
土壤酸度及铝胁迫对接种AMF茶苗活性氧代谢的影响
2022-06-16 02:38胡方洁翟秀明杨海滨盛忠雷
南方农业 2022年9期
胡方洁,翟秀明,邓 敏,杨海滨,盛忠雷
(重庆市农业科学院茶叶研究所/重庆市茶叶工程技术研究中心,重庆永川 402160)
茶树[Camellia sinensis(Linn.)O.Kuntze],多年生经济作物,具有一定耐铝聚酸的特性。当前,酸雨影响及化肥施用不合理等原因导致茶园土壤酸化问题日益严重,西南茶区在不同pH 值区段内土壤酸化特征明显,并呈现日趋严峻态势。土壤过度酸化和活性铝富集过量,会加速土壤营养元素流失,降低土壤的盐基饱和度、阳离子交换量,减少茶树根际有益微生物种群数量,抑制土壤中微生物的活性,影响植物根系发育和对养分的吸收。而我国南方红黄壤具有脱硅富铝化成土过程,Al2O3含量高,交换性Al3+占阳离子交换量的20%~80%,pH 值4.5~6.0[1]。酸雨和铝毒被认为是南方地区农作物减产的重要原因之一。目前,学界关于铝胁迫对茶树的影响研究主要集中在茶叶的生长发育、养分变化、铝含量、叶绿素荧光等方面[2-3],抗氧化酶保护系统的酶活性研究有少量报道[4-5],而对于酸铝复合胁迫下茶苗的活性氧代谢研究鲜有报道。
丛枝菌根真菌(Arbuscular mycorrhiza fungi,AMF),可提高植物在酸化、盐碱、洪涝、干旱等胁迫下的抗性,提高植物在逆境下的生存能力[6-7]。摩西球囊霉(Glomus mosseae)是目前国内外研究中采用较多的一种模式菌株。摩西球囊霉提高了拂子矛属红丁香(Calamagrostis villosa)和曲芒发草(Deschampsia flexuosa)耐酸性[8-9]。接种AMF 能活化难溶态磷,提高土壤有效磷的含量,改善紫花苜蓿在酸性土壤中的生存环境[10]。AMF 能够提高植物抗逆性的研究已得到证实,但接种AMF 茶树的抗氧化酶保护系统对酸铝胁迫的响应机制尚不清楚。因此,研究酸铝胁迫对接种AMF 茶苗抗氧化酶系统的影响,探究AMF 摩西球囊霉在茶树酸铝胁迫过程中发挥的作用,以及AMF 摩西球囊霉提高茶植株耐酸铝胁迫的保护机制,可为缓解茶树酸铝胁迫寻找方法,对改善西南茶园土壤酸化问题有重要的理论和现实意义。
1 材料与方法
1.1 试验地点
试验地位于重庆市农业科学院茶叶研究所试验基地(永川区茶山竹海),东经105°53′、北纬29°23′,海拔610 m,气压94 500 Pa,年平均气温17.9 ℃,降雨量1 034.9 mm。
1.2 试验材料
1)供试植株选取生长健壮、长势一致的1年生无性系茶苗,品种为“福鼎大白”。2)AMF 摩西球囊霉,由北京市农林科学院植物营养与资源研究所“丛枝菌根真菌种质资源库”提供。3)盆栽容器为塑料盆,上口内径41 cm、下口内径27 cm、盆深31 cm,基质为6∶4的泥炭土和茶园原土,重量16 kg·盆-1。
1.3 试验方法
1.3.1 酸、铝胁迫试验设置
采取盆栽法,将复合肥和AlCl3·6H2O 混匀拌入基质并装入盆中,设4个土壤酸度胁迫:pH3.5、pH4.0、pH4.5、pH5.0,土壤pH 值调节参考陈雪娇等[11]的方法。每种土壤酸度下再设置5个铝浓度梯度(T1、T2、T3、T4、T5),土壤铝的含量分别为:0、0.06、0.12、0.24、0.48 g·kg-1;空白设置1 个处理:不接种丛枝菌根真菌、不设置酸铝胁迫。共计21个处理,每个处理设置3 个重复,每个重复1 盆,每盆栽种长势一致的茶苗5株,共计63盆。
1.3.2 茶苗叶片抗性指标活性测定
试验于2020年10月进行,定植1个月后在盆栽土壤中接种扩繁的摩西球囊霉,之后1周开始胁迫试验。试验期间,每隔10 d 施加对应的处理液以保持相应酸铝浓度。分别在胁迫后的120 d 时,每盆固定选取当年生芽下第3 片健康成熟叶片进行测定。所有指标测定均重复3次。
过氧化物酶(POD)采用酶标法测定[12-13],每克组织(鲜重)在每毫升反应体系中每分钟A470 变化0.05为1个酶活力单位。
过氧化氢酶(CAT)采用钼酸铵比色法测定[14],以每克组织(鲜重)每分钟催化1 μmol H2O2降解定义为1个酶活力单位。
超氧化物歧化酶(SOD)采用酶标法测定[15],以每克组织(鲜重)在黄嘌呤氧化酶偶联反应体系中抑制百分率为50%时的SOD 酶活力定义为1 个酶活力单位。
1.4 数据处理
试验数据采用Microsoft Excel 2013 软件进行数据统计并作图,利用SPSS 22.0软件进行单因素方差分析(One-way ANOVA),用LSD法进行多重比较。
2 结果与分析
2.1 酸铝胁迫对茶苗POD活性的影响
POD 作为植物抗逆境保护体系中一种适应性酶,在防御细胞膜过氧化、降低细胞受伤害程度中发挥重要作用。在酸铝胁迫下,茶苗叶片中POD 活性存在较大差异。本研究中,空白处理下的茶叶POD 活性为11.06 U·g-1FW,接种过AMF 的茶苗叶片POD 活性均高于空白组。从图1 可以看到随着土壤pH 值从3.5 升高5.0 时,在相同的铝浓度T1 处理下,茶苗叶片中POD 活性呈现出先下降再升高的趋势,其中在pH 值为4.5 时,此时POD 活性显著低于其余三种pH 处理(P<0.05);在铝浓度T2 处理下,POD 活性在土壤pH值为3.5 时最低;在铝浓度T3 处理下,POD 活性在土壤pH值为4.5时最低;在铝浓度T4处理下,POD活性在土壤pH 值为5 时最低;在铝浓度T5 处理下,POD活性随着土壤酸性降低呈现先增大后减小,其中在土壤pH 值为4.5 时最低。而在相同土壤酸度条件下,POD 活性受铝胁迫的程度差异较大。在T1、T2、T3、T4 处理下,POD 活性均远远小于T5 水平,差异显著(P<0.05)。除T1 处理外,T2、T3、T4、T5 处理下POD 活性随着土壤酸性下降均呈现出先略有升高再降低最后大幅升高的趋势。由此说明,酸、铝这两种胁迫因子分别对茶苗叶片POD 活性都有明显影响,其中当土壤铝浓度达到0.48 g·kg-1时,影响达到极大(P<0.05);而酸胁迫在3.5~4.0 的范围时对茶苗叶片POD活性影响最大。

图1 酸铝胁迫对接种AMF茶苗叶片过氧化物酶(POD)活性的影响
2.2 酸铝胁迫对茶苗CAT活性的影响
CAT是植物体内的一种抗逆境胁迫的氧化还原酶,能维持活性氧代谢的平衡状态。本研究中,空白处理的茶苗叶片 的CAT 活性为16.12 μmol·min-1·g-1FW。从图2 看出接种AMF 后,随着土壤酸性和铝含量的增加,茶苗叶片的CAT 活性发生了明显的变化。其中,在T1 和T4 处理下,当土壤pH 值逐步增大时,CAT 活性表现出逐步降低然后再升高的趋势;而T2 和T3 处理下,CAT 活性表现的规律呈现相反的趋势;而在T5处理下,CAT活性则呈现先升高后降低的趋势。从图2得出,pH 值在3.5 和4.0 时,在最高浓度的铝胁迫下CAT 活性达到最大,显著高于其余的4 个处理(P<0.05);pH 值在4.5 时,T3 处理下的CAT 活性达到最大,显著高于另外4 个处理(P<0.05);pH 值在5.0时,T1 处理下的CAT 活性达到最大。从图2 还可以得知,当土壤pH 值在4.0且土壤铝含量在0.48 g·kg-1时,CAT 活性最大,显著高于其余处理(P<0.05);而在当土壤pH 值在5.0 且土壤铝含量在0.24 g·kg-1时,CAT 活性最小,显著低于其余处理(P<0.05)。综上可见,在土壤酸性较强的情况下,土壤铝含量高低显著影响了茶苗叶片的CAT 活性,表现为高浓度的铝胁迫显著提高了茶苗叶片的CAT 活性;在土壤酸性较弱的情况下,高浓度的铝胁迫对茶苗叶片的CAT 活性影响程度低于低浓度铝胁迫。
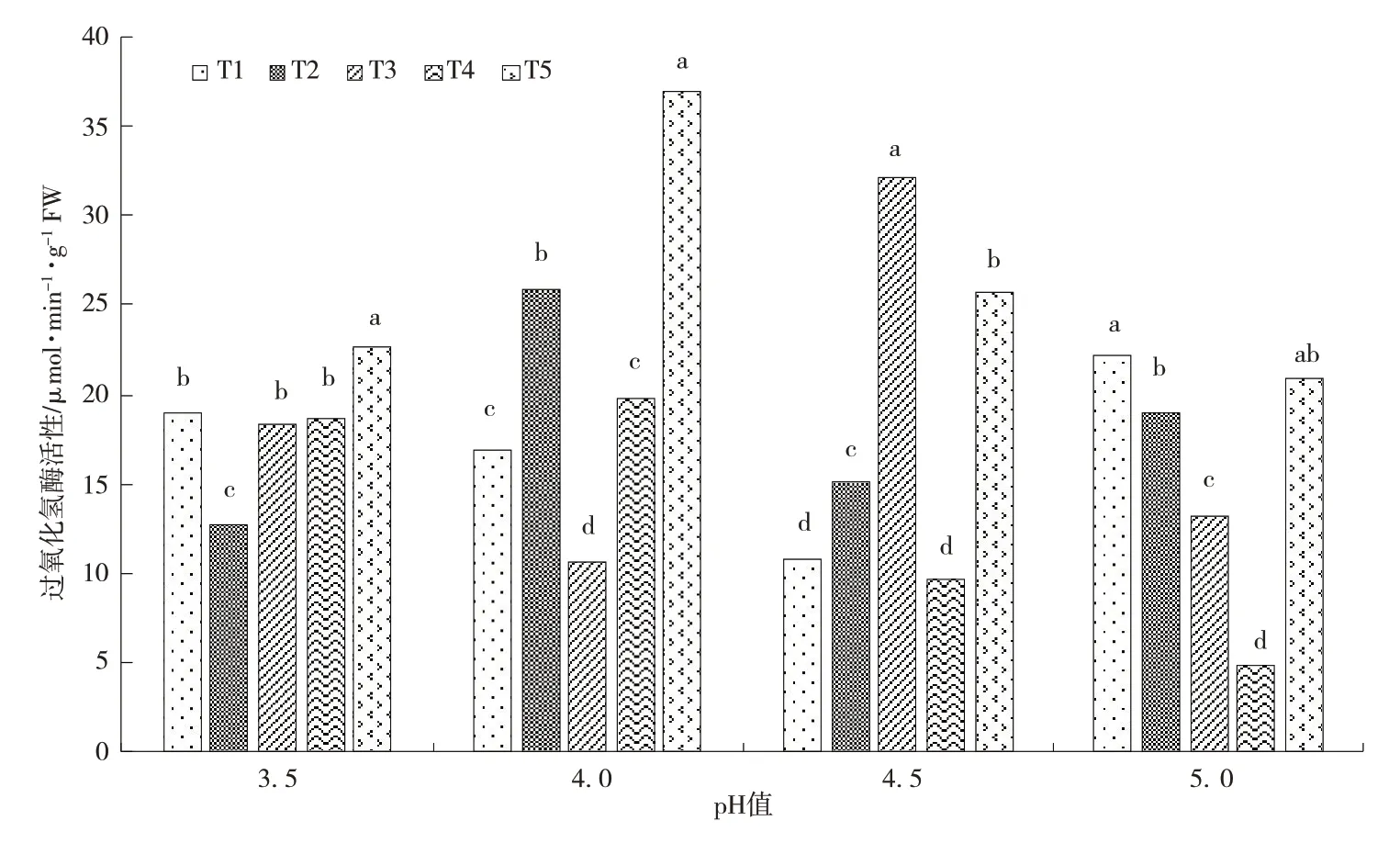
图2 酸铝胁迫对接种AMF茶苗叶片过氧化氢酶(CAT)活性的影响
2.3 酸铝胁迫对茶苗SOD活性的影响
逆境中,SOD 可以清除植物受胁迫产生的多余O2·-,并将活性氧歧化反应成H2O 和O2来维持细胞体正常的生理[16]。本研究中空白处理茶叶SOD 活性值为151.13 U·g-1FW,均低于接种AMF 处理。从图3 得知接种AMF 后,土壤的pH 值和铝含量对茶苗茶叶的SOD 活性有着十分重要的影响。在T1 和T2 处理下,SOD 活性随着土壤酸性的降低呈现先下降再逐步升高的趋势;在T3处理下,SOD 活性在土壤pH 值为4.0时达到最大;在T4 处理下,SOD 活性在土壤pH 值为4.5时达到最大;在T5 处理下,SOD 活性在土壤pH 值为3.5 时达到最大。而在不同土壤pH 值影响下,SOD 活性均随着土壤铝浓度的增加呈现增大趋势,其中pH值为3.5、4.0、5.0 时,SOD 活性均在最高浓度铝胁迫下达到最大,显著高于另外4 个处理(P<0.05)。综上可见,铝胁迫显著影响了茶叶的SOD 活性(P<0.05),随着土壤铝浓度增高茶苗叶片SOD活性随之增大的趋势;而酸胁迫对茶叶的SOD 活性有一定程度的影响,但影响趋势规律不明显。
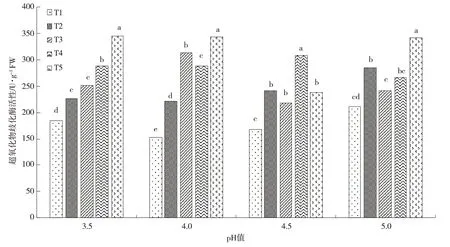
图3 酸铝胁迫对接种AMF茶苗叶片超氧化物歧化酶(SOD)活性的影响
3 结论与讨论
抗氧化酶保护系统由SOD、CAT 和POD 等抗氧化酶组成[17]。在逆境下,植物会通过调动抗氧化酶保护系统来对抗胁迫,它们在植物体内协同作用下清除体内的超氧阴离子自由基、羟自由基和过氧化氢来减少活性氧累积,防止膜质过氧化,减轻细胞膜的损伤,降低了细胞内膜系统的破坏程度,其活性的高低可以衡量植物对胁迫的耐受程度[18-20]。相关研究表明,植物SOD 在抵御单一金属胁迫时能起到一定的保护作用,表现出随着金属浓度的提高,SOD 活性呈先升后降趋势[21-22]。本研究中,pH 值为4.5 时,茶苗叶片SOD 活性随着土壤铝含量的增加呈先升后降趋势,这与曹林等[16]、李西等[23]研究趋势相似。当pH 值为3.5、4.0、5.0 时,此时茶苗叶片SOD 活性随着土壤铝含量的增加几乎呈一直增大趋势,出现这种趋势的原因可能是本研究中的铝胁迫浓度还远未达到茶苗的最大耐受性。罗亮等[4]研究发现低浓度的铝处理(10 mg·L-1)促进茶树POD、CAT 活性增加,而高浓度的铝处理(100 mg·L-1)降低POD、CAT 活性。本研究中,不同pH 值下茶苗POD 活性均在土壤铝含量为0.48 g·kg-1时达到最大值,原因可能是丛枝菌根真菌减弱了茶苗受到酸铝2 个因子胁迫的程度,进而导致POD 活性得到进一步提升,在同一水平的酸胁迫下茶苗叶片POD 活性随铝浓度增大呈现先降低再升高再降低的趋势,这与叶锦培等[5]的研究结果十分相似;Cakmak 等发现,大豆根中的CAT 活性在铝胁迫下呈下降趋势[17],而叶锦培等发现随铝浓度增大,茶苗叶片CAT 活性表现为先降低再升高再降低再升高[5]。本研究中,在pH 值为3.5 和4.0 较强酸性的胁迫下,茶苗叶片CAT 活性在铝胁迫增大下呈先增大的趋势,而且效果明显;在pH值为4.5 时,茶苗叶片CAT 活性在铝胁迫下的变化趋势与叶锦培等的研究结果一致。丛枝菌根真菌与部分植物构成互惠共生体系,促进了宿主植物在逆境中的存活与生长,提高了植物的抗胁迫能力。覃圣峰等的研究结果显示,AMF 能促进玉米在酸性土壤中生长,增强玉米耐铝性[24]。张爽等认为在一定的铝胁迫条件下,接种丛枝菌根真菌能提高植物的抗铝毒能力,还能缓解和改善过酸对植物的毒害,这可能是丛枝菌根真菌利用菌根侵染结构丰度的改变忍耐酸铝毒害[25]。李信茹等的研究发现,接种丛枝菌根真菌后,水稻叶片SOD、POD、CAT 活性均升高[26]。沈亚琴等[27]的研究结果也证实,丛枝菌根真菌能有效增加水稻叶片SOD、POD、CAT 的活性,进而提高植物抗氧化酶系活性来缓解重金属毒害作用,增强植物抗逆性。从本研究的结果来看,在茶苗接种AMF后,茶苗叶片抗氧化酶保护系统的SOD 和POD 活性相比对照得到不同程度的提高;多数处理中,CAT 活性也较对照有明显提高,在较强酸胁迫时茶苗叶片SOD、CAT 和POD 活性比较弱酸胁迫时高,在较高浓度或高浓度铝胁迫时茶苗叶片的SOD、CAT 和POD 活性也比低浓度时高,这表明接种AMF后在一定程度上能提高茶苗的酸、铝耐受性。
接种AMF 后茶苗对逆境胁迫的耐受性有一定提高。本文仅对抗氧化酶保护系统的部分酶活性进行研究分析,下一步将对接种AMF 茶苗的生长特征如株高、地径、叶面积、生物量、叶绿素荧光及光合作用等方面进行分析,以期更深刻、更全面地阐述AMF提高茶苗耐受酸铝胁迫的机制。
猜你喜欢
辽宁林业科技(2022年2期)2022-11-24
中草药(2022年21期)2022-11-05
西南农业学报(2022年8期)2022-10-28
作物学报(2022年12期)2022-10-14
亚热带资源与环境学报(2022年2期)2022-06-30
中国食用菌(2022年5期)2022-06-08
土壤学报(2021年3期)2021-07-22
河南农业大学学报(2021年2期)2021-04-27
科学导报(2019年5期)2019-06-11
农村百事通(2018年10期)2018-07-30
