
乌鲁木齐周边典型荒漠草地群落物种多样性及生物量关系研究
2022-06-02 02:02袁小强张鲜花檀嘉琦韩禧卿
草地学报 2022年5期
袁小强,王 敏,张鲜花,檀嘉琦,袁 惠,韩禧卿
(新疆农业大学草业学院,新疆草地资源与生态实验室,新疆 乌鲁木齐 830052)
草地作为陆地生态系统的重要组成部分,其面积占全球陆地面积的30%[1-2]。我国荒漠草地资源丰富,其面积为4.50×107hm2,占草地总面积的11.47%[3]。在全球气候变化背景下,近十几年来持续干旱、风沙、盐碱化等环境问题愈加严重,造成草地植被覆盖度降低、植物群落结构发生改变、裸地面积增加以及生产力下降,使草地生态系统稳定性和功能性下降[4]。草地退化已成为干旱和半干旱区最为突出的生态环境问题之一[5],极大地影响荒漠草地生态系统的生物多样性、生产力,严重制约着我国北方及其周边地区社会、经济和生态环境的可持续发展[6]。物种多样性可以直接反映植物群落特征及利用现状[7],是生态系统服务功能的的重要指标;生物量则可反映天然草地的健康现状以及生产潜力[8],是生态系统的结构和功能的综合体现,所以探讨物种多样性与生产力之间的关系对生态系统功能的影响具有重要意义。近年来围绕物种多样性与生产力的关系,已经成为生态学研究的热点[9-14]。杜忠毓等[11]研究表明,与对照相比,在养分添加处理下,群落地上生物量与物种多样性之间呈正相关关系,而养分添加则能够缓解水分对荒漠草原植物生长的限制,由此可见,在外界因素干扰下可以改变植物群落地上生物量,进而影响物种多样性与生物量二者之间的关系;朱桂丽等[12]研究发现,高寒草地的经度分布格局对群落特征和生物多样性均有明显的作用,地上生物量和物种丰富度之间存在显著的幂指数关系,二者均受降雨量和平均气温所驱动,研究结果为该地区草地可持续发展提供了数据支撑;新疆温性草甸草原和温性荒漠草原地上生物量与物种多样性呈极显著的线性关系,温性草原则呈单峰型曲线关系,表明水热因子是新疆山地草地物种多样性格局形成的主要决定因素[13],对新疆草地退化防治及其多样性保护具有建设性意义;孙一梅等[14]研究表明,由于试验周期较短,物种多样性无明显变化造成其与生物量之间无显著的相关关系,而极端干旱条件下对草地群落生物量以及物种多样性的影响更需要在大尺度、大环境下进一步探究。以上研究结果出现较大差异,表明研究对象、研究尺度和研究区域不同,使得物种多样性与生物量二者之间关系具有较大的不确定性,但是水热因子是造成物种多样性差异的主要因素[12-13]。
新疆作为我国六大牧区之一,地处欧亚干旱半干旱荒漠区,气候干燥,其中荒漠草地作为新疆主要的冬牧场或春秋牧场,面积为2.69×107hm2,占全疆可利用草地面积的56.04%[15]。乌鲁木齐属于北疆典型荒漠区,地带性自然植被受区域影响主要为温带荒漠植被[16],荒漠草地作为陆地生态系统的重要组成部分在维持和保护生物多样性、养分循环、气候调节、保持水土、防风固沙等方面作用明显[6]。近年来,对乌鲁木齐草地的研究主要集中在人为干扰对群落特征的影响[17-18],但是自然状态下荒漠草地的物种多样性与生物量的关系并未引起足够重视,尚缺乏对自然状态的深入研究。基于此,本研究以乌鲁木齐市周边荒漠草地典型群落为研究对象,通过对草地群落结构和生物量的测定分析,明确群落生物量的分布特征,探讨物种多样性与生物量、地上生物量与地下生物量、凋落物与地下生物量之间的响应关系,研究结果可为退化荒漠草地的恢复及生物多样性保护、生态系统功能维持与恢复提供基础数据和理论依据。
1 材料与方法
1.1 研究区概况
研究区位于新疆维吾尔自治区乌鲁木齐市周边典型荒漠草地区,地理位置为86°37′33″~88°58′24″ E,42°45′32″~45°08′00″ N,海拔423~1 338 m。气候极其干燥,属典型的中温带气候。最热的7月平均气温23.5℃;最冷的1月平均气温-14.6℃;年平均温度5.9℃;无霜期147 d;年均降水量为150~200 mm,年均蒸发量为2 000 mm。土壤基质为沙质、土质、沙砾质、砾石质等,植被以梭梭(Haloxylonammodendron)、膜果麻黄(Ephedraprzewalskii)、霸王(Sarcozygiumxanthoxylon)、驼绒藜(Ceratoideslatens)、伊犁绢蒿(Seriphidiumtransiliense)、小蓬(Nanophytonerinaceum)、角果藜(Ceratocarpusarenarius)、猪毛菜(Salsolaspp.)、叉毛蓬(Petrosimoniasibirica)等为主。
1.2 研究方法
1.2.1野外样地设置及采样 取样点设置:根据20世纪80年代1∶100万的新疆草地类型图并综合考虑乌鲁木齐市周边荒漠草地的分布特点,采用路线调查和典型样地布设相结合的方法,以地理区域、地形地貌及土壤基质为主要因子,将研究区划分为北部、南部、中部及西部的沙质、砾石质、土质及沙砾质荒漠草地,植被类型选择以分布面积较大、自然状态下人类尚未干扰利用、土壤基质不同的梭梭(沙质荒漠)、梭梭+膜果麻黄(砾石质荒漠)、膜果麻黄+霸王(砾石质荒漠)、伊犁绢蒿(土质荒漠)、小蓬群落(沙砾质荒漠)为主的典型群落为研究对象。研究区采样信息见表1。群落中还伴有蛇麻黄(Ephedradistachys)、沙漠绢蒿(Artemisiadesertorum)、囊果苔草(Carexphysodes)、针茅(Stipacapillata)、木本猪毛菜(Salsolaarbuscula)、细枝木蓼(Atraphaxisdecipiens)、琵琶柴(Reaumuriasongoarica)、短叶假木贼(Anabasisbrevifolia)、角果藜(Ceratocarpusarenarius)、木地肤(Kochiaprostrata)等植物。

表1 样点信息统计表Table 1 Statistical table of sample point information
样地设置:对于每个典型群落分别设置5~6个采样点,采样点之间距离为2~3 km,每个采样点选取具有代表性区域、微生境差异较小的地段,布设1个100 m×100 m的典型样地。每个样地内,等距离布设3条样线(100 m),每条样线距离为30 m,每条样线等距离(30 m)布设3个1 m×1 m的草本样方,共计9个;针对梭梭型、梭梭+膜果麻黄型、膜果麻黄+霸王型草地,还要按照十字交叉法再设置5个10 m×10 m的灌木样方。具体见图1。
采样方法:2019年7-8月,分种测定样方内植物自然高度、密度、盖度、地上及地下生物量,同时收集地上凋落物,记录样地周边生境信息。
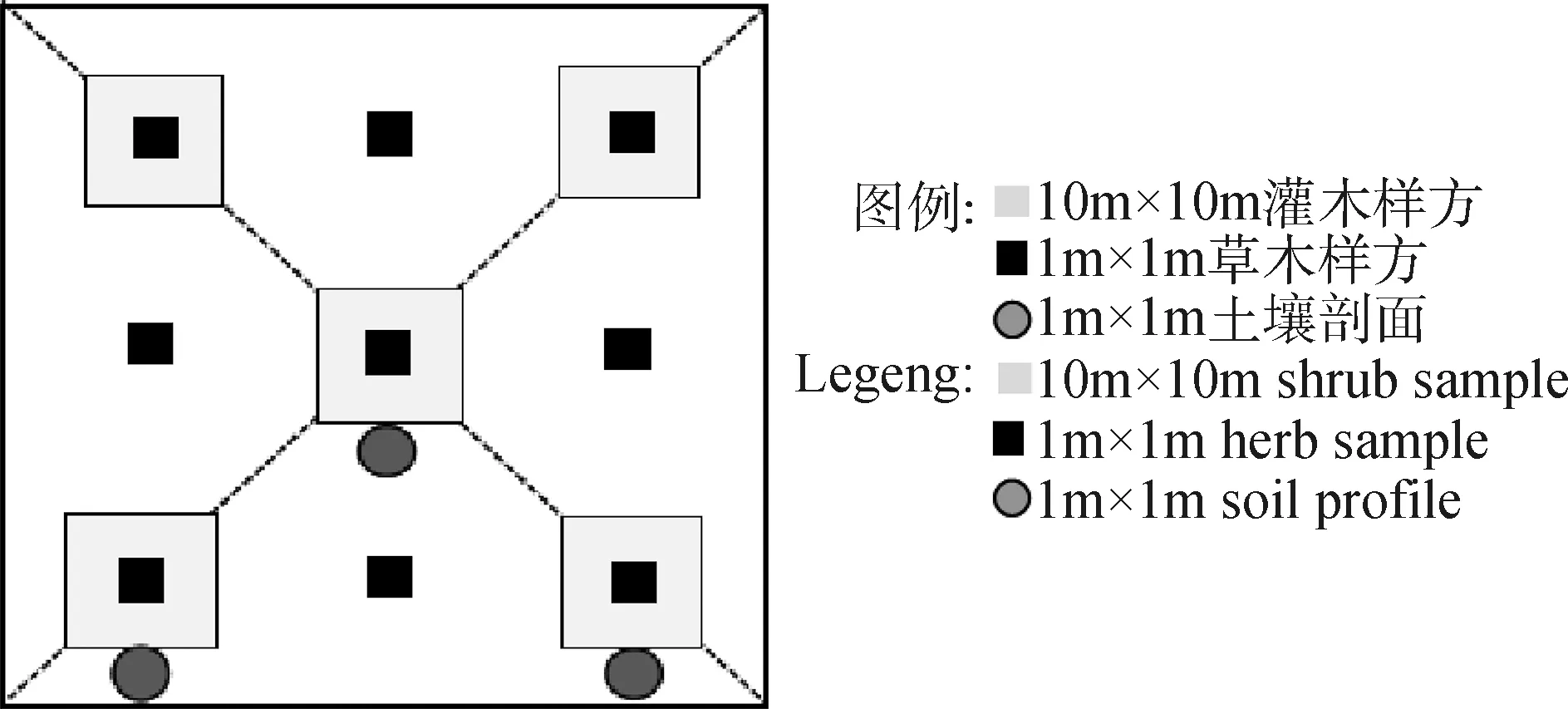
图1 样方布置Fig.1 Sample layout
1.2.2测定指标 高度:每个样方内用卷尺分种测量植株的自然高度。
盖度:草本样方采用针刺法,即样方中该植物的基盖度(针刺到该物种的次数)占整个样方面积的百分比,灌木样方通过测定植株株丛径计算植被盖度。
密度:采用直接计数法测定密度,即每个样方单位面积内各个物种的株丛数。
地上生物量:对于草本样方则采用将样方内每种植物分别齐地面剪割,称其鲜重,而对于灌木样方则采用标准枝法称其鲜重,装入信封袋标记并带回实验室置于烘箱中105℃杀青30 min后于80℃烘干24 h,称其干重。
地下生物量:在完成草本及灌木地上生物量测定后,采用三角取样法选取样地内的3个样方,在每个样方挖取面积为1 m×1 m的土壤剖面,采用挖土块法,表面积为10 cm×20 cm,按土层深度0~5 cm,5~10 cm,10~20 cm,20~30 cm,30~50 cm对根系分层取样,把碎石、杂物分拣后将根系装在已做好标签的网袋(孔径为0.5 mm)带回实验室后,用自来水冲洗根系表面的泥土,然后在65℃下烘干至恒重(48 h),称其干重。
凋落物生物量:直接收集地上凋落物,并装入信封袋带回室内置于烘箱中80℃烘干24 h,称其干重。
1.2.3数据统计分析
(1)重要值
在对各样地内每种植物的高度、盖度、密度及生物量统计的基础上,分别计算样方内草本与灌木的重要值[19],公式为:

(1)
(2)物种多样性测度
不同群落的物种多样性选用Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(E),计算公式如下:
Shannon-Wiener指数:
(2)
Pi=ni/N
(3)
Simpson指数:
(4)
均匀度指数:
E=H/lnS
(5)
式中S为物种数目;Pii为第i个物种的相对重要值;ni为第i个物种的重要值;N为群落中所有物种重要值之和。
(3)群落相似性
本研究采用Jaccard(1901)相似性系数进行分析测定[20],其计算公式如下。
群落相似性系数:
(6)
式中,a表示群落A、B都有的物种数,b表示群落群落A没有但是B有的物种数,c表示群落A有但是群落B没有的物种数。当相似性系数(Cj)达到0.00~0.25时为极不相似,0.25~0.50时为轻度相似,0.50~0.75时为中等相似,0.75~1时为极相似[21]。
(4)数据处理
利用Microsoft Excel 2013和SPSS 20.0对数据进行统计分析:采用SPSS 20.0多元回归分析对荒漠草地物种多样性与生物量之间的关系进行曲线拟合;采用单因素方差(one-way ANOVA)对不同群落特征和多样性指数进行差异性分析(P<0.05),结果均以均值±标准差表示,采用Origin 2018绘图。
2 结果与分析
2.1 群落数量特征差异
2.1.1主要物种组成及其重要值 由表2可知,梭梭群落共6科7属7种植物,其中藜科植物有2种,占总数的28.57%,梭梭的重要值达到0.773;梭梭+膜果麻黄群落共有6科9属9种植物,其中藜科植物达到了总数的44.44%,其他各科均只有1种,梭梭和膜果麻黄的重要值分别为0.357和0.323;膜果麻黄+霸王群落出现9种植物,隶属于4科9属,其中藜科植物有5种,所占比例超过总数的50%,膜果麻黄和霸王的重要值分别为0.267和0.361,仍为主要优势种;伊犁绢蒿群落中出现17种植物,隶属于9科17属,其中种数最多的为藜科植物和禾本科植物,分别有6种和5种,占到总数的35.29%和29.41%,伊犁绢蒿的重要值达到0.727;小蓬群落共出现10种植物,隶属于2科10属,其中藜科植物有7种,占到总数的70%,十字花科有3种植物,占总数的30%,小蓬的重要值达到了0.766。由此可知,各群落物种组成单一,7~17种植物不等,但是优势植物的重要值都为群落中最高,其中在西北干旱荒漠区为主的藜科植物占比达到28.57%~70%,占居优势地位。
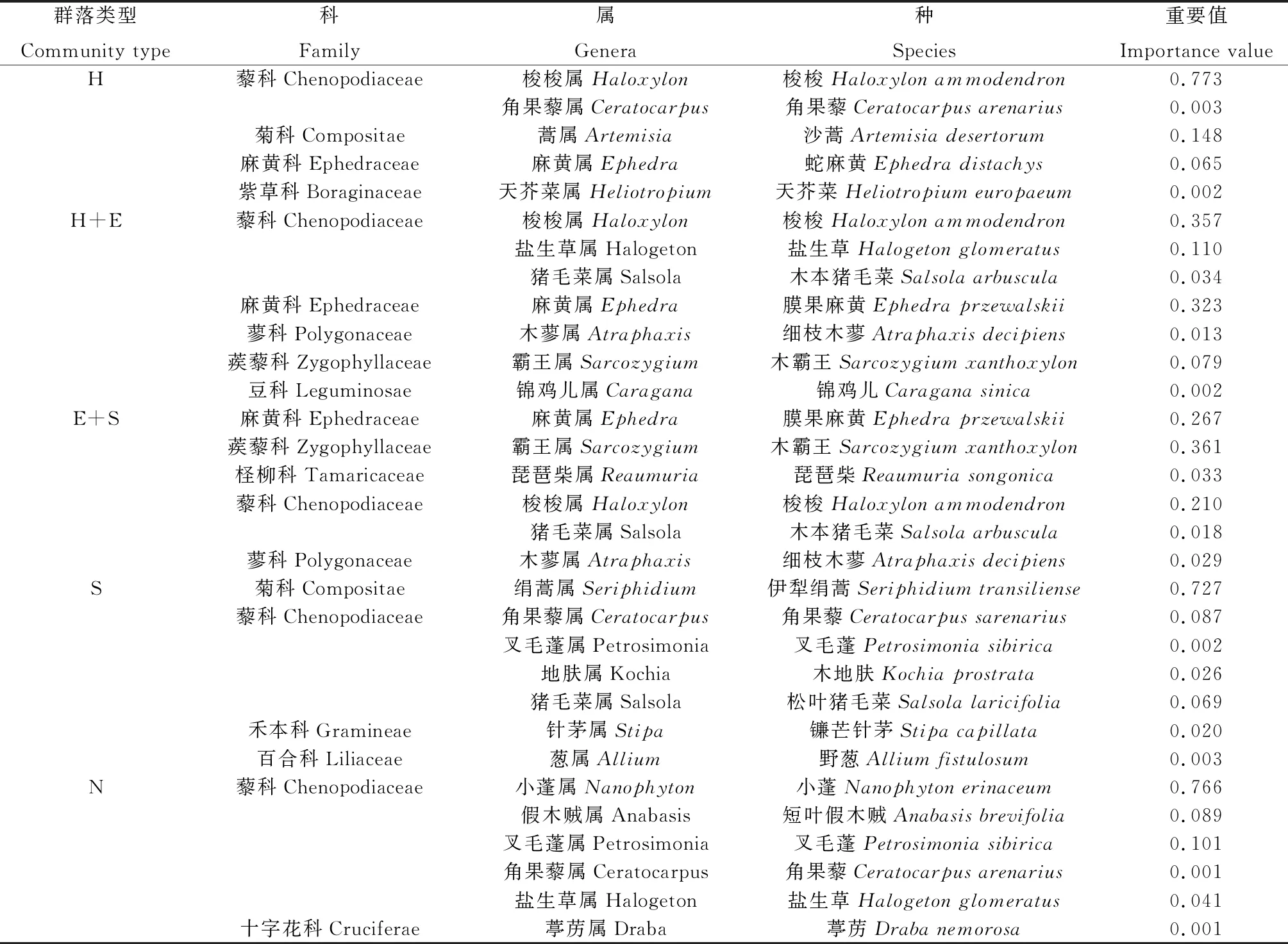
表2 荒漠草地不同群落的主要物种组成及重要值Table 2 Main species composition and importance values of different communities in desert grassland
2.1.2群落高度、盖度及密度差异 由表3可知,各群落间平均高度、盖度及密度呈现出显著性的差异(P<0.05)。其中群落平均高度介于4.28~163.46 cm之间,表现为梭梭群落最高,小蓬群落最低;盖度上表现为伊犁绢蒿群落盖度最高,梭梭群落、小蓬群落、梭梭+膜果麻黄群落次之,膜果麻黄+霸王群落盖度最低;密度方面表现为膜果麻黄+霸王群落密度显著高于其他群落(P<0.05),伊犁绢蒿群落、梭梭+膜果麻黄群落、小蓬群落次之,而梭梭群落的密度最小为8.2株·m-2。

表3 荒漠草地不同群落高度、盖度、密度Table 3 The height,coverage and density of different communities in desert grassland
2.1.3群落地上-地下及凋落物生物量差异 由表4可知,不同群落地上、地下、凋落物生物量分布具有一定规律。综合来看,均表现为地下生物量>地上生物量>凋落物生物量;梭梭、梭梭+膜果麻黄、膜果麻黄+霸王、伊犁绢蒿、小蓬群落地上生物量占总生物量的比例分别为47.30%,14.98%,23.39%,17.65%,23.43%,地下生物量占总生物量的比例分别为50.85%,84.21%,75.50%,75.81%,73.70%,凋落物生物量占总生物量的比例分别为1.85%,0.81%,1.11%,6.54%,2.87%;不同群落地上生物量变化范围在109.59~505.85 g·m-2之间,地下生物量变化范围在344.75~2 276.77 g·m-2之间,凋落物生物量变化范围在13.42~45.51 g·m-2之间;膜果麻黄+霸王、梭梭+膜果麻黄、梭梭群落地上生物量显著高于伊犁绢蒿、小蓬群落(P<0.05),膜果麻黄+霸王、梭梭+膜果麻黄群落地下生物量显著高于梭梭、伊犁绢蒿、小蓬群落(P<0.05),凋落物生物量对不同群落的响应不明显(P>0.05)。

表4 不同群落生物量Table 4 The biomass of different communities
各群落地下生物量随土层变化规律如图2所示,在0~50 cm土层深度中,各群落地下生物量随土层深度变化而表现出不同规律。梭梭+膜果麻黄、膜果麻黄+霸王、伊犁绢蒿、小蓬群落地下生物量在0~5 cm土层中均为最高,显著高于其他土层(P<0.05),并且随着土层深度增加,大致呈降低的趋势;而梭梭群则随着土层深度增加,地下生物量呈现出先增加后降低的趋势,并且各土层间差异不显著(P>0.05)。

图2 群落地下生物量Fig.2 The underground biomass of different communities注:图中不同小写字母表示不同群落之间差异性显著(P<0.05),下同Note:Different lowercase letters indicate significant difference between different communities at the 0.05 level in the figure,the same as below
2.2 群落物种多样性分析
2.2.1物种多样性分析H多样性指数结果表明(表5),各群落H多样性指数介于0.68~1.26之间,小蓬群落与其他群落差异均显著(P<0.05),而其他群落彼此之间均表现为差异不显著(P>0.05)。其中梭梭+膜果麻黄群落H多样性指数最高,伊犁绢蒿群落、梭梭群落、膜果麻黄+霸王群落次之,小蓬群落的H多样性指数最低。
D多样性指数结果表明,各群落D多样性指数介于0.38~0.67之间,小蓬群落与梭梭、梭梭+膜果麻黄、膜果麻黄+霸王、伊犁绢蒿群落均差异显著(P<0.05)。
Pielou均匀度均指数结果表明,群落均匀度范围为0.62~0.91之间,小蓬群落与梭梭、梭梭+膜果麻黄、膜果麻黄+霸王、伊犁绢蒿群落差异均显著(P<0.05),其余各群落之间均未达到显著差异(P>0.05)。

表5 群落物种多样性特征Table 5 Characteristics of community species diversity
2.2.2群落相似性分析 由表6可以看出,乌鲁木齐周边各群落相似性系数介于0.040~0.636之间,均未达到极相似程度,其中梭梭+膜果麻黄群落与膜果麻黄+霸王群落之间的相似性系数最高为0.636,达到中等相似水平,而其他各个群落之间的相似性系数均未达到0.25,表明各群落之间物种数量及生境差异明显。
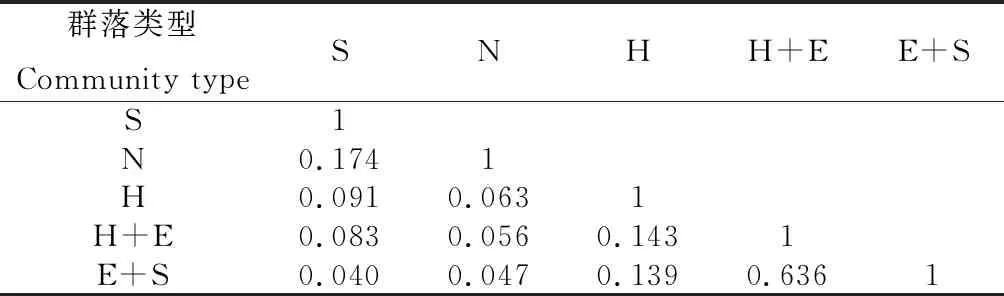
表6 不同群落间相似性系数Table 6 Similarity coefficient of different communities
2.3 群落地上-地下生物量与凋落生物量、物种多样性的模型拟合
为探讨地上生物量、凋落物生物量对地下生物量的响应关系,本研究对各群落的地上、凋落物生物量分别与地下生物量进行曲线拟合。由图3a可知,通过对地上生物量与地下生物量之间建立的最优模型曲线,可以发现二者呈指数函数关系,拟合方程为:y=231.375×e0.003 3x(R2=0.714 8),表明在植物生长过程中,地上生物量处于累积的过程,植物地上生物量对地下生物量贡献较大;由图3b可知,凋落物生物量与地下生物量呈正相关关系,对应的拟合方程为:y=13.818x+259.812(R2=0.616 1),初步表明荒漠草地各群落生长呈异速生长模式。
采用多元回归分析,将地上、地下生物量分别与物种多样性指数进行曲线拟合,并选择R2较大的函数模型建立最优的回归方程。由图3c可知,群落地上生物量与物种多样性之间具有极显著负非线性关系,符合单峰曲线,对应方程为:y=-5.210 4x2+0.004 5x+0.369 9(R2=0.307 1,P<0.01),表明群落物种多样性随地上生物量的增加而表现出先上升后下降的趋势,即植物生长初期,地上生物量对群落物种多样性作用明显,而随着时间推移,到达一定阶段后物种多样性达到饱和,最后呈现出下降的趋势;从图3d可以看出,植物群落地下生物量与物种多样性之间呈显著正相关关系,对应方程为:y=3.073 5x+0.678 2(R2=0.543 4,P<0.05),表明群落地下生物量对群落多样性的影响不受时间尺度的限制,在植物生长周期内,地下生物量对物种多样性作用保持不变。由此说明,植物可以通过内部机理协调运作,控制地上生物量与地下生物量生长速率,进而影响群落物种多样性以应对荒漠复杂的环境变化。

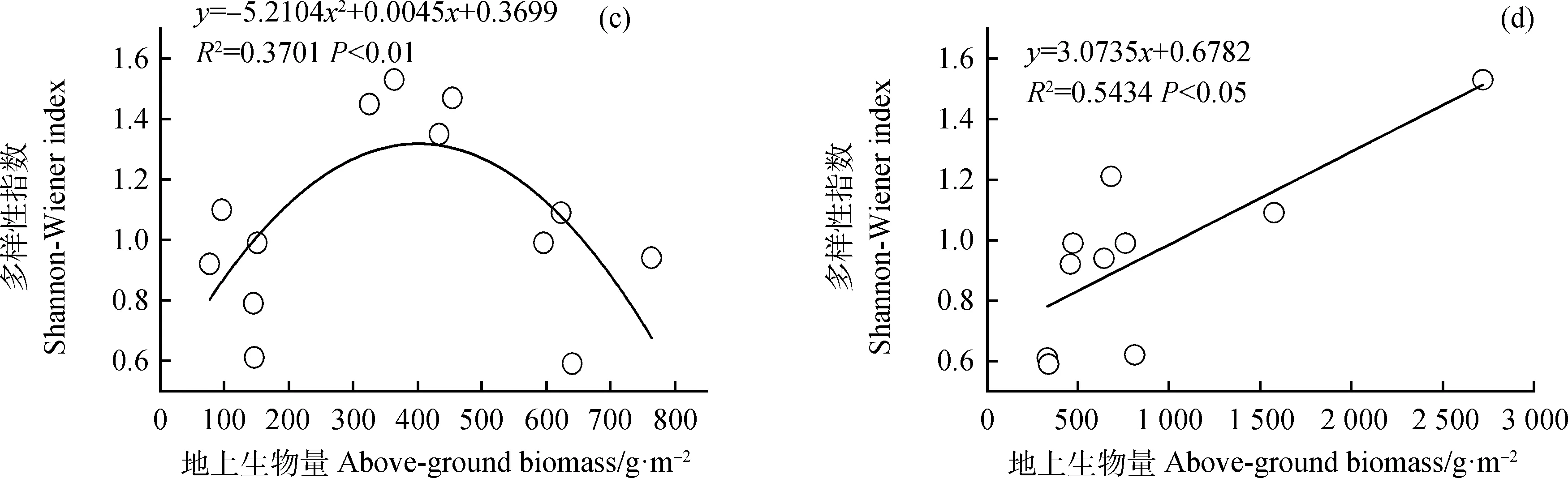
图3 荒漠草地生物量、物种多样性指数的模型拟合Fig.3 Model fitting of biomass and species diversity index in desert grassland
3 讨论
3.1 群落地上-地下生物量、凋落物生物量分析
研究表明乌鲁木齐周边荒漠草地不同群落地上、地下、凋落物生物量分布特征具有一定规律,即表现为地下生物量>地上生物量>凋落物生物量,这与杨阳和刘秉儒[22]对宁夏荒漠草原、崔雨萱等[23]对蒿类荒漠草地的研究结果一致。由于土壤基质、样地所属区域水热条件不同,导致各群落间地上生物量、地下生物量存在显著差异,本研究中各群落地下生物量(图2)0~50 cm土层之间差异显著(P<0.05),伊犁绢蒿、梭梭+膜果麻黄、小蓬、膜果麻黄+霸王群落地下生物量在0~5 cm最高,显著高于其他土层(P<0.05)。0~5 cm和5~10 cm土层分别占到地下生物量的43.52%~53.82%和11.81%~30.49%,并且随着土层深度增加地下生物量逐渐减小,这与胡龙飞等[24]对内蒙古温带荒漠草地、杜美琦等[25]对山区灌草群落的研究结果一致。造成这种差异可能与不同质地土壤的积水和持水能力的差异相关。由于研究区属于干旱半干旱荒漠区,植被覆盖度较低,减少了植被对降水的截留与缓冲,使得有限的降水以地表径流的形式迅速流失,而表层土壤质地变粗,容重增大,导致土壤持水能力减少,土壤渗透率降低[26],所以0~5 cm土层的地下生物量远远高于其他土层。而梭梭群落则刚好相反,即随着土层深度增加,地下生物量呈现出先增加后降低的趋势,其原因一方面可能是梭梭群落生境属于沙漠,常年干旱少雨,表层土壤由于干旱环境使得水分大量蒸发,根系未达到水分供给,而随着土层深度增加,地下水供给充足使得根的分布数量也会增加[27-28];另一方面是因为梭梭垂直根系发达,侧根相对较少,在水平方向上,细根主要集中在0~100 cm土层内,细根生长密度较大[29-30],综合以上两点原因本研究在0~50 cm土层中,梭梭群落地下生物量随着土层深度的增加呈先增加后降低的趋势。
乌鲁木齐周边荒漠草地群落地上与地下生物量的分配关系符合指数函数,其线性方程斜率并不等于1,表现为显著的异速生长关系,这与杨阳和刘秉儒[22]、王晓薇等[31]的研究结果一致。张帆等[32]研究表明,在不同植物功能群地上生物量分配显著不同,且优势种的贡献较大,而地下生物量主要受光合产物的量和光合产物分配机制的影响[33],植物将光合产物中较大的一部分投入到根系的生长,投入到叶片的光合产物较少。本研究中由于各群落物种大不相同,且受不同质地土壤的积水、持水能力以及土壤养分的的影响[26],导致群落地上生物量和地下生物量差异显著;群落物种增多导致覆盖度增加,根据最优分配假说,通过内部调节,植物将分配更多的生物量给能够获取外部资源的营养器官中,从而获得更大的生长速率[34]。
乌鲁木齐周边荒漠草地地表凋落物生物量与地下生物量呈显著的线性关系,初步表明凋落物在植物生长过程中具有重要作用。造成这种结果的原因可能有3方面:一是较多的凋落物输入会提高荒漠草草地生态系统土壤有机质含量,凋落物的分解有利于土壤腐殖质含量增加[35],改良了土壤的理化性质,为植物生长提供了良好的立地条件;二是土壤微生物通过对凋落物的分解[36],为植物生长提供有效养分,充足的养分供应提高了地下生物量,从而促进植物根系生长;三是荒漠草地气候干旱、植被稀疏,由于灌木层的存在,凋落物覆盖度会降低土壤温度,提高土壤含水率,尤其是显著降低表层土壤温度,减少地表蒸发量,以此保证植物地下生物量的积累不受影响[37]。
3.2 群落物种多样性与生物量关系分析
大量的研究表明,物种多样性与生物量之间主要有4种形式,即线性关系[11,13]、单峰关系[22,38]、S型曲线[39]和不相关[15,40]。影响群落物种多样性和生物量之间关系的主要因素包括地形因子、气候因子、土壤因子、水热因子、养分、种间关系等[11,25,27,41-42]。本研究中群落物种多样性与地上生物量之间存在单峰曲线关系,与前人的研究结果一致[22,38],其中单峰曲线在小蓬和伊犁绢蒿群落表现为上升阶段,峰值表现为梭梭+膜果麻黄,下降区域出现在梭梭、膜果麻黄+霸王群落。由于小蓬和伊犁绢蒿群落物种丰富,生态位较窄,导致种间资源竞争并未达到最大,物种多样性很低时,环境资源利用地低,生物量较小;而梭梭+膜果麻黄群落植物密度、盖度相对较低,竞争适度,资源利用率最高,生物量最大;梭梭、膜果麻黄+霸王群落物由于高生产力环境资源异质性下降,导致环境中竞争排除加强,群落物种多样性降低[43]。出现单峰的原因,一方面是由于新疆荒漠生态系统这一特殊的生境下,物种间的竞争更加明显,随着生物量增加到一定程度,群落中一些物种因为竞争而消失,从而物种多样性减少[44],另一方面由于植物种类以不同方式使资源的利用率达到最大化,达到了环境最大容量的限制,物种多样性达到峰值后呈下降趋势[31,38]。
本研究中物种多样性与地下生物量二者之间表现为正相关关系,与杨阳和刘秉儒[22]的研究结果一致。表明地下生物量并不受群落种间竞争以及环境资源限制的影响,出现这种关系的原因:其一是群落地下生物量的变化主要由于群落功能群组成的变化而引起[45],物种多样性一般受到小环境及种间互作的影响[46],小尺度的地质生境会忽略由于尺度增大而导致较强的空间异质性对物种多样性和生物量关系的影响[47],由于研究区处于干旱区,环境相对脆弱,水热条件、资源可利用率等自然条件下,植物群落可以通过物种冗余和功能互补来实现对环境变化的适应[43],从而群落地下生物量越高,种多样性越高;其二可能是受光合产物再分配和土壤微生物作用,植物对光合产物进行重新分配有关,将光合产物较大的部分分配到植物根系,植物通过根系分泌物改变土壤微生物群落,从而影响植物季节性生长[48],进而改变地下生物量,物种多样性升高。
4 结论
综上所述,乌鲁木齐周边荒漠草地群落物种组成相对单一,各群落间植被盖度、高度及密度差异显著;生物量均表现为地下生物量>地上生物量>凋落物生物量,不同群落水平地上-地下生物量符合指数函数拟合,支持异速生长模型,凋落物生物量与地下生物量呈显著的线性关系;物种多样性与地上生物量呈单峰曲线关系,与地下生物量呈指数函数关系。本研究为进一步探究新疆荒漠草地群落物种多样性和生物量之间关系奠定了基础。
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15
哈哈画报(2022年4期)2022-04-19
保健与生活(2022年1期)2022-01-26
儿童故事画报·自然探秘(2021年12期)2021-08-16
文萃报·周二版(2021年3期)2021-04-01
中学生天地(B版)(2019年4期)2019-05-08
扬子江(2018年5期)2018-09-26
儿童故事画报·智力大王(2016年7期)2017-02-08
六盘山(2014年4期)2016-06-24
绿色中国(2015年6期)2015-07-03
