
日照对安康飞播油松成苗效果的影响
2022-06-09 14:42童开林韩崇选李建春
西北林学院学报 2022年3期
童开林,韩崇选,李建春
(1.杨凌职业技术学院,陕西 杨陵 712100;2.西北农林科技大学 林学院,陕西 杨陵 712100;3.陕西省飞机播种造林工作站,陕西 西安 710082)
日照是地球生物圈热量的主要来源,也是影响气候变化的主要动因[1]。随着全球气候变量,环境污染加重,我国日照时数有缩短的趋势,而温度有所上升,降水相对减少[2-5]。气候因子年间和季节变化加剧,高温、冰冻、霜害、干旱、暴雨、冰雹和风灾等极端气象灾害交替发生,严重威胁粮食安全和生态文明建设[6-7]。由于飞播造林的特殊性,对气象因素反应更灵敏,更易受到气候变化的影响。大范围持续性的高温干旱直接导致飞播后种子发芽迟缓,甚至不发芽,增加了被鸟兽取食的风险,使成苗量减少,林地幼苗生存空间收缩,飞播造林成效降低[8-9],甚至影响林木的分布和生长[10-11]。为此,在安康飞播油松与降水量和温度关系研究的基础上,利用1997年安康多效抗旱驱鼠剂(RPA)拌种和对照飞播区油松成苗期的有苗样方频度和成苗量资料[12],分析了日照指标与飞播油松造林成效的相关性及其季节动态,并通过与降水量和温度研究结果对比分析[13],探讨了日照、温度和降水对成苗效果影响的差异。以期为制定最佳飞播期和合理评估RPA拌种飞播效果提供科学依据。
1 材料与方法
1.1 研究地概况
按照1996年安康市飞播造林作业设计方案[14],选择安康市汉滨区油松播区为研究基地。播区地处北亚热带北缘的秦巴山地丘陵沟壑区,主要分为川道、丘陵和山地三大自然地貌属北亚热带大陆湿润性季风气候。年均温度15.7(15.4~16.1)℃;平均年降水量796.6 mm,约60%分布在7-9月,汛期5-10月,占全年降水量的82.2%,其他月份平均降水量都在60 mm以下,其中1月降水量不足5 mm;最少年降水量540.3 mm(1966年),最大年降水量为1 109.2 mm(1983年)。无霜期263 d。年日照时数1 811.5 h,≥10 ℃年积温为4 000~4 400 ℃,历时220 d[15-16]。区内水热条件随海拔高度的变化,植物群落垂直变化明显,属亚热带与温带植物混交类型。亚热带植物主要有茶树(Camelliasinensis)、柑桔(Citrusreticulata)、柚(Citrusmaxima)、橙(Citrusaurantium)、油桐(Verniciafordii)、油茶(Camelliaoleifera)、乌桕(Triadicasebifera)和棕榈(Trachycarpusfortunei)等。毛白杨(Populustomentosa)、旱柳(Salixmatsudana)和榆树(Ulmuspumila)等温带植物分布很广。
1.2 调查方法与数据处理
1997年飞播后,选择RPA拌种与不拌种播区,每年10月按照“M”或“Z”形取样方法,随机抽取50块1 m×2 m样方,调查样方油松株数。重复5次,连续调查6 a。日照资料来源于1997-2004年陕西省统计年鉴。按单位面积成苗量(survival seedling quantity,Qss)和有苗样方频度(frequentness of sample plots with available seedling,F)表述飞播后油松成苗效果(表1)[12]。采用相关分析法和逐步回归模型分析日照指标与油松成苗效果的关系,以模型参数、相关系数和指标量综合分析各日照指标对成苗效果作用稳定性和影响强度,并通过降水量和温度与成苗效果关系研究结果对比分析[13],探讨日照温度降水因子对飞播油松成苗效果作用变化。
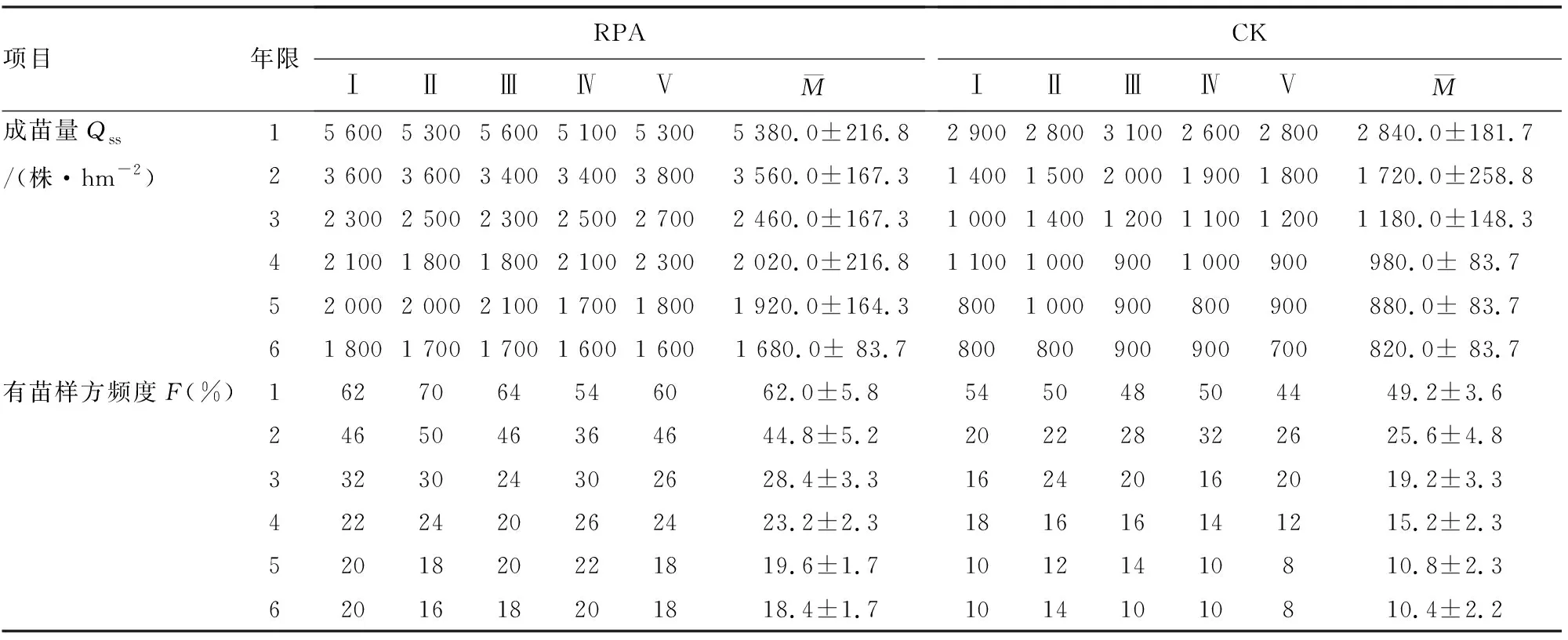
表1 安康飞播油松成苗效果调查(1998-2004年)
2 结果与分析
2.1 成苗效果与日照时数相关分析
年日照时数与飞播成苗效果呈一定正相关性,但相关不显著(r>0,P>0.05)。成苗效果与1月、2月、6月和8月日照时数相关度较低(P>0.05)。RPA成苗量与5月和7月呈显著负相关(r<0,P<0.05),与3月负相关极显著(r<0,P<0.01),而与10月和12月呈显著正相关(r>0,P<0.05),与4月、9月和11月正相关极显著;RPA有苗样方频度与5月和7月呈显著负相关(r<0,P<0.05),与3月负相关极显著;而与4月、9月、11月和12月呈极显著正相关。对照成苗量与7月呈显著负相关,与3月和5月负相关极显著,而与10月和12月呈显著正相关,与4月、9月和11月正相关极显著;有苗样方与5月和7月呈显著负相关,与4月负相关极显著,而与10月呈显著正相关,与4月、9月和11月正相关极显著(表2)。
2.2 成苗效果与日照指标的逐步回归模型分析
逐步回归模型分析显示,9月和7月日照时数(D)对安康飞播油松成苗效果影响较大,首先进入成苗效果模型。接着4月、12月和11月日照时数依次进入RPA成苗量模型,年日照时数和5月日照时数进入RPA有苗样方模型,2月和8月日照时数进入对照有苗样方模型(图1)。
Qssck=-1 038.078+17.434Dsm9
(n=30,R2=0.792,rm9=0.890,F=106.604,P=0.000)
Qssck=4 874.631+26.466Dsm9+11.305Dsm7
(n=30,R2=0.945,rm7=-0.426,F=231.862,P=0.000)
Qssrpa=-1 587.956+31.597Dsm9
(n=30,R2=0.802,rm9=0.896,F=113.414,P=0.000)
Qssrpa=-8 910.358+48.833Dsm9+21.577Dsm7
(n=30,R2=0.974,rm7=-0.415,F=502.350,P=0.000)
Qssrpa=-9 487.963+54.666Dsm9+25.586Dsm7-6.738Dsm4
(n=30,R2=0.979,rm4=0.718,F=408.504,P=0.000)
Qssrpa=-10 857.176+68.362Dsm9+34.867Dsm7-18.265Dm4-8.153 3Dsm12
(n=30,R2=0.983,rm12=0.451,F=358.547,P=0.000)
Qssrpa=-32 401.081+232.406Dsm9+149.518Dsm7-106.570Dm4-98.825Dsm12-49.565Dsm11
(n=30,R2=0.993,rm11=0.640,F=325.393,P=0.000)
Fck=-23.132+0.320Dsm9
(n=30,R2=0.748,rm9=0.865,F=83.083,P=0.000)
Fck=-99.203+0.499Dsm9+0.278Dsm7
(n=30,R2=0.916,rm7=-0.394,F=147.497,P=0.000)
Fck=-101.257+0.523Dsm9+0.278Dsm7-0.118Dsm2
(n=30,R2=0.936,rm2=-0.145,F=126.798,P=0.000)
Fck=-108.466+0.519Dsm9+0.352Dsm7-0.536Dsm2+0.204Dsm8
(n=30,R2=0.955,rm2=0.027,F=133.436,P=0.000)
Frpa=-20.153+0.378Dsm9
(n=30,R2=0.746,rm9=0.864,F=82.093,P=0.000)
Frpa=-112.322+0.595Dsm9+0.272Dsm7
(n=30,R2=0.923,rm7=-0.386,F=32.771,P=0.000)
Frpa=75.346+1.310Dsm9+1.217Dsm7-0.287Dsa
(n=30,R2=0.949,ra=0.347,F=162.655,P=0.000)
Frpa=132.575+1.479Dsm9+1.481Dsm7-0.361Dsa-0.061Dsm5
(n=30,R2=0.957,rm5=-0.402,F=139.238,P=0.000)
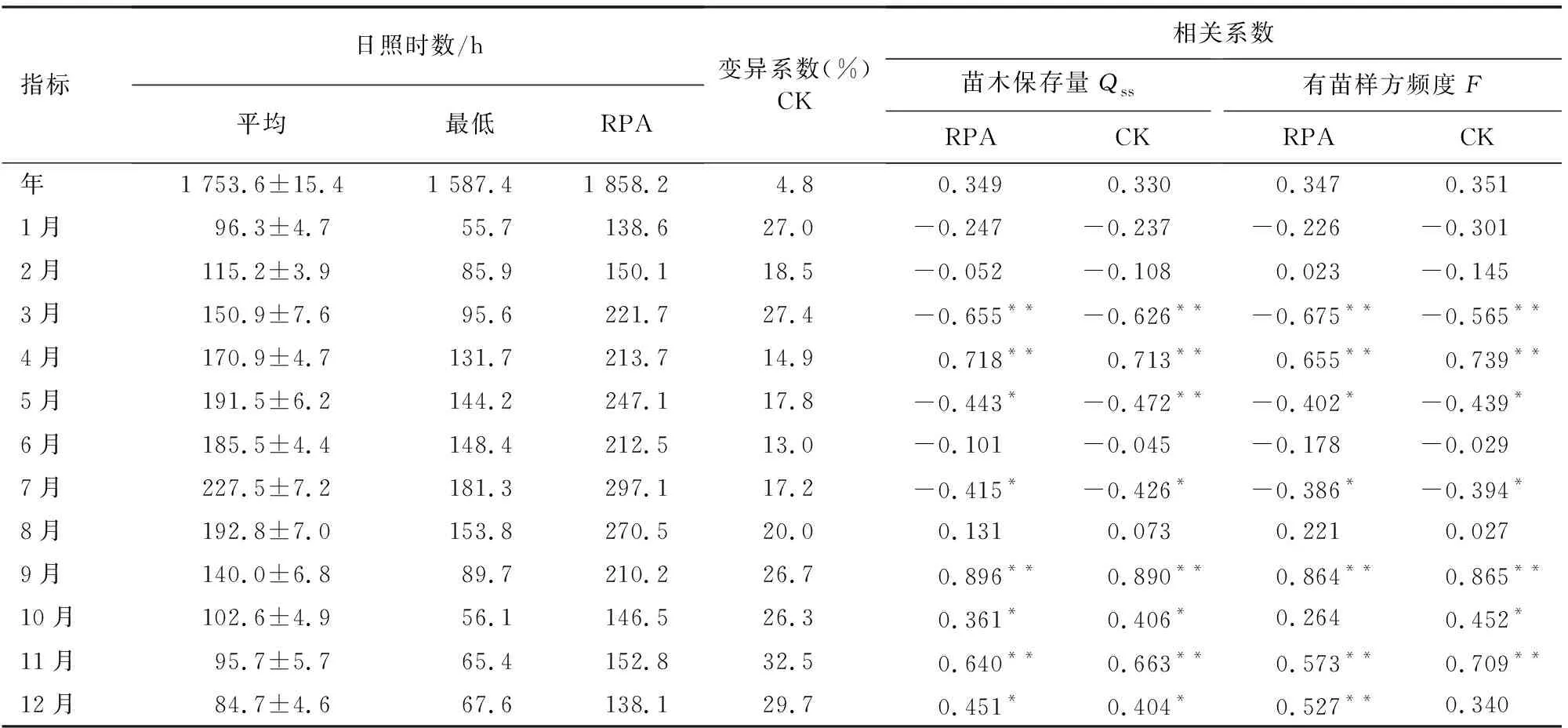
表2 1998-2004年安康日照时数变异与油松成苗效果相关分析
从成苗量模型分析,9月日照时数与成苗量呈极显著正相关(r=0.896、0.890,P=0.000);RPA和对照模型在38.8 h/M附近交汇,斜率比值1.8,在当地9月日照时数范围内{89.7 h/M,210.2 h/M},RPA模型值大于对照,差距有随日照时数延长扩大趋势。在控制9月日照作用下,7月日照与成苗量线性关系极为密切(r=0.932、0.858,P=0.000),对RPA和对照成苗量贡献率为41.8%和41.0%,低于9月日照时数的58.2%和59.0%。排除9月和7月日照因子干扰,4月日照时数与RPA成苗量呈显著负相关(r=-0.454,P=0.015);3个日照因子共同作用下,9月日照对成苗量影响最大,贡献率为52.%;4月影响最小,贡献率仅为7.9%。控制上述日照因子对成苗量影响,12月日照时数与成苗量负相关显著(r=-0.419,P=0.030);4个日照因子配合下,9月和7月日照对成苗量作用相对较大,贡献率为44.9%和37.2%,12月日照作用最小,贡献率仅为3.2%。在5个日照因子协同作用下,7月日照对成苗量影响略大于9月,贡献率为34.8%和33.2%,而4月和12月作用相对较小,贡献率分别为18.6%和8.6%;11月日照作用最小,贡献率仅为4.8%。
与成苗量一样,9月日照时数对有苗样方频度也起着关键作用,两者呈极显著正相关(r=0.864、0.865,P=0.000),模型在-51.4 h/M附近相交,斜率比值1.2;RPA和对照模型值变化规律与成苗量相同。在控制9月日照时数作用下,7月日照与有苗样方也呈极显著正相关(r=0.835、0.817,P=0.000),对RPA和对照模型贡献率为42.6%和42.2%,小于9月的57.4%和57.8%。在排除9月和7月日照干扰下,年日照时数与RPA有苗样方呈极显著负相关(r=-0.587,P=0.001),2月日照时数与对照有苗样方也呈极显著负相关(r=-0.487,P=0.009),但2月日照时数复相关系数仅为-0.145,与对照有苗样方关系比较疏远,作用效果不稳定。3个日照因素作用下,年日照时数对RPA成苗量作用最大,贡献率为52.2%,9月作用最小,贡献率为19.0%;而9月和7月日照对对照有苗样方影响较大,贡献率分别为48.8%和42.1%,2月作用最小,贡献率仅为9.1%。在4个日照因子相互作用下,去除其他因子干扰,5月日照与RPA呈显著负相关(r=-0.388,P=0.045),8月日照与对照正相关极显著(r=0.548,P=0.003),但8月日照时数复相关系数仅为0.027,与对照有苗样方关系极为松散,作用极为不稳定。年日照时数对RPA贡献率为53.2%,远大于7月和9月的28.3%和17.4%,而5月日照作用极小,贡献率仅为1.0%。7月和9月日照对对照有苗样方作用效果稳定,贡献率分别为31.5%和28.6%;而2月和8月日照时数作用效果不稳定,影响相对较小,贡献率为24.3%和15.5%。
引入时间变量,对照油松成苗量与日照因子模型关系不变;而RPA成苗量与飞播年限和5月、4月、9月日照时数关系密切,有苗样方与飞播年限和5月、9月日照时数相关度强,对照有苗样方与飞播年限和6月、7月日照时数关系紧密,分别进入模型(图2)。
Qssrpa=5 222.667-681.714t
(n=30,R2=0.803,rt=-0.890,F=114.255,P=0.000)
Qssrpa=8 146.164-667.246t-15.533Dsm5
(n=30,R2=0.964,rm5=-0.443,F=361.105,P=0.000)
Qssrpa=5 357.781-537.334t-10.001Dsm5+9.101Dsm9
(n=30,R2=0.979,rm9=0.896,F=396.016,P=0.000)
Qssrpa=4 526.205-493.817t-10.917Dsm5+7.556Dsm9+6.269Dsm4
(n=30,R2=0.985,rm4=0.718,F=419.941,P=0.000)
Fck=45.973-6.926t
(n=30,R2=0.752,rt=-0.867,F=84.856,P=0.000)
Fck=2.094-8.646t+0.269Dsm6
(n=30,R2=0.925,rm6=-0.029,F=166.728,P=0.000)
Fck=22.641-8.235t+0.231Dsm6-0.065Dsm7
(n=30,R2=0.955,rm7=-0.397,F=185.144,P=0.000)
Frpa=62.613-8.537t
(n=30,R2=0.820,rt=-0.905,F=127.307,P=0.000)
Frpa=95.119-8.376t-0.173Dsm5
(n=30,R2=0.949,rm5=-0.402,F=251.575,P=0.000)
Frpa=69.985-7.205t-0.123Dsm5+0.082Dsm9
(n=30,R2=0.957,rm9=0.864,F=191.961,P=0.000)
从RPA成苗量模型分析,飞播年限与成苗量呈极显著负相关(r=-0.896,P=0.000)。在消除飞播年限干扰下,5月日照时数与成苗量负相关极显著(r=-0.904,P=0.000),对成苗量贡献率为59.8%,大于飞播年限贡献的40.2%。排除飞播年限和5月日照影响,9月日照与成苗量呈极显著正相关(r=0.637,P=0.000),对模型值贡献率为26.5%,小于5月日照和飞播年限贡献的39.9%和33.6%。在控制上述因子作用下,4月日照与成苗量也呈极显著正相关(r=0.561,P=0.002);4个因子共同作用下,5月日照时数作用最大,贡献率为36.7%,9月和4月日照作用相对较小,贡献率为18.6%和18.8%。
从有苗样方模型分析,飞播年限与其也呈极显著负相关(r=-0.905、-0.867,P=0.000)。去除飞播年限影响,5月日照时数与RPA负相关极显著(r=-0.847,P=0.000),6月日照时数与对照呈极显著正相关(r=0.836,P=0.000),但5月,尤其是6月日照复相关系数较小,与有苗样方关系相对疏远,作用不稳定;对模型值贡献率为56.9%和65.8%,大于飞播年限贡献的43.1%和34.2%。在3个因子协同作用下,控制模型中其他因子作用下,9月日照与RPA正相关极显著(r=0.390,P=0.004),7月日照与对照负相关极显著(r=-0.635,P=0.000),但7月复相关系数为-0.394,与对照有苗样方关系较松散,作用不稳定;其中,5月日照对RPA有苗样方影响最大,6月对对照作用最大,贡献率分别为41.6%和52.0%;而9月对RPA影响最小,7月对对照作用最弱,贡献率为20.3%和18.0%。
以飞播年限权重分析,凸显了9月和7月日照对油松成苗效果的影响,首先进入成苗效果模型。接着4月和12月日照时数进入成苗量模型,年日照和5月日照时数进入RPA有苗样方模型,2月和8月日照进入对照有苗样方模型,最后3月日照时数进入RPA成苗量模型。
Qssck=-583.101+13.169Dsm9
(n=30,R2=0.519,rm9=0.720,F=30.197,P=0.000)
Qssck=-4 527.655+25.100Dsm9+10.546Dsm7
(n=30,R2=0.893,rm7=-0.107,F=112.709,P=0.000)
Qssck=-5 057.157+29.826Dsm9+14.012Dsm7-5.364Dsm4
(n=30,R2=0.913,rm4=0.536,F=91.417,P=0.000)
Qssck=-6 756.402+44.975Dsm9+23.777Dsm7-16.137Dsm4-9.358Dsm12
(n=30,R2=0.939,rm12=0.545,F=95.987,P=0.000)
Qssrpa=-825.781+24.238Dsm9
(n=30,R2=0.517,rm9=0.719,F=30.004,P=0.000)
Qssrpa=-8 556.797+47.622Dsm9+20.699Dsm7
(n=30,R2=0.940,rm7=-0.080,F=212.340,P=0.000)
Qssrpa=-9 512.068+56.148Dsm9+26.922Dsm7-9.678Dsm4
(n=30,R2=0.960,rm4=0.569,F=206.340,P=0.000)
Qssrpa=-11 238.328+71.538Dsm9+36.843Dsm7-20.621Dm4-9.506Dsm12
(n=30,R2=0.967,rm12=0.599,F=185.867,P=0.000)
Qssrpa=-12 022.210+112.831Dsm9+89.200Dsm7-64.492Dm4-63.461Dsm12-32.075Dsm3
(n=30,R2=0.973,rm3=-0.510,F=174.561,P=0.000)
Fck=-13.473+0.231Dsm9
(n=30,R2=0.435,rm9=0.659,F=21.515,P=0.000)
Fck=-91.053+0.465Dsm9+0.207Dsm7
(n=30,R2=0.830,rm7=-0.051,F=65.856,P=0.000)
Fck=-101.307+0.532Dsm9+0.290Dsm7-0.152Dsm2
(n=30,R2=0.871,rm2=-0.199,F=58.451,P=0.000)
Fck=-108.625+0.520Dsm9+0.352Dsm7-0.536Dsm2+0.204Dsm8
(n=30,R2=0.924,rm8=0.356,F=76.356,P=0.000)
Frpa=-12.324+0.297Dsm9
(n=30,R2=0.469,rm9=0.685,F=24.708,P=0.000)
Frpa=-109.858+0.592Dsm9+0.261Dsm7
(n=30,R2=0.875,rm7=-0.064,F=94.086,P=0.000)
Frpa=106.710+1.382Dsm9+1.332Dsm7-0.325Dsa
(n=30,R2=0.934,ra=0.336,F=122.535,P=0.000)
Frpa=143.765+1.514Dsm9+1.532Dsm7-0.376Dsa-0.062Dsm5
(n=30,R2=0.949,rm5=-0.005,F=116.436,P=0.000)
从成苗量模型分析,飞播年限权重下,9月日照与成苗量仍然呈极显著正相关(r=0.719、0.720,P=0.000),但相关度明显降低;RPA和对照模型交点在21.9 h/m附近,斜率比值1.8;模型变化规律与上述模型分析相同,进一步说明RPA对提高油松成苗量的重要作用。在控制9月日照因子作用下,7月日照与成苗量正相关极显著(r=0.936、0.882,P=0.000),但复相关度明显削弱,仅为-0.080和-0.107,对成苗量作用极不稳定;对模型贡献率为41.4%和40.6%,低于9月日照贡献的58.6%和59.4%;与前面模型分析相同。去除9月和7月日照干扰,4月日照时数与RPA成苗量呈极显著负相关(r=-0.571,P=0.001),与对照呈显著负相关(r=-0.436,P=0.020);其偏相关性有所增强,而复相关性明显降低。3个日照因子相互配合下,9月日照对成苗量影响最大,贡献率为50.4%和59.4%,明显大于7月的39.2%和38.5%;而4月日照影响最小,贡献率仅为10.6%和11.1%。排除9月、7月和4月日照因子对成苗量作用,12月日照时数与RPA成苗量呈显著负相关(r=-0.438,P=0.022),与对照负相关极显著(r=-0.542,P=0.003);从RPA相关度分析,其偏相关度有所降低,而复相关度有所增强;9月和7月日照对成苗量贡献率为45.4%、38.0%和41.3%和35.5%,明显大于4月的16.0%和18.1%;12月日照对RPA和对照成苗量影响最小,贡献率仅为0.7%和5.2%。从RPA成苗量5个因子模型分析,在排除模型中其他因子干扰下,3月日照与成苗量呈显著负相关(r=-0.421,P=0.032);7月日照对成苗量作用大于9月,贡献率分别为42.8%和33.3%,而12月和3月日照影响极小,贡献率均为0.3%。
从有苗样方分析,有苗样方频度与日照指标关系性质保持不变,只是模型参数和相关度发生变化。9月日照与有苗样方线性关系紧密,正相关极显著(r=0.685、0.659,P=0.000),相关度明显下降。RPA和对照模型交点在-17.4 h/M附近,斜率比值1.3,比上述分析结果提高。消除9月日照影响,7月日照时数也与成苗量正相关极显著(r=0.874、0.836,P=0.000);与成苗量变化类似,其偏相关性有所增强,复相关性明显减弱,对模型作用极不稳定。贡献率为41.7%和42.0%,明显低于9月的58.3%和58.0%;结论与前面结果一致。排除9月和7月日照因子干扰,年日照时数与RPA有苗样方负相关极显著(r=-0.688,P=0.000),2月日照与对照负相关极显著(r=-0.491,P=0.009);年日照时数对RPA有苗样方影响最大,贡献率为53.4%,明显大于7月和9月的28.4%和18.2%;而2月日照对对照影响最低,贡献率为11.1%,远低于7月和9月的41.7和47.2%;与前面结果基本一致。在飞播年限权重下,控制以上因子干扰,5月日照与RPA有苗样方呈显著负相关(r=-0.478,P=0.012),8月日照与对照正相关极显著(r=0.643,P=0.000);与前面分析结果相比,5月日照偏相关性均有所增强,而复相关性有所下降,对RPA有苗样方作用极不稳定;而8月日照偏相关性和复相关性均明显提高,对对照有苗样方作用效果稳定性增加。4个因子协同下,年日照时数对RPA模型贡献率为53.5%,远大于7月和9月的28.3%和17.2%,5月贡献最小,仅为1.0%;而7月、9月、2月和8月日照对对照有苗样方贡献率依次为31.5%、28.7%、24.3%和15.5%,与前面结果完全一致。
3 结果与讨论
3.1 成苗效果与日照指标相关性及时空变化
太阳辐射存在着显著的日变化节律和季节变化,从而导致温度也发生相应的变化[8-10],也直接引起大气环流不规则的周期性变动,造成降水量的年间差异和季节变化[11-12]。林木形成了自身应对气候变化的能力,产生了季节性的生长变化,也导致了气候因子对林木作用的时空差异[14-15]。
3.1.1 年日照指标相关性的地域差异 由于温度与日照时数呈正相关,降水量与日照时数呈负相关[7-12]。所以年日照时数和年均温与飞播油松成苗效果呈正相关,而与年降水量呈负相关,其中,与年均温相关极显著,与年降水量除对照有苗样方频度外相关均显著。
3.1.2 月日照时数相关性时空变化 冬季,12月日照时数和均温与成苗效果呈正相关。其中,日照与对照和RPA成苗量相关显著,与RPA有苗样方频度相关极显著,与对照有苗样方频度相关不显著;而均温除与对照有苗样方频度相关显著外,与其余相关均极显著。12月降水量与成苗效果均呈极显著负相关。1月和2月日照与成苗效果呈不显著负相关;1月均温和降水量与成苗效果也呈负相关,而2月均温和降水量呈不显著正相关,但仅1月均温与RPA有苗样方频度相关显著[13]。春季,成苗效果与3月日照和均温负相关极显著,与降水量正相关极显著。与4月日照和均温正相关极显著,与降水量极不相关。与5月日照显著负相关,与均温不显著负相关,与降水量正相关极显著[12-13]。夏季,与6月和7月日照负相关,与8月正相关,其中与7月显著。与6月均温相关极不显著,与7月正相关不显著,8月均温与成苗量正相关显著,与RPA有苗样方频度正相关不显著,与对照有苗样方频度正相关极显著。成苗效果与6月降水量负相关极显著,与7月负相关不显著,与8月正相关不显著[12-13]。秋季,9月和11月日照与成苗效果正相关极显著,与10日照除RPA有苗样方频度外正相关显著。与各月均温正相关极显著,与降水量负相关极显著[12-13]。
3.2 不同日照、温度和降水因子对成苗效果的协同作用
3.2.1 排除时间因子的协同作用 逐步回归模型显示,9月和7月日照时数与成苗效果关系紧密。在控制模型中其他因素干扰下,2月和6月日照也对对照有苗样方频度产生影响,4月、11月和12月日照对RPA成苗量发生作用,年和5月日照与RPA有苗样方频度也有一定的相关性。4月和11月均温对成苗效果作用较大,7月均温与对照成苗量也有一定相关,而年极端最高温度也对对照有苗样方频度发生作用;3月均温对RPA成苗效果也有一定影响[13]。12月、11月和9月降水量与成苗量关系密切,而12月、11月和8月降水量对有苗样方频度协调作用[12]。
3.2.2 时间因子下的协同作用 在时间因子协调下,对照油松成苗量与日照因子模型关系不变;5月、8月和4月日照对RPA成苗量作用较大,6月和7月日照与对照有苗样方频度关系紧密,5月和9月日照对RPA有苗样方频度影响较大。3月、10月和11月均温与对照成苗量关系密切,5月均温、年极端最高温度和4月均温对RPA成苗量有所作用,年极端最高温度和4月均温对对照有苗样方频度影响较大,3月和10月均温与RPA有苗样方频度相关度较强[13]。2月和5月降水对成苗量作用较大,2月降水与对照有苗样方频度相关性强,4月、1月和2月降水对RPA有苗样方频度依次产生一定的作用[14]。
3.2.3 飞播年限权重下的协同作用 在飞播年限权重下,9月和7月日照也与成苗效果关系密切。但在控制其他因素作用下,4月和12月日照与对照成苗量相关,4月、12月和3月日照与RPA成苗量也有一定相关,2月和8月日照与对照有苗样方频度相关性也较强,年日照和5月日照也会对RPA有苗样方频度产生影响。在飞播年限权重下,温度与成苗效果模型关系不变[13],降水与成苗效果模型关系也基本保持不变,只是在对照松成苗量模型中增加了1月降水量指标[12]。
从以上模型综合分析,9月和7月日照时数是影响成苗效果的关键日照因子;其次,4月、6月、12月和2月日照也对对照成苗效果有关,5月、8月、年、4月、12月、11月和3月日照也与RPA成苗效果相关。4月、11月、3月和5月均温是影响对照和RPA成苗效果的共同温度指标,与成苗效果密切相关;其次,7月和10月均温及年极端最高温度对对照也有影响,10月均温和年极端最高温度与RPA也有一定的相关度[13]。12月、2月和11月降水量是影响成苗效果和对照有苗样方频度的主要降水指标,5月、9月和1月降水对成苗量也有所作用,8月降水对对照有苗样方频度作用也较大;而RPA有苗样方频度与12月、4月、11月、8月、1月和2月降水量依次相关[12]。在当地日照、温度和降水因子的协同作用下,RPA飞播油松成苗量和有苗样方频度均优于对照[13],说明使用RPA拌种飞播造林能显著削弱林地水热资源年间和季节剧烈变化对油松成苗的作用,提高幼苗抵御极端气候灾害的能力,扩大幼苗在林地的适应范围。
猜你喜欢
当代水产(2022年6期)2022-06-29
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
安徽林业科技(2021年3期)2021-07-27
——入侵植物响应人为扰动的适应性进化方向探究
发明与创新(2021年2期)2021-01-19
妇女之友(2017年3期)2017-04-20
中国科技纵横(2016年15期)2016-12-29
云南农业(2011年1期)2011-08-15
现代农业科技(2009年19期)2009-03-20
中学英语之友·上(2008年11期)2008-12-08
