
细叶云南松混交林土壤微生物群落功能特性及与林分结构关系的研究
2022-06-09 14:38:36胡靓达李远发周海菊叶绍明喻素芳
西北林学院学报 2022年3期
胡靓达,李远发,周海菊,叶绍明,喻素芳
(广西大学 林学院,广西 南宁 530000)
土壤微生物是森林地下生态系统的重要生物驱动力之一[1],对土壤肥力演变,尤其是养分循环具有重要意义[2]。土壤微生物对所生存的微环境变化十分敏感,因此土壤微生物群落特性常被用作土壤环境质量评价和早期预测的敏感生物指标[3]。土壤微生物多样性是评价微生物群落特征的重要内容,反映土壤微生物功能多样性的指标通常包括土壤微生物活性和微生物群落代谢多样性[4]。土壤微生物在生长代谢活动过程中会释放酶,酶的活性、种类和含量不仅可以反映微生物的代谢活性及其对营养元素的需求状况,还可以反映出土壤中物质循环和能量流动的状态[5],因此土壤酶活性是分析土壤微生物群落功能的另一个潜在指标,结合土壤微生物群落功能多样性和土壤酶活性能更全面地反映整个微生物群落功能特征。
由于林分环境因子复杂,土壤微生物特征的变化受植物与土壤协同作用的影响[6]。目前,针对土壤微生物群落功能与植物群落间关系的研究越来越多[7-11],但对土壤微生物群落特性与林分结构相关关系的研究鲜见报道。林分的水平结构和垂直结构直接决定着林下的光照、降水、温度及林内微环境[12],会对植被生长和恢复产生重要影响,同时林分结构在调节关键生态系统过程方面也发挥着重要的作用[13],深入系统地探讨土壤微生物群落功能特性与林分结构的关系,有助于进一步揭示森林生态系统退化、恢复的机理,从而为土壤质量监测、植被恢复评价和可持续生态系统的建设提供科学依据。
在过去几十年,中国西南喀斯特地区南盘江流域的森林经历了大规模的“拔大毛”式的采伐、耕地侵占、盗伐、割皮、割松脂、放牧等干扰,致使原始森林遭受了严重的破坏,后期通过长时间实施封育措施形成了次生林[14-16]。以细叶云南松为建群种的松栎混交林是南盘江流域亚热带次生林的典型代表,对该区域水土保持、水源涵养和生物多样性保护等方面发挥着关键作用[17]。本研究以3种不同类型的细叶云南松次生林为对象,分析其林分结构特征、土壤酶活性和土壤微生物群落碳源利用特征,较为系统地阐明不同类型松栎混交林的土壤酶活性和微生物功能多样性的变化特征,探索影响土壤微生物群落特征的主要林分结构因子。
1 材料与方法
1.1 研究区概况
细叶云南松(Pinusyunnanensisvar.tenuifolia)仅分布于南盘江下游两岸海拔300~1 600 m的丘陵山地,属于微域分布的植被类型,广西雅长兰科植物国家级自然保护区(106°11′31″-106°27′04″E,24°44′16″-24°53′58″N)西靠南盘江(图1),细叶云南松分布较多且保存相对完整。保护区属于典型的喀斯特岩溶地貌区,湿热同季,年平均气温16.8~20.4 ℃,年均降水量941~1 217 mm,降水偏少且集中分布于5-8月,年均相对湿度80%左右,年均霜日8~9 d,全年日照时数1 200~1 500 h,属于典型的亚热带季风气候区。土壤以红壤、黄壤和石灰岩发育的石灰土为主,土层较薄,通气性及透气性能良好,富含有机质及其他营养物质(N、P、K、Ca等)。保护区内森林格局以常绿和落叶种群混生为主,常见树种有:细叶云南松、栓皮栎(Quercusvariabilis)、青冈栎(Cyclobalanopsisglauca)、铁坚油杉(Keteleeriadavidiana)、水锦树(Wendlandiauvariifolia)、小果珍珠花(Lyoniaovalifoliavar.elliptica)、南烛(Vacciniumbracteatum)、二叶列柃(Euryadistichophylla)、盐肤木(Rhuschinensis)等。广西雅长兰科植物国家级自然保护区也是全国兰科植物种类最丰富、分布最密集的区域。
1.2 样地调查
据广西国有雅长林场记载,南盘江流域细叶云南松原始林在20世纪60~90年代受到严重干扰后,在无人干扰条件下经长时间自我恢复形成的次生林主要以松栎混交林为主[18]。2016年春在广西雅长兰科植物国家级自然保护区内选取3种保存相对完整、树种组成不同的细叶云南松天然次生混交林作为研究对象,分别为细叶云南松和多种阔叶树混交林(PYⅠ)、细叶云南松和栎类混交林(PYⅡ)、细叶云南松和铁坚油杉及栎类混交林(PYⅢ)(表1),在3种混交林中分别沿对角线在上、中、下坡设置3个20 m×30 m样方,采用全站仪内置的“偏心模式”程序精确定位样方内所有活树(乔木、灌木)的相对坐标(x,y,z),并对所有胸径大于5 cm的乔木进行每木检尺,记录所有树木的树种名称、胸径、树高、冠幅和健康状况等,同时,对样方的立地环境[16]进行了详细调查。
1.3 林分结构因子测定
从植物多样性、林分空间结构及非空间结构3个方面对林分因子进行了分析,其中,植物多样性的测度因子主要为物种丰富度指数(T_R)、Shannon-Wiener多样性指数(T_H′)、Simpson优势度指数(T_D)、Pielou均匀度指数(T_J)[19];林分空间结构因子主要有林分平均混交度(M)、角尺度(W)、大小比数(U)、开敞度(Ki)及竞争指数(Ci)[19-20];非空间结构因子主要有林分平均胸径(DBH)、树高(H)、郁闭度(CC)、冠幅(CW)及株数密度(N)。
1.4 土壤样品采集及测定
相关研究发现,不同土层(0~10 cm和10~20 cm)土壤环境存在明显差异,这势必会影响土壤微生物群落的垂直分布[21]。本研究在每个20 m×30 m的样方内,沿双对角线随机设置5个土壤取样点,在移除凋落物后,使用内径为8 cm的土钻分0~10 cm(A层)、10~20 cm(B层)2层取样,然后将同一样方内5个取样点同一土层的土壤样品混合,置于灭菌的自封袋内,采用冰袋保鲜的方式带回实验室。移除植物根系和石块后,将样品过2 mm筛,置于4 ℃冰箱中保存,用于土壤微生物群落特性的测定。
土壤酸性磷酸酶用4-氨基氮替比林液和铁氰化钾比色法测定;土壤脲酶采用苯酚钠比色法测定;土壤过氧化氢酶采用高锰酸钾滴定法测定;土壤蔗糖酶采用3,5二硝基水杨酸比色法测定[22]。
土壤微生物功能多样性用Biolog Eco微平板方法测定,接种操作参考宋贤冲等[8]的方法。将接种好的Biolog Eco微孔板置于25 ℃的恒温培养箱中黑暗培养,每隔24 h利用多功能酶标仪(Synergy H4,美国)测定各个生态板孔在590 nm波长下的吸光度值,连续培养192 h。
通过计算每孔颜色平均变化率(average well-color development,AWCD公式中用AWCD表示)来反映土壤微生物利用单一碳源的能力,计算公式如下。
AWCD=∑(Ci-Ri)/n
(1)
式中:Ci为每个有培养基孔的吸光值,Ri为对照孔的吸光值,n为培养基孔数。
Biolog-Eco板有96孔,3组平行,每组31种碳源加1个空白对照,按照化学基团性质,细分为碳水化合物类、多聚物、氨基酸类、胺类、羧酸类、酚酸类6类碳源。
用培养120 h的吸光值计算土壤微生物群落丰富度指数(M_R)、Shannon-Wiener多样性指数(M_H′)、Simpson优势度指数(M_D)、Pielou均匀度指数(M_J)来表征土壤微生物功能多样性指数[10]。
1.5 数据处理
在SPSS 19.0软件中采用双因素方差分析林分类型和不同土层深度及其交互作用对土壤微生物功能多样性因子的影响。采用单因素方差检验不同林分土壤微生物功能多样性因子差异显著性,并利用LSD法进行多重比较(P<0.05)。上述绘图利用Sigmaplot 10.0软件完成。在CANOCO 4.5软件中运用蒙特卡洛检验方法对影响土壤微生物功能多样性的林分因子进行显著性检验,同时利用冗余分析对单个土壤微生物多样性因子与林分因子的相关性进行分析,并完成绘图。所有数据在进行冗余分析之前均经过对数转换。
2 结果与分析
2.1 林分因子
从乔木层植物多样性指标看,Shannon-Wiener指数、Simpson指数、丰富度和Pielou指数均表现为PYⅡ较高,PYⅢ较低,且PYⅡ与PYⅢ的差异均达到了显著水平(P<0.05)。林分非空间结构因子中,PYⅠ的平均胸径、平均树高和冠幅均低于PYⅡ和PYⅢ,但其郁闭度和株数密度高于PYⅡ和PYⅢ。从空间结构来看,不同林分类型混交度和大小比数排序由大到小为PYⅡ、PYⅠ、PYⅢ,角尺度和竞争指数的排序由大到小为PYⅠ、PYⅡ、PYⅢ,而开敞度的排序由大到小为PYⅡ、PYⅢ、PYⅠ(表1)。
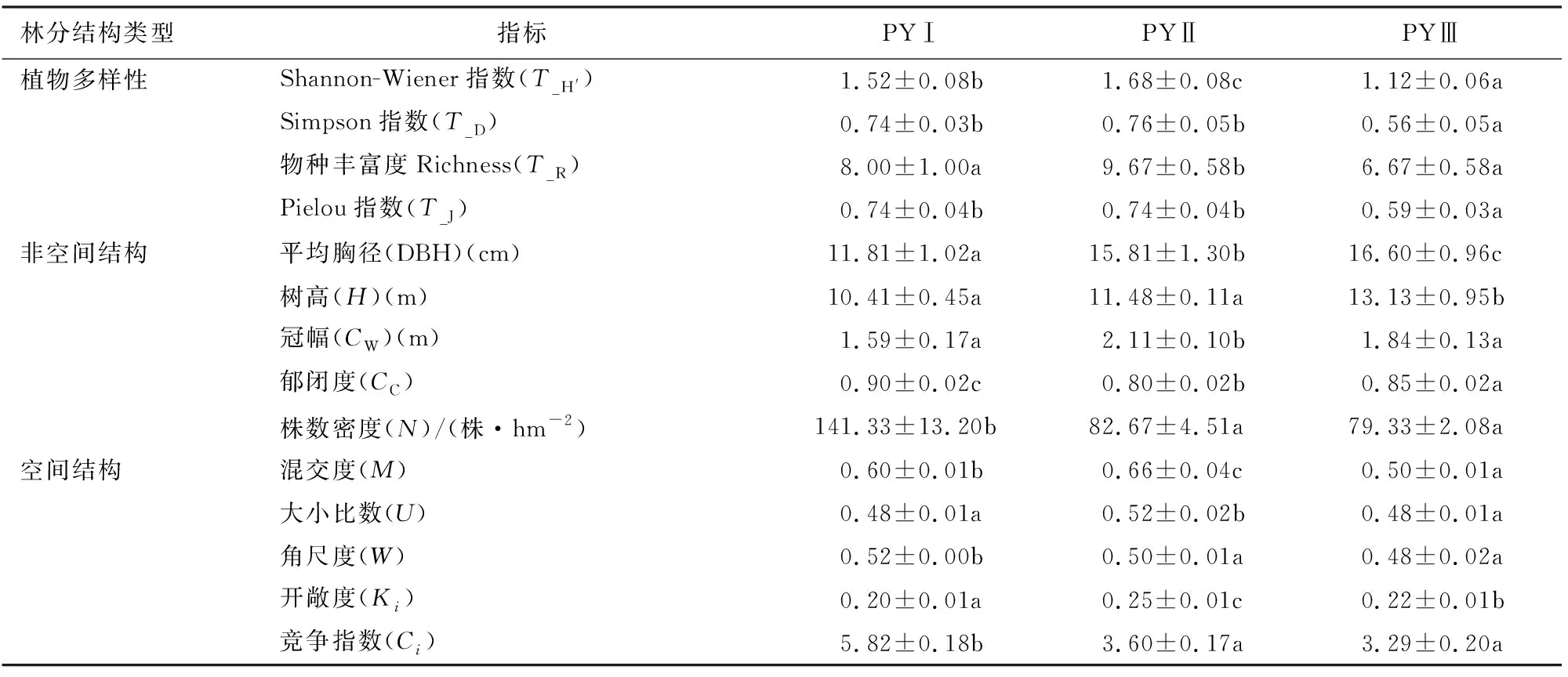
表1 3种针阔混交林的林分结构因子特征
2.2 土壤酶活性
林分类型和土层深度都显著影响了酸性磷酸酶、脲酶、过氧化氢酶和蔗糖酶活性(P<0.05)(表2),但二者的交互作用对4种酶的活性均未产生显著影响。不同类型次生混交林中4种酶的活性均表现出A层显著高于B层。其中,酸性磷酸酶酶活性A、B 2层由大到小均表现为PYⅢ、PYⅠ、PYⅡ;而脲酶、过氧化氢酶、蔗糖酶酶活性A层均由大到小表现为PYⅢ、PYⅡ、PYⅠ,B层由大到小则是PYⅡ、PYⅢ、PYⅠ(表2)。

表2 3种针阔混交林的土壤酶活性方差分析(均值±标准差,n=3)
2.3 土壤微生物群落代谢活性变化特征
不同针阔混交林的AWCD值均随着培养时间的延长而提高(图2),在培养起始的24 h内,土壤微生物代谢非常微弱,AWCD值变化不明显,24 h后进入快速增长期一直延续至120 h时,120 h后增速减缓并逐渐趋于稳定。整个培养过程中,土壤微生物对碳源的利用程度都整体表现为A层高于B层,同一土层不同针阔混交林间土壤微生物对碳源利用程度也存在一定的差异,A层由大到小表现为:PYⅢ、PYⅡ、PYⅠ;B层由大到小则为PYⅡ、PYⅠ、PYⅢ。
2.4 土壤微生物对6类碳源利用特征
林分类型显著影响了土壤微生物对6类碳源的利用,除胺类碳源外,土壤微生物对其他5种碳源的利用也显著受到了土层深度的影响(表3)。6类碳源中,氨基酸、碳水化合物和羧酸类碳源是3种针阔混交林土壤微生物利用的主要碳源(图3),其中对氨基酸和碳水化合物类碳源的利用量A层PYⅢ显著高于PYⅠ和PYⅡ,B层PYⅡ显著高于PYⅠ和PYⅢ;对羧酸类碳源的利用量A层和B层都为PYⅡ较高,PYⅠ较低。从土层深度对比来看,土壤微生物对碳水化合物、氨基酸、羧酸类、多聚物、酚酸等5类碳源的利用量A层均显著高于B层。
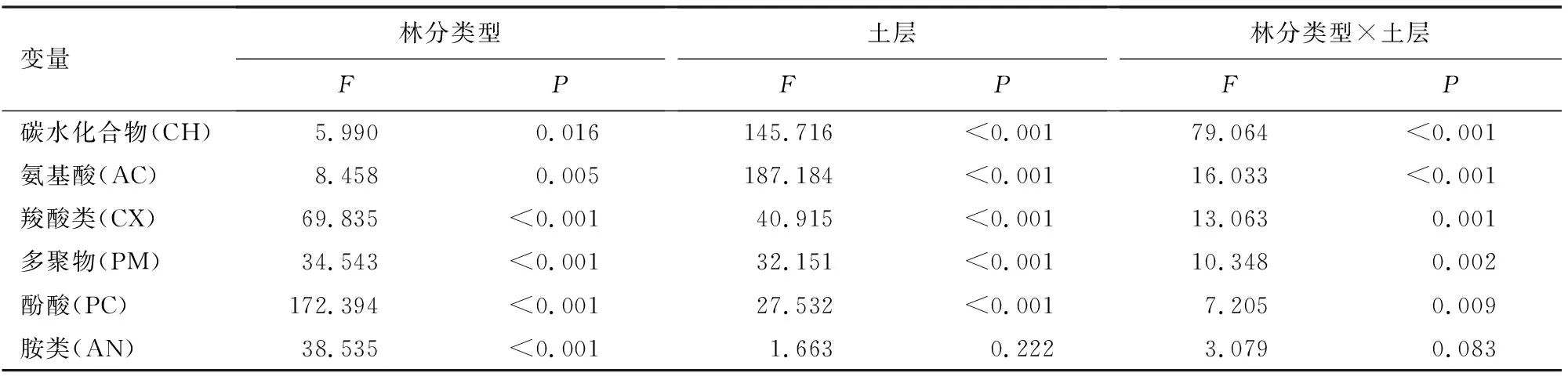
表3 林分类型和土层深度及交互作用对土壤微生物利用碳源的影响
2.5 土壤微生物功能多样性指数特征
林分类型和土层深度均显著影响土壤微生物群落的丰富度指数、Shannon-Wiener指数、Simpson指数和均匀度指数,但二者的交互作用对4个指数无显著影响(表4)。4个指数中,丰富度指数、Shannon-Wiener指数、Simpson指数变化趋势相似,A层显著高于B层,且在两层都表现为:PYⅡ和PYⅢ无显著差异,二者均显著高于PYⅠ。均匀度指数B层显著高于A层,A层PYⅡ显著高于PYⅠ和PYⅢ,B层3种针阔混交林无显著差异(图4)。
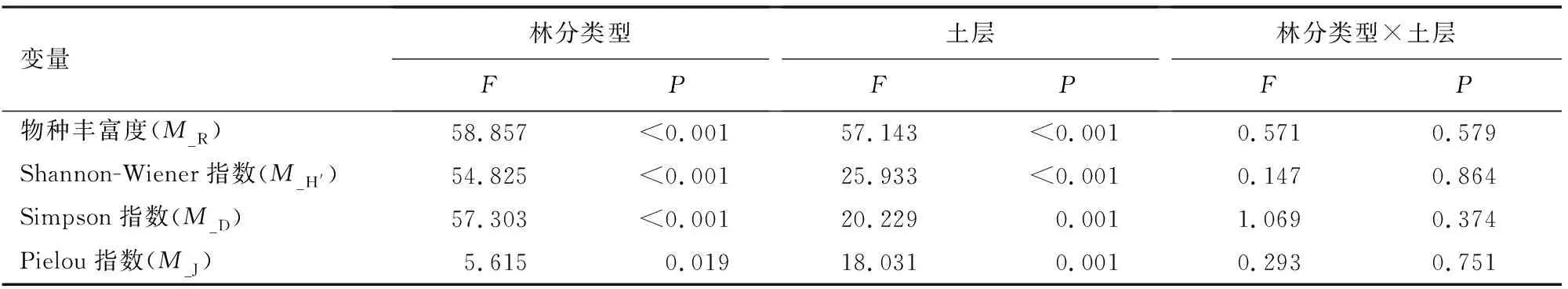
表4 林分类型和土层深度及交互作用对微生物功能多样性影响
2.6 林分结构因子与土壤微生物功能多样性相关关系
利用土壤微生物功能多样性各指标和林分结构因子进行冗余分析,结果显示:A土层第一和第二轴分别能解释土壤微生物功能多样性与林分结构关系变异的62.0%和29.6%,B土层第一和第二轴分别能解释土壤微生物功能多样性与林分结构关系变异的53.6%和32.4%(图5)。Monte Carlo置换检验表明,对A土层土壤微生物功能多样性影响显著的主要林分结构因子有林木株数密度(P=0.002)、林木开敞度(P=0.004)、林木竞争指数(P=0.004)、冠幅(P=0.008)、胸径(P=0.01)、树高(P=0.01)、林木角尺度(P=0.012)、郁闭度(P=0.024);而对B土层土壤微生物功能多样性影响显著的主要为乔木层植被Shannon-Wiener指数(P=0.002)、林木混交度(P=0.002)、林木密度(P=0.004)、乔木层植被Pielou指数(P=0.01)、树高(P=0.014)、乔木层物种丰富度(P=0.014)、郁闭度(P=0.036)、林木大小比数(P=0.038)。
通过土壤微生物功能多样性各指标与单个林分结构因子相关关系的排序可以发现(图5),A土层中,土壤微生物功能均匀度指数、羧酸类碳源的利用量、脲酶活性、过氧化氢酶活性与株数密度、竞争指数、林分平均胸径密切相关;蔗糖酶活性与林分开敞度、平均冠幅、郁闭度密切相关;酚酸碳源利用量与林分平均树高、平均角尺度密切相关。B土层中,多聚物、碳水化合物、胺类碳源的利用量与4个乔木层植被多样性指数均具有密切相关关系;4个微生物功能多样性指数级氨基酸、羧酸类碳源的利用量均与郁闭度密切相关。
3 结论与讨论
PYⅡ林分类型中乔木层植被Shannon-Wiener多样性指数、Simpson优势度指数及Pielou均匀度指数均高于PYⅠ和PYⅢ,这是由于PYⅠ和PYⅢ地势较为平坦,周围人为活动干扰了植物群落以及多样性[17],而PYⅡ位于山顶,无人为干扰,因此植物多样性更高。在林分非空间结构因子中,PYⅠ郁闭度和株数密度较大,导致其平均胸径、平均树高和冠幅均低于PYⅡ和PYⅢ,因为林分密度越大,林木个体之间地下营养以及地上光照竞争等更为激烈,这些竞争关系在一定程度上限制林内树木个体生长,相反地,林分密度小则更有利于林木生长[23]。从空间结构来看,PYⅠ林分林木空间分布格局总体呈团状分布(W>0.517),PYⅡ和PYⅢ总体则呈随机分布(W为0.475~0.517)[24]。3种针阔混交林混交度平均值均以PYⅡ高于PYⅠ和PYⅢ,PYⅠ中由于小果珍珠花株数占比较大,存在本种相邻的现象导致混交度较低;而PYⅢ主要是在经营过程中为了促进铁坚油杉和栓皮栎的更新,出现择伐作业,导致优势树种数量较多而呈现的一些单种聚集分布现象影响了林分整体混交度。3种混交林大小比数平均值差异较小,这是由不同生长状态的林木株数比例较小引起,导致林木之间的竞争不激烈[25],与我们的林木竞争关系分析结果一致(表1)。
3种针阔混交林土壤酶活性、微生物活性和功能多样性都存在差异。研究结果显示,针叶落叶占比较大的PYⅠ林分过氧化氢酶、蔗糖酶和脲酶活性小于其他2种混交类型,因为针叶林有机质通常低于阔叶林[26],为微生物生长提供底物更少[27],该林分中外生菌根类松属植物通过分泌胞外酶获取有机质中的可溶性有机氮[28],致使营腐生的自由微生物可利用底物数量和质量降低[29];其次,PYⅠ的上层郁闭度较大,减少可到达地面的光照和土壤水分,减弱土壤微生物活动[30],从而抑制微生物的生长和繁殖。因此,PYⅠ林分需要通过提高过氧化氢酶、蔗糖酶活性促进土壤有机质分解[31],通过脲酶促进土壤氮素转化[32]。此外,土壤磷酸酶影响土壤碳、氮、磷的转化与循环[32],其活性常与土壤有效P质量分数呈负相关关系[33],PYⅢ土壤酚酸含量显著高于其他2种类型,有利于刺激土壤微生物释放磷酸酶[34],导致该林分类型中磷酸酶活性高于其他2种林分。研究结果还表明土层显著影响土壤酶活性,土壤表层存在较多凋落物,腐殖质含量多,且含有较多的微生物、植物、动物以及有机残体等,较多种类的土壤动物、微生物、植物使得其生理活性增加,释放的酶会增加[35],导致不同针阔混交林中酸性磷酸酶、脲酶、过氧化氢酶和蔗糖酶活性随着土层的加深逐渐降低,这与大多数研究结果一致[36]。
3种针阔混交林土壤微生物利用量较高的碳源类型均是氨基酸和碳水化合物,和针叶类或针阔类树种土壤微生物利用碳源种类的结果一致[37-38]。研究地处于南亚热带,土壤高度风化和酸化[39],硝化作用增强,使土壤中NH4+含量大都远高于NO3-,形成以NH4+占绝对优势的“氮营养生境”[40]。针叶树种长期所处NH4+优势生境将会产生充分适应性,表现出典型的“喜铵性”[40],在喜铵的针叶树种根部含有大量能将游离NH4+转化成氨基酸的酶类,如谷氨酰胺合成酶、谷氨酸合成酶、谷氨酸脱氢酶等[41],这些酶类的存在能使土壤中的NH4+转化成氨基酸[42],从而为微生物提供了大量氨基类碳源,该结果解释了本研究针阔混交林中氨基酸更多被土壤微生物利用。研究结果还发现羧酸类碳源的利用量、脲酶活性、过氧化氢酶活性与林分平均胸径成正相关,即微生物活性越强,越有利于提高林木胸径的增长[43],但当株数密度较大时,胸径大小显著降低[44]。土壤微生物功能多样性同样受株数密度显著影响,株数密度越大,郁闭度越高,微生物功能多样性指数增加[45],但其会降低氨基酸、羧酸类碳源的利用量。由此可见,植被-土壤-微生物三者之间的相互作用,是促进退化森林恢复的重要因素[8]。然而,自然恢复过程中侵入树种杂乱,林分株数密度增大,导致水热以及光照、土壤营养竞争激烈,直接影响或间接通过土壤微生物活动影响树高、胸径和郁闭度等因子[23]。由于树种间竞争并逐步淘汰所需时间长,因此,森林生态恢复过程中仍需采取人工促进措施,对种类组成与密度进行适度调控,改善林分结构及立地环境,以加速恢复退化的森林。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05 07:20:20
今日中国·西班牙文版(2021年4期)2021-04-09 11:22:46
昆钢科技(2021年6期)2021-03-09 06:10:20
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
现代园艺(2018年2期)2018-03-15 08:01:03
广西林业科学(2016年1期)2016-03-20 05:33:02
广西林业科学(2016年4期)2016-03-16 05:44:58
电源技术(2016年9期)2016-02-27 09:05:25
