
广西新型鸭呼肠孤病毒流行毒株GX01-2020全基因组分子特征分析
2022-06-17 02:11谢守玉刘惠心熊陈勇施开创屈素洁苏文广温新瑞李春英邓桂潮尹彦文
中国兽医学报 2022年4期
谢守玉,刘惠心,周 媛,熊陈勇,施开创,屈素洁,苏文广,温新瑞,李 媛,李春英,邓桂潮,尹彦文*
(1.广西壮族自治区动物疫病预防控制中心,广西 南宁 530001;2.广西大学 动物科学技术学院,广西 南宁 530005;3.北海市动物疫病预防控制中心,广西 北海 536000; 4.钦州市动物疫病预防控制中心,广西 钦州 535099;5.浦北县动物疫病预防控制中心,广西 钦州 535300)
禽呼肠孤病毒(avian reovirus,ARV)宿主谱广泛,不仅能感染鸡、火鸡、鸭、鹅等家禽类[1-4],而且可以感染野鸟[5],给养殖业造成巨大经济损失。1950年,水禽源禽呼肠孤病毒(waterfowl-origin avian reovirus,WRV)在南非首次报道[6]。1972年,法国首次发现经典番鸭呼肠孤病毒(Muscovy duck reovirus,MDRV)[7],随后,欧洲多个国家相继分离出MDRV毒株[8-10]。1997年,我国首次发现MDRV感染引发的疫情。MDRV主要感染3周龄内的雏番鸭,可持续带毒感染至6周龄,病死率为10%~30%[11]。2005年,我国东南沿海地区出现新型鸭呼肠孤病毒(novel duck reovirus,NDRV)引发临床上以肝脏不规则坏死及出血、脾脏严重肿大出血为特征的新发疫病,病死率为5%~50%,宿主谱比MDRV更为广泛,如番鸭、半番鸭、麻鸭、雏鹅等均能感染[12]。NDRV感染可导致患病鸭免疫器官萎缩或坏死,成为严重危害水禽养殖业健康发展的重要免疫抑制性疫病[13]。近年来,NDRV感染呈逐年上升趋势,已蔓延至全国养鸭主产地区,给水禽养殖业造成了重大经济损失。
NDRV为呼肠孤病毒科正呼肠孤病毒属,分片段的双股RNA病毒[14]。NDRV有10个RNA片段,大基因L1、L2、L3,分别编码λ A、λ B、λ C 蛋白;中基因M1、M2、M3,分别编码μ A、μ B、μ NS 蛋白;小基因S1、S2、S3和S4,分别编码P10、P18、σ C、σ A、σ B和σ NS 蛋白[15]。近年来,广东、福建、浙江、山东等地上传了NDRV全基因组序列,经序列分析发现国内的NDRV分离株亲缘性较近,但广西地区的NDRV相关报道很少,广西NDRV全基因组序列分子特征尚不明确。本研究从发病鸭的组织病料中鉴别诊断为NDRV阳性(命名为GX01-2020),对其进行全基因组测序,并与国内外不同地区的NDRV、MDRV及CRV(chicken-origin avian reovirus)进行序列比对,分析GX01-2020全基因组的核苷酸序列及氨基酸序列,以期为广西地区NDRV的流行情况提供数据支持,为NDRV的有效防控和致病机理研究奠定基础。
1 材料与方法
1.1 主要试剂MiniBEST viral RNA/DNA Extraction Kit Ver 5.0(批号:AK2501),PrimeScriptTMOne Step RT-PCR Kit Ver.2(批号:ASF0454A),MiniBEST Agarose Gel DNA Extraction Kit Ver 4.0(批号:AK11054A),pMD18-T载体(批号:ASF1711A),DH5α感受态细胞(批号:AK71029A)均购于TaKaRa公司。
1.2 病料检测病料样品来自2020年广西北海某养鸭场发病的番鸭,采集肝脏、脾脏、肾脏、肺脏等组织样品,放置含有适量PBS溶液(体积比4∶1,pH 7.2)的灭菌离心管中,经组织研磨仪磨至糜状,离心取上清用于总核酸提取,应用自主设计的特异性引物和TaqMan探针进行荧光定量RT-PCR检测。上游引物:5′-GGGTCGCACTACAGAGCAACT-3′,下游引物:5′-CGCCTCATCATAGTAATCTGCAA-3′,探针:CY5-CTTGATCAATATGCCGTTGCTCTGCATG-BHQ3。反应体系为20 μL:Premix Ex TaqTM(Probe qPCR) 10 μL,特异性上、下游引物(20 μmol/L)各0.5 μL,探针(20 μmol/L)0.2 μL,模板3 μL,灭菌双蒸水5.8 μL。反应程序:95℃ 20 s;95℃ 5 s,58℃ 34 s,40个循环,同时收集荧光信号。结果判定:有扩增曲线,且Ct值小于35则判为阳性;无扩增曲线或Ct值大于35则判为阴性。
1.3 病毒全基因组扩增测序以检测结果为阳性的核酸为模板,应用本研究设计的NDRV全基因组测序引物(表1),进行RT-PCR扩增。建立50 μL反应体系:模板2 μL,PrimeScript One Step Enzyme Mix 2 μL,2×One Step Buffer 25 μL,上、下游引物各1 μL(20 μmol/L),无RNA酶灭菌水补足体系。反应程序:50℃ 30 min;94℃ 2 min;94℃ 30 s,52℃ 30 s,72℃ 2 min,35个循环。反应结束后,PCR产物通过1.5%琼脂糖凝胶电泳鉴定。

表1 全基因组测序引物
PCR产物胶回收纯化后,连接至pMD18-T载体,转化DH5α感受态细胞,筛选阳性克隆送至测序公司测序,每个片段重复测序3次。使用DNAStar软件包中的SeqMan对获得的基因片段进行序列拼接,最终得到GX01-2020全基因组序列。
1.4 NDRV全基因组序列分析将拼接后的10段核苷酸序列上传至NCBI的BLAST中进行比对,检验相似性最高的毒株。应用BioEdit软件对GX01-2020及国内外参考毒株(表2)基因组核苷酸序列及其氨基酸序列同源性进行分析。应用MEGA 7软件对比对后的序列进行最佳核苷酸替换模型计算:L1为GTR+G+I;L2,L3为GTR+G;M1为HKY+G+I;M2为K2+G+I;M3为GTR+I;σ A为K2+I;σ B,σ C为K2+G;σ NS为TN93+G+I。以最佳核苷酸替换模型为基础,采用最大似然法(maximum likelihood)绘制系统发育树,Bootstrap值定义为1 000次。应用RDP 4(recombination detection program 4)和SimPlot(ver 3.5.1)软件进行全基因组的重组分析,检测是否有重组。
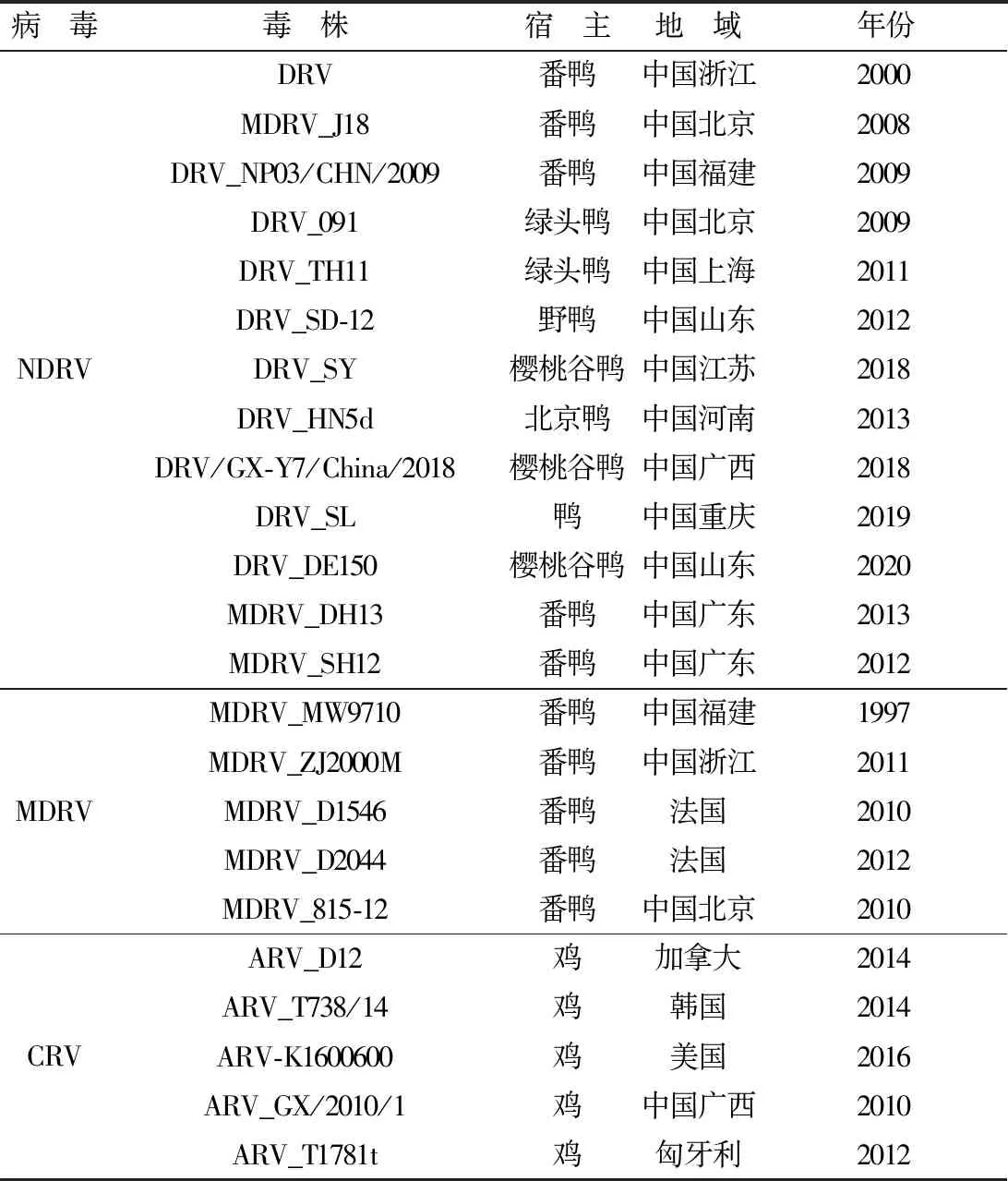
表2 NDRV主要参考毒株信息
2 结果
2.1 全基因组测序结果应用SeqMan软件将测序得到的片段进行拼接,结果显示,GX01-2020株全长为23 353 bp,共有10个基因组片段:L1(3 958 bp),L2(3 830 bp),L3(3 900 bp),M1(2 284 bp),M2(2 134 bp),M3(1 996 bp),S1(1 568 bp),S2(1 290 bp),S3(1 202 bp),S4(1 191 bp)。应用NCBI上的ORF预测结果显示,有9个片段只编码单个ORF,编码的氨基酸大小为97~1 293 aa,而S1编码3个相互有重叠区域的ORF,分别为p10(20~313 bp)、p18(273~761 bp)及σ C(571~1 536 bp)(表3)。
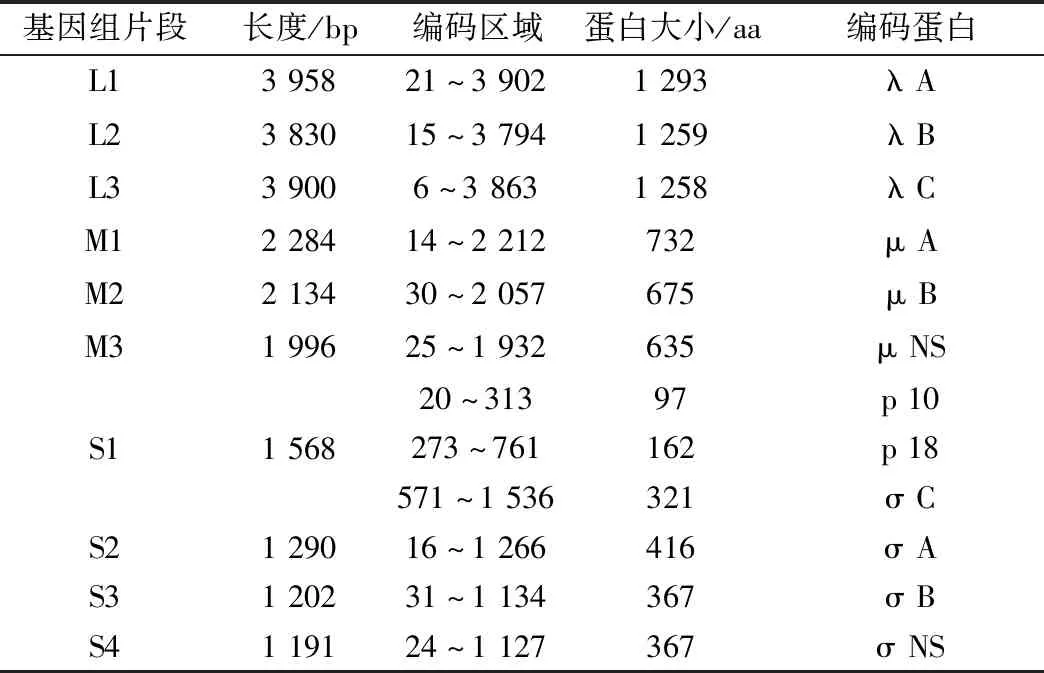
表3 GX01-2020株基因组结构特征
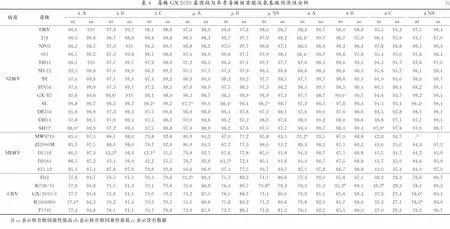
2.2 全基因组核苷酸及氨基酸同源性分析应用BioEdit软件将广西GX01-2020株与国内外参考毒株全基因组核苷酸序列进行比对发现,10个片段的基因组序列与参考毒株的核苷酸序列同源性为42.1%~99.4%(表4)。其中,λ系列蛋白核苷酸序列与参考毒株的同源性为42.1%~98.6%,氨基酸序列同源性为15.5%~99.9%,核苷酸同源性最高的分别为广东SH12株(98.0%)、广西GX-Y7株(98.6%)及重庆SL株(98.2%),氨基酸同源性最高的分别为广东SH12株(99.9%)、河南HN5d株(99.7%)及山东DE150株(99.6%)。μ系列蛋白核苷酸序列与参考毒株的同源性为63.5%~98.7%,氨基酸序列同源性为77.2%~99.4%,核苷酸及氨基酸同源性最高的均为重庆SL株,同源性分别为97.7%和99.0%,98.4%和99.4%,98.7%和98.7%。σ系列蛋白核苷酸序列与参考毒株的同源性为21.2%~99.4%,氨基酸序列同源性为23.5%~99.7%,核苷酸及氨基酸同源性最高的分别为北京J18株(98.4%和99.7%)、广西GX-Y7株(99.4%和99.7%)、广东SH12株(97.9%和97.8%)及重庆SL株(99.3%和99.4%)。
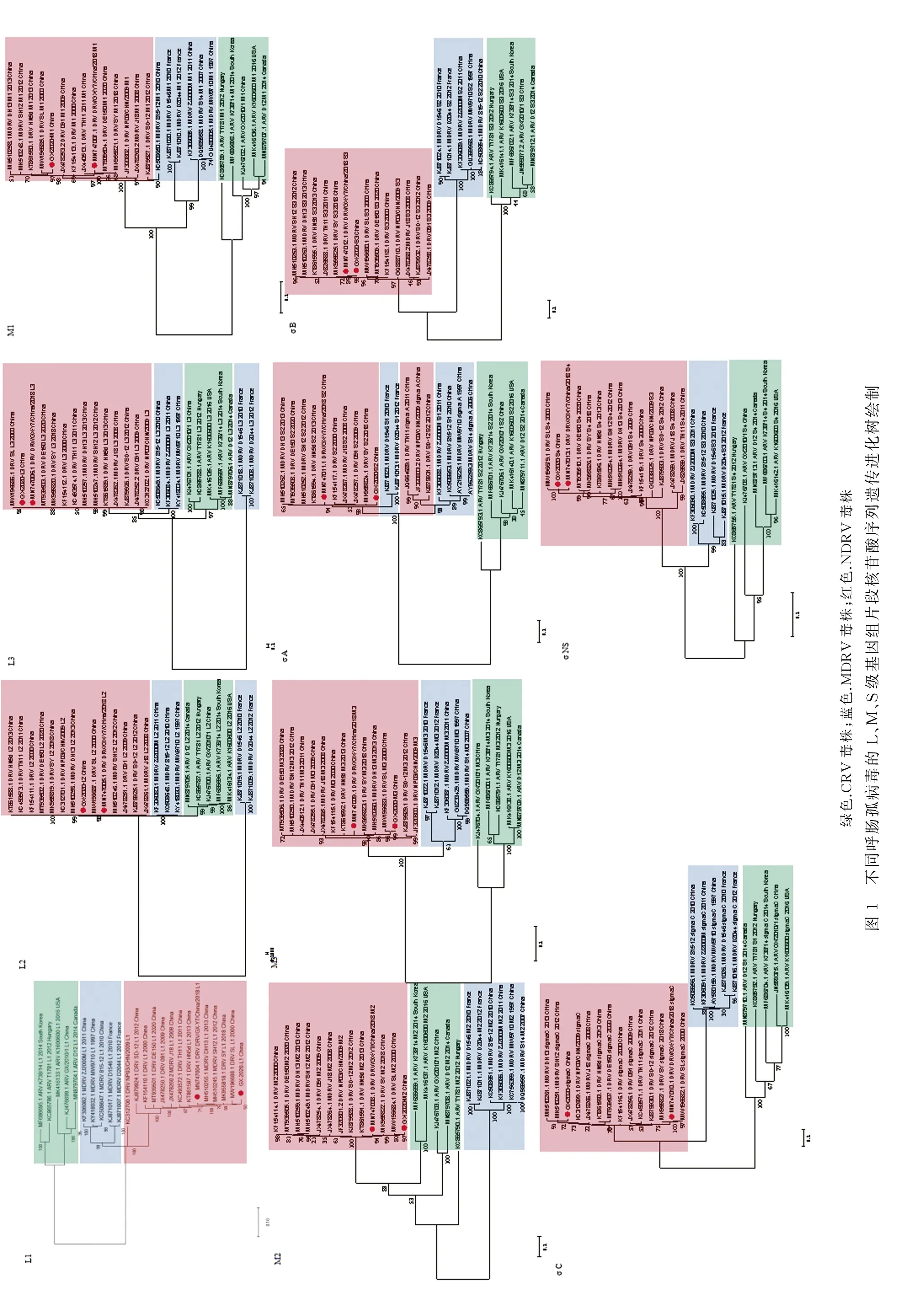
2.3 遗传进化分析对GX01-2020株及参考毒株的10个全基因组片段进行系统发育分析(图1),结果显示,10个片段的核苷酸遗传进化树基本相似,以GX01-2020、广西GX-Y7株与国内外的NDRV代表毒株为一个进化分支,以经典MDRV和CRV为另外两个独立的进化分支。但也有不同,如L2、L3的进化树中,MDRV代表毒株组成两个不同方向的进化分支;σ A的进化树中NDRV分成两个进化分支,其中上海TH11株、福建NP03株及山东SD-12株与国内的MDRV参考毒株组成一个分支,而GX01-2020株、广西GX-Y7株及其他NDRV参考毒株与法国MDRV毒株组成另一个分支。此外,遗传距离也有不同。M2片段进化树中,NDRV与CRV遗传关系较近,而其他片段进化树中NDRV则与MDRV较近;σ A系统发育树中,上海TH11株、福建NP03株及山东SD-12株与国内的MDRV参考株遗传关系较近,而GX01-2020株、广西GX-Y7株等其他NDRV毒株与法国2个MDRV毒株关系较近。
2.4 重组分析应用RDP4和SimPlot软件对GX01-2020株的10个基因组片段与参考毒株的基因组核苷酸序列进行重组分析。结果,SimPlot软件检测到GX01-2020株的L1片段存在重组信号,重组亲本为北京J18株和福建NP03株(图2A)。RDP4软件检测到北京J18株的L1片段和广东SH12株的L2片段有重组信号,重组亲本分别为广东SH12株与福建NP03株,山东SD-12株与广东DH13株,但SimPlot未检测出重组的具体位点。此外,浙江ZJ2000M株的S1片段在RDP4和SimPlot中均有重组信号,重组亲本为北京815-12株与法国D1546株(图2B)。
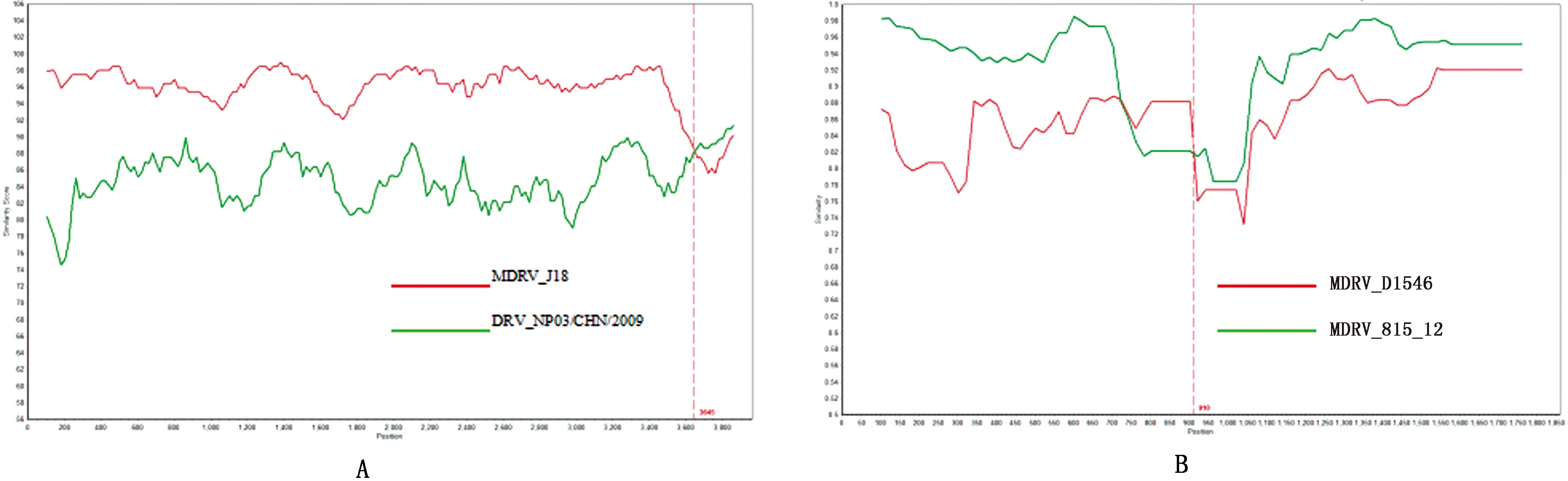
A.GX01-2020,MDRV_J18和DRV_NP03/CHN/2009的L1片段重组分析;B.MDRV_ZJ2000M,MDRV_815-12和MDRV_D1546的S1片段重组分析
3 讨论
广西属于亚热带气候边境地区,为野鸟迁徙过冬常驻区域,加强边境地区水禽源ARV流行病学调查及分子流行病学研究至关重要。本研究采集的病料源自广西北海的番鸭养殖场,解剖发现典型的病理变化为肝脏肿大坏死、脾脏严重肿大充血,呈“樱桃”状。通过自主设计的特异性引物及探针进行实验室分子诊断,检测结果为NDRV阳性,而经典MDRV、AIV、NDV、DTMUV、MDPV、GPV等病原核酸检测结果均为阴性。经全基因组序列测序发现,GX01-2020株共有10个基因组片段:L1~L3、M1~M3、S1~S4。基因组核苷酸序列分析显示,除S1片段外,其他基因组片段大小同WRV和CRV相似。ORF预测表明GX01-2020株除S1外的片段为单顺反子结构,只编码单个ORF,而S1片段为多顺反子结构,编码3个部分重叠区域的ORF。这与CRV类似,但与WRV不同[16-18]。表明,GX01-2020株为不同于MDRV的NDRV。
核苷酸序列同源性分析结果显示,L级片段中λ A核苷酸序列同源性最高的为广东SH12株,该毒株为ZHANG等[19]2012年从广东省的雏番鸭组织中分离到的NDRV毒株;λ B核苷酸序列同源性最高的是广西GX-Y7株,这是基因库上来源广西地区NDRV毒株;λ C核苷酸同源性最高的为重庆SL株。表明,L基因组片段不完全具有地域遗传特征。本研究中GX01-2020株L组片段核苷酸序列与NDRV代表株的同源性均在92.5%以上,这与部分学者研究结果一致[20-22]。此外,发现GX01-2020株L组片段核苷酸及氨基酸序列同源性最低的均为CRV(法国2株MDRV除外),推测可能的原因是该组片段序列来源于CRV几率较低,主要来源于WRV的重排。M级基因组片段核苷酸同源性最高的均为重庆SL株,为98%左右。此外,μ B核苷酸及氨基酸序列同源性最低的毒株均为经典MDRV,而μ A、μ NS片段的则是CRV,这从遗传进化树上也能得出相似结论。表明,M级基因组序列的来源祖先不同。S级片段编码的σ系列蛋白中,σ A核苷酸同源性最低的为MW9710株(21.2%),这是1997年从福建的番鸭分离到的MDRV,由于该片段不完整,仅有300 bp,故导致同源性低于CRV毒株。σ C蛋白在病毒感染早期阶段起关键作用,调节病毒粒子与宿主细胞间相互作用及诱导产生特异性中和抗体[23-24]。此外,对σ C蛋白的核苷酸序列比对分析,通常是作为ARV毒株鉴定与分类的遗传标签[25-28]。本研究中,GX01-2020株σ C核苷酸序列与NDRV代表株同源性为89.1%~97.9%,远高于MDRV的41.5%~43.0%和CRV的36.2%~39.2%,再次印证GX01-2020株为NDRV毒株。另外,通过核苷酸及氨基酸序列同源性分析发现,同源性最低的毒株均为CRV,表明σ级序列来源WRV的几率要大于CRV。以上结果表明,GX01-2020株全基因序列与NDRV流行毒株有高度的同源性,具有遗传多样性的分子特征。
本研究的系统发育分析结果中,基于10个全基因组片段核苷酸序列的遗传进化树基本按ARV种类分化,即NDRV、经典MDRV与CRV组成3个不同的进化分支。但L2、L3、σ A的进化树中,MDRV的代表毒株形成2个独立的进化分支,而其他片段的进化树中MDRV参考毒株组成一个独立的进化分支。其中,来自法国的MDRV毒株D1546和D2044组成一个分支,来自国内的MDRV毒株组成另一个分支。这与GX01-2020株核苷酸同源性分析结果一致,L2、L3片段与法国MDRV毒株同源性显著低于其他MDRV代表毒株。此外,σ A进化树中,NDRV毒株分成2个进化分支,一支是由TH11、NP03、SD-12与MDRV毒株ZJ2000M、815-12、MW9710组成;另一支是GX01-2020、GX-Y7等NDRV毒株与法国MDRV毒株组成,这与σ A同源性分析结果相符合。另外,在遗传距离方面,NDRV的M2片段与CRV较近,与MDRV较远,这与其他片段不同,表明核苷酸序列来源祖先可能不同。以上结果与其他学者研究结论相似[19,29-31]。最后,CRV参考毒株来源地有加拿大、美国、匈牙利、韩国和中国,10个片段系统发育树中,CRV毒株均在同一个进化分支,表明不同地域的毒株并未呈现不同的进化趋势,参考毒株没有明显的地域性遗传特征。
重组分析发现GX01-2020株与北京J18株的L1片段检测到重组信号,这与WANG 等[30]研究结果一致。ZHANG等[19]研究发现J18株的M2片段,广东SH12株及DH13株的S2片段均存在重组现象,但本研究并未检测到该片段的重组信号。此外,本研究中广东SH12株的L2片段和浙江ZJ2000M株的S1片段均检测到重组信号,其亲本毒株均为同种属的病毒,但ZHANG等[19]并未有相关报道。表明,目前重组只发生在NDRV之间及MDRV内部,并未发现NDRV、MDRV与CRV毒株相互之间有重组现象。但随着病毒自身的不断演化,宿主免疫系统的压力及外部环境因素的影响等综合因素作用下,这种情况也许会发生。因此,需密切关注ARV,尤其是对新型NDRV的分子流行病学研究将为该病毒引发疫病的有效防控奠定基础。
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
中国典型病例大全(2022年7期)2022-04-22
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
环球时报(2021-07-12)2021-07-12
科学导报(2021年29期)2021-06-03
医学信息(2019年4期)2019-10-08
科海故事博览·下旬刊(2019年6期)2019-04-16
烹调知识(2019年3期)2019-03-01
分析化学(2014年4期)2014-10-24
