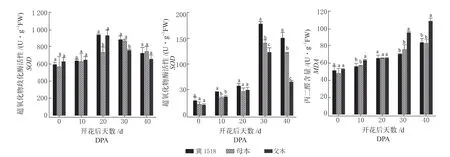
图3 ‘冀1518’及亲本棉铃对叶位超氧化物歧化酶(SOD)、过氧化物酶(POD)和丙二醛(MDA)分析Fig.3 Analysis of SOD, POD and MDA of subtending leaf of cotton boll between ‘Ji1518’ and its parents
2.6 ‘冀1518’及亲本光合生理性能相关指标的相关性分析
对各时期棉铃对位叶光合生理性能相关指标进行Pearson 相关性分析,结果如图4 所示,‘冀1518’与双亲的各项指标间相关性及其显著程度大致相同,Pn与Gs、Tr、SPAD、CE均呈极显著正相关,在一定范围内各指标变化一致,而与MDA、SOD、POD均呈显著以上水平负相关,说明随光合速率降低,叶片走向衰老,细胞保护酶开始活跃并发挥作用;Gs与Tr、SPAD、CE均呈极显著正相关;Tr与SPAD、CE均呈极显著正相关,与WUE呈显著负相关;SPAD与CE呈极显著正相关。不同的是‘冀1518’的Ci与Tr、CE显著负相关,Ci与WUE、WUE与SOD极显著正相关,而在亲本中相关性均不显著;‘冀1518’的SOD与POD正相关未达显著水平,而在亲本中极显著正相关。
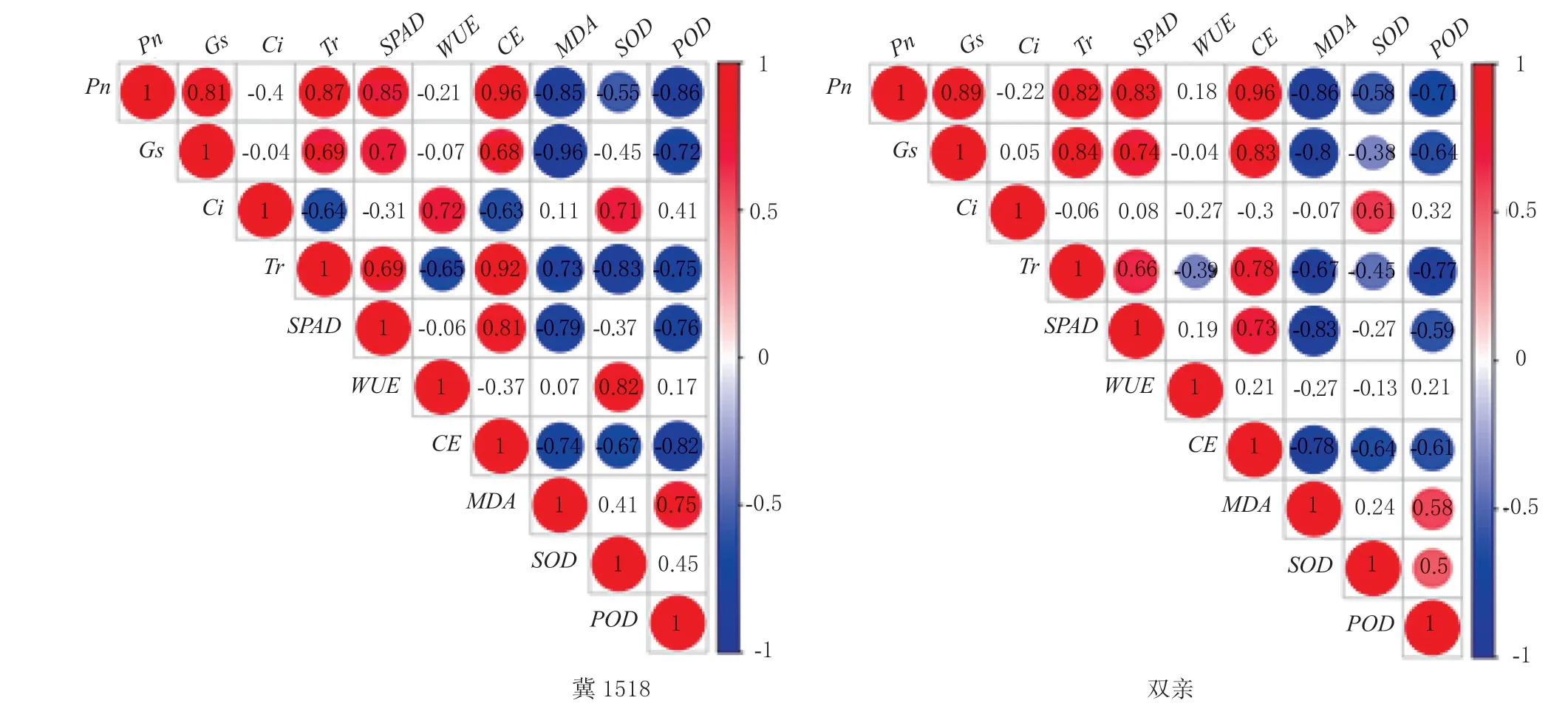
图4 ‘冀1518’与双亲棉铃对位叶光合生理性能指标的相关性分析Fig.4 Correlation analysis of photosynthetic physiological properties of subtending leaf of cotton boll of ‘Ji1518’ and its parents
2.7 ‘冀1518’棉铃对位叶光合生理性能杂种优势分析
‘冀1518’棉铃对位叶光合生理性能各项指标的杂种优势率和超亲优势率统计表明(图5),Pn,CE和POD在棉铃发育的各个时期均具有较强的超亲的杂种优势;Gs,WUE各时期为正 向 优 势, 且Gs在20DPA ~40DPA、WUE在10DPA ~30DPA 为 超 亲 优 势;Ci在40DPA 前 均表现为负向优势,40DPA 是转为超亲优势;MDA在开花当日表现为正向优势,之后整个时期表现为负向优势;SOD与MDA相反,在开花当日表现为负向优势,之后各时期均表现为正向优势,并在20DPA ~30DPA 表现出较低的超亲优势;Tr在10DPA 时负向优势,其余时期为正向优势;SPAD在10DPA 和30DPA 时表现为负向优势,其余时期为正向优势,但超亲优势不明显。可见,‘冀1518’棉铃对位叶的杂种优势随棉铃开始就已体现,并随棉铃发育进程推进而更加显著,尤其Pn,CE、POD和WUE的杂种优势体现最为明显。
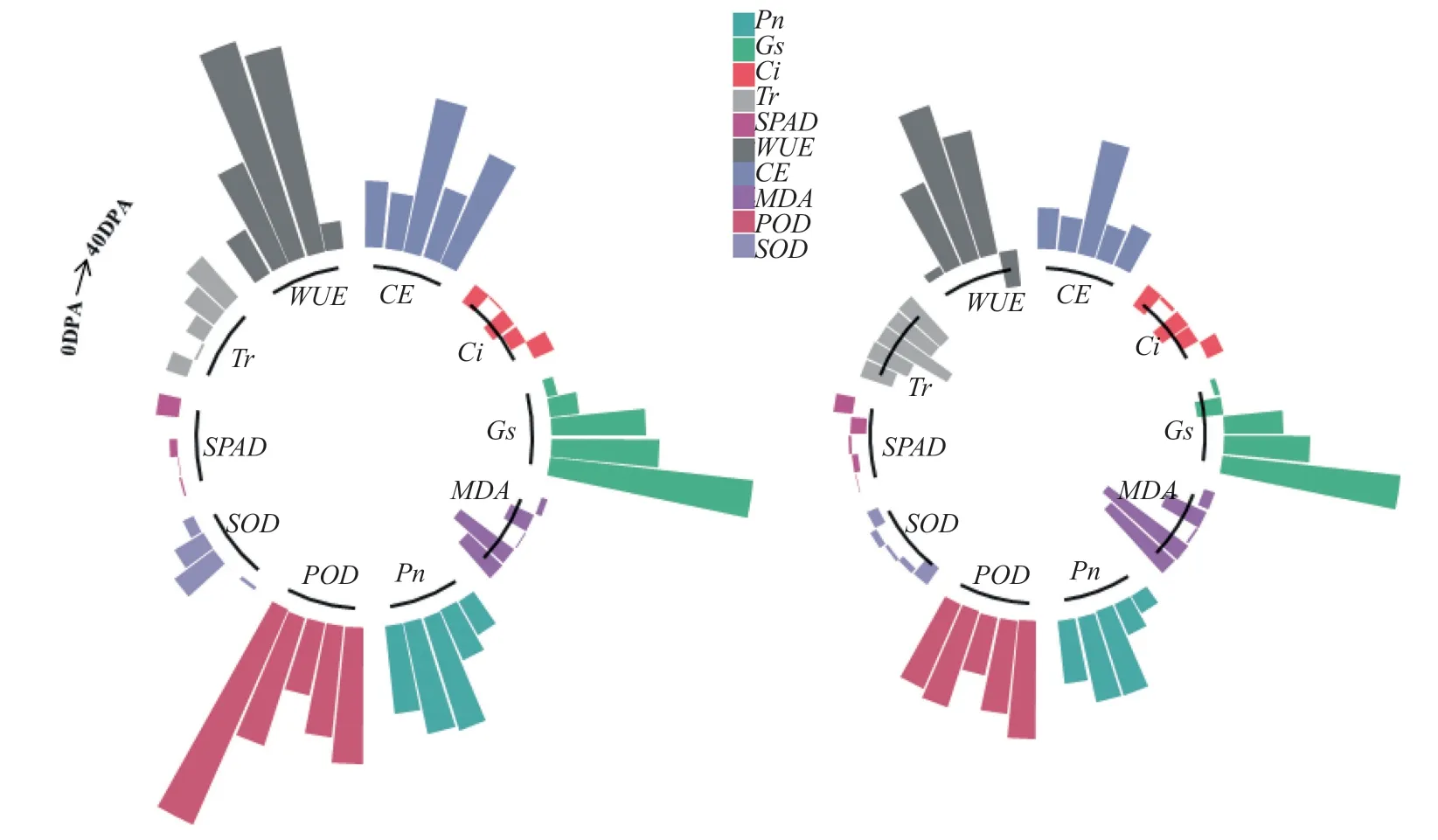
图5 ‘冀1518’棉铃对位叶光合生理性能的杂种优势分析Fig.5 Heterosis analysis of photosynthetic physiological properties of subtending leaf of cotton boll in ‘Ji1518’
2.8 棉铃对位叶光合相关基因表达分析
本研究选取陆地棉中Rubisco 的大、小亚基各1 个基因及活化酶(Rubisco activase, RCA)基因进行表达量测定。图6 随着棉铃对位叶衰老,‘冀1518’与双亲的RbcL基因表达迅速下调,至30DPA ~40DPA 表 达 量 均 极 低,0DPA ~20DPA棉铃对位叶各时期在‘冀1518’中表达量显著高于双亲;‘冀1518’与双亲的RbcS基因表达量均呈缓慢下降趋势,其中在‘冀1518’与母本中表达量仅在10DPA、40DPA 差异显著,而在各时期与父本均差异显著; RCA 基因在‘冀1518’和母本中先上调表达,在20DPA 达到峰值后下降,父本则呈现持续下调的趋势,在‘冀1518’中的表达量在0DPA ~20DPA 未表现优势表达,在中后期30DPA ~40DPA 则超亲表达。
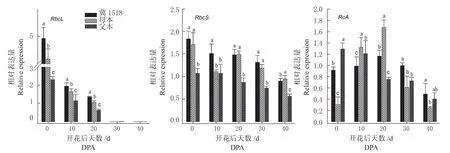
图6 ‘冀1518’及亲本棉铃对叶位光合相关基因表达分析Fig.6 Relative expression analysis of photosynthetic genes of subtending leaf of cotton boll between ‘Ji1518’ and its parents
3 讨论与结论
较大的叶片光合面积为作物高产优质提供基础[13],并主要影响对产量贡献最大的铃数、铃重等[27]。本研究‘冀1518’产量构成各因素中平均铃重、衣分及单株铃数均表现出超亲优势,单株铃数>衣分>铃重;‘冀1518’单株总果节数和叶面积表现杂种优势但均较父本低而显著高于母本,说明‘冀1518’和父本均具有较强成铃潜力;‘冀1518’在实际成铃率和有效成铃对位叶叶面积均表现了明显的超亲优势,并最终获得超亲优势的产量,说明提高有效果节数和有效叶面积,更有利于棉株群体产量发展,是棉花取得高产的重要因素,这与前人研究结果一致[28-29]。棉铃对位叶是棉铃发育的主要源器官,对棉铃发育贡献最大,其生理代谢状况直接影响棉铃生物量的积累[10]。本研究‘冀1518’棉铃对位叶在主要发现光合指标中表现出显著杂种优势,具有更强的光合能力,整个时期尤其是中后期表现出高净光合速率、低胞间CO2浓度、高气孔导度、高羧化效率和高水分利用率。研究与Dong 等[30]的研究结果一致,高光合能力的杂交种在产量上较双亲常规种表现出优势。
进一步探索高产杂交棉F1具有高光合能力的原因,发现随棉铃开始发育,虽然基因RbcL、RbcS随棉铃对位叶衰老均下调表达,但‘冀1518’中基因RbcL在各时期表达量均表现出超亲优势,RbcS呈超亲优势或正向优势,RCA在30DPA ~40DPA表现出超亲优势。复杂的光合基因网络调控的同时,叶片激发细胞保护酶活性从而减缓细胞衰老损伤速率,‘冀1518’棉铃对位叶SOD和POD活性自10DPA 后均表现出杂种优势,尤其POD活性整个时期具有超亲优势,有效的抑制了MDA的增长。
但与葛勇等[16]研究认为叶绿素含量表现明显的杂种优势以维持高效率的光合作用结果不同,本研究中‘冀1518’叶绿素含量在整个时期与双亲差异不显著,并未表现或只在某些时期表现微弱的杂种优势,在‘冀1518’棉铃光合生产效率显著的杂种优势中未起到关键作用,这可能是由于‘冀1518’双亲自身叶功能均较好,虽然叶绿素与光合速率呈极显著正相关,但这一性状在本研究材料中所起到的杂种优势作用不显著。因此,‘冀1518’在杂合基因调控下,具有较大的叶片光合面积,棉铃对位叶通过Rubisco 和Rubisco 活化酶等光合相关基因的表达保持较高活力,使棉铃对位叶光合能力强,同时细胞保护酶的杂种优势延缓叶片衰老,营养物质运转快,光合产物积累多,为棉铃提供丰富的养分。这是‘冀1518’表现出产量杂种优势的主要原因。本研究为进一步探索杂种优势机理提供依据,为后续利用杂种优势培育高产杂交棉品种提供参考。
猜你喜欢
蔬菜(2021年7期)2021-11-27 03:45:25
河南科技学院学报(自然科学版)(2021年6期)2021-11-15 05:43:06
棉花学报(2021年1期)2021-02-05 09:37:16
东坡赤壁诗词(2020年5期)2020-11-06 07:33:43
中国棉花(2019年3期)2019-03-27 06:56:14
小学生时代·综合版(2016年4期)2016-11-19 08:41:24
山西农经(2016年5期)2016-02-28 14:24:37
化工进展(2015年3期)2015-11-11 09:18:44
茶叶通讯(2014年2期)2014-02-27 07:55:39
生物学杂志(2013年1期)2013-04-11 04:19:35
