
青藏高原高寒湿地生态系统碳水通量与水分利用效率研究
2022-06-02 01:37王云英裴薇薇郭小伟杜岩功
草地学报 2022年5期
王云英, 裴薇薇, 郭小伟, 杜岩功*
(1.中国科学院西北高原生物研究所, 青海 西宁 810008; 2.中国科学院大学, 北京 100049)
湿地生态系统土壤和植被碳储量丰富,碳密度高[1],高寒湿地是青藏高原高寒生态系统长期适应高寒气候环境的主要植被类型[2]。青藏高原湿地约占中国湿地总面积三分之一[3],对气候变化较为敏感,可作为气候变化的指示器[4]。同时,气候变化对湿地面积和分布也有重要影响[5]。作为青藏高原特殊生态系统类型,高寒湿地海拔高、温度低[6],面积约13.3×106hm2,在全球碳平衡中发挥着重要作用,是青藏高原重要水源涵养地[7]。
高寒湿地碳循环和水循环之间存在密切耦合关系[8],可为环境补充地下水、为野生动物提供栖息地等[9]。较高的水分利用效率表明可在有限时间内获得更多光合产物[10]。2015—2017年三年期间大九湖泥潭湿地生态系统水分利用效率日均值大于0,为3.26 gC·kg-1H2O[11]。季节尺度上,金河湾湿地水分利用效率在冬季呈现最高值,而在夏季呈现最低值,全年均值为4.09 mg·g-1[12]。生态系统碳呼吸是陆地和大气[13]、草地生态系统碳交换[14]过程的主要环节,其包括地上部分植物呼吸和土壤呼吸两部分[15]。土壤呼吸动态变化直接影响着全球碳收支平衡[16]。植物呼吸排放的CO2量约占整个生态系统的碳交换的30%~70%[17]。
作为一种调控因子,气象因子平均气温、年降水量对湿地面积变化、景观格局改变[9]、青藏高原环境变化[18]等有着重要反馈作用。此外,气象因子光合有效辐射、若尔盖湿地生态系统碳吸收主要受光合有效辐射影响[19]。土壤温度是影响青藏高原高寒湿地[20]、三角洲滨海湿地[21]、黄河三角洲芦苇湿地碳通量的主要调控因子[22]。光量子通量密度是影响鄱阳湖南矶湿地净生态系统CO2交换量的主要调控因子[19]。有研究表明,当太阳辐射强度较低时,光合有效辐射的增加会引起CO2净交换量增加[23]。增温促进了河源湿地生态系统呼吸[24],使得小泊湖湿地生态系统整个生长季呼吸速率提升了24.50%[25]。青海湖高寒湿地生态系统水分利用效率主要受降水量变化调控,与温度之间呈现极显著正相关关系[26]。草甸湿地生态系统水分利用效率受到净辐射和空气温度影响[8]。土壤温度由于其自身对环境变化较为敏感,进而作为青海湖湿地生态系统CO2通量的主要环境因子[27]。
目前,大部分研究证明湿地生态系统总体表现为碳汇[28]。碳通量作为高寒湿地生态系统研究重点之一,对于碳通量变化及其调控机制研究涉及的很多,但现阶段对于高寒帕米尔苔草湿地生态系统基于通量变化水分利用效率研究涉及的较少。故本研究借助于2004—2009年间青海海北高寒帕米尔苔草湿地生态系统碳通量数据,计算出生态系统总初级生产力和蒸散量的变化,进而计算出水分利用效率值,探讨高寒草甸湿地生态系统碳通量和水分利用效率变化。并耦合气象因子进一步明晰高寒草甸生态系统碳通量数据和水分利用效率的主要气象调控因子。
1 材料与方法
1.1 研究区概况
本项研究是在中国科学院海北高寒草甸生态系统定位站(海北站,37°36′51.12″N,101°20′13.92″E,海拔3 235 m)附近的典型高寒帕米尔苔草湿地生态系统上进行。海北站地处青藏高原东北隅祁连山北支冷龙岭东段南麓的大通河河谷。区域多年平均气温为-1.7℃,年降水量较高,约580 mm,降水主要集中于暖季的5—9月,占年降水量的80%。湿地植被以适冷、湿生、多年生地下芽、具有发达通气组织的草本植物莎草科、毛茛科植物为主,湿地中央以帕米尔薹草(Carexpamirensis)为优势种,边缘以西藏嵩草(Kobresiatibetica)为优势种。
1.2 研究方法与数据分析的采集
1.2.1生态系统呼吸速率和生态系统CO2碳净交换量(Net ecosystem exchange,NEE)数据和生态系统呼吸速率(Ecosystem respiration,Re) 站点通量数据来自国家生态系统观测研究网络科技资源服务系统(http://rs.cern.ac.cn/),数据时间尺度为2004—2009年。利用通量数据Re、NEE和潜热通量(Latent heat flux,LE)计算生态系统总初级生产力(Gross primary productivity,GPP)和蒸散量[29]。
蒸散量ET计算公式如下:
ET=LE/λ
λ=(2500-2.4Ta)×103
其中λ为蒸发潜热,单位是J·kg-1,式中Ta是空气温度,单位是℃。LE是潜热通量,单位是W·m-2。
总初级生产力GPP的计算公式如下:
GPP=Re-NEE
式中,GPP为植被总初级生产力,单位为gC·m-2;Re为生态系统呼吸量,单位为gC·m-2;NEE为生态系统CO2净交换量,单位为gC·m-2。
水分利用效率的计算公式如下:
式中,WUE为水分利用效率,单位为gC·m-2·mm-1;GPP和ET分别为生态系统总初级生产力和实际蒸散量,单位分别为gC·m-2和mm。
1.2.2气象数据 本研究中气象数据均来自于青海海北高寒草地生态系统国家野外科学观测研究站气象观测站。
1.3 数据处理
数据整理采用Excel,均值、标准误、显著性检验、结构方程模型的计算等采用R语言进行计算处理,进一步结合气象因子结合结构方程模型探讨生态系统CO2碳净交换量、生态系统呼吸速率及水分利用效率变化与气象因子的关系。最初通过选取间接影响达到显著性水平的气象因子,最后参与结构方程模型中的为气象因素:风速、相对湿度、空气温度、降水量和净辐射。比较拟合系数(CFI),Tucker-Lewis指数(TLI)和近似误差均方根(RMSEA)用来模型评估。最大似然法,多被结构方程采用的卡方估计来评估模型卡方值(χ2)和路径系数。方程的拟合是在R中使用lavaan包实现的。运用Origin 2019进行图件的绘制。
2 结果与分析
2.1 湿地生态系统碳水通量和水分利用效率年际变化特征
年际尺度上,三者均呈极显著上升趋势(P<0.01,图1)。NEE年内变化呈“V”型波动式变化趋势,且下降幅度远大于上升幅度生态系统呼吸速率和水分利用效率变化呈倒“V”型变化趋势。6年来,NEE,Re和WUE均值分别为0.33 gC·m-2·d-1,1.68 gC·m-2·d-1,0.52 gC·m-2·m-1。
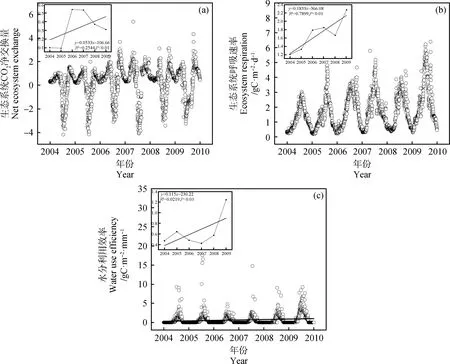
图1 CO2通量、生态系统呼吸速率和水分利用效率年际变化Fig.1 Interannual variation of ecosystem net exchange of CO2,ecosystem respiration rate and water use efficiency
2.2 湿地生态系统碳水通量和水分利用效率季度变化特征
季节尺度上,连续6年间非生长季生态系统碳排放总量大于生长季碳吸收总量(图2a)。生态系统呼吸速率和水分利用效率均呈现出生长季值大于非生长季值(图2b;图2c)。整体而言,生长季NEE,Re和WUE与非生长季变化呈现出较大差异(图2d)。
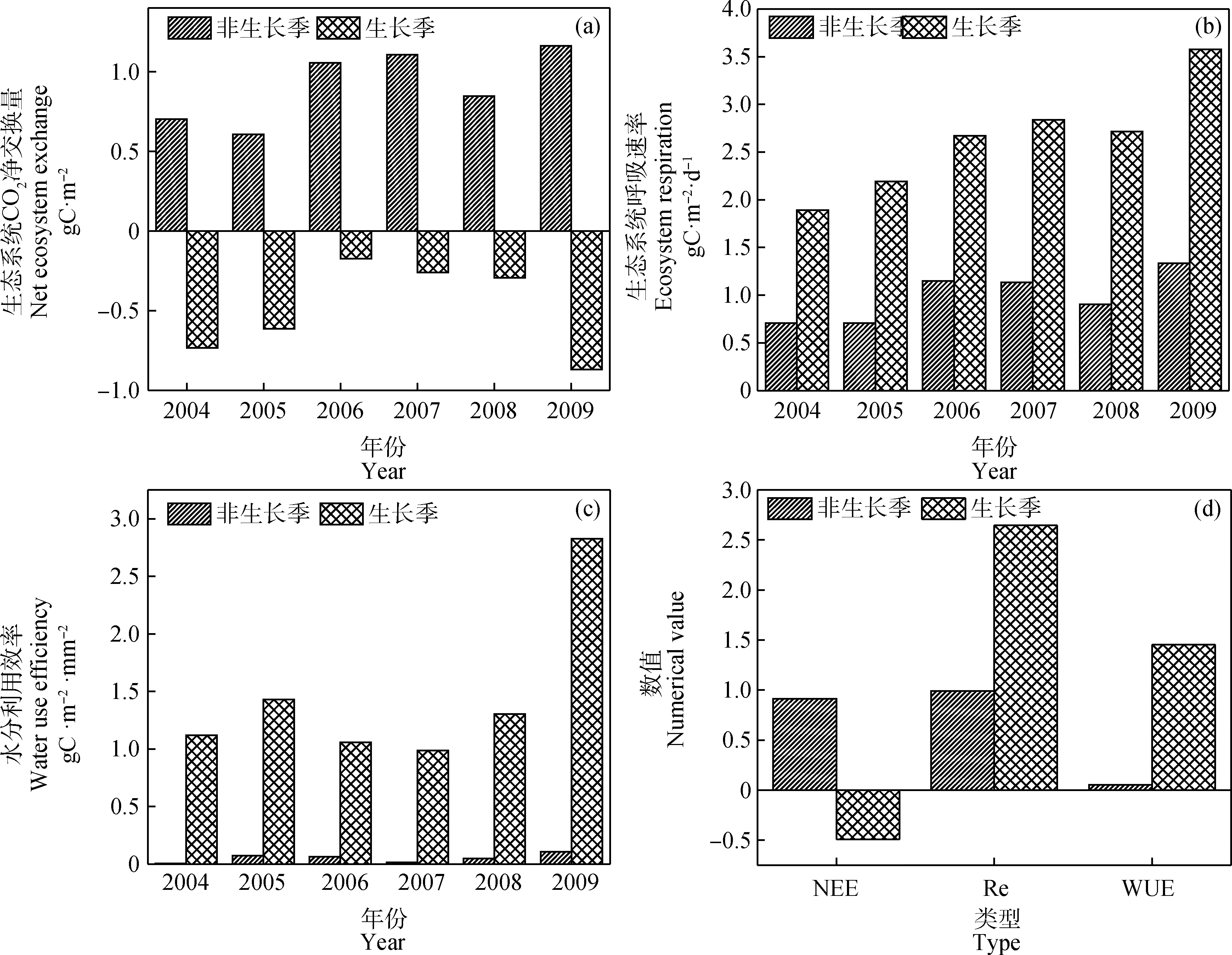
图2 生态系统CO2碳净交换量、生态系统呼吸速率和水分利用效率季节变化Fig.2 Seasonal changes of ecosystem net exchange of CO2,Ecosystem respiration and water use efficiency
2.3 高寒湿地生态系统水分利用效率对气象因子的响应
结构方程模型结果表明,在整个研究期间,降水量、风速、净辐射、空气温度和相对湿度是影响高寒湿地NEE的主要影响因子,且均达到极显著性水平(P<0.01),相对湿度对生态系统NEE的显著性影响最大,直接影响和间接影响效应系数值分别为-0.21和-0.09,净辐射影响次之,直接和间接影响的效应值分别为-0.16和0.19;相对湿度和空气温度是影响高寒湿地生态系统呼吸速率的极显著正效应影响因子(P<0.01)。直接和间接效应值分别为0.20,0.72和0.22,0.04;降水量和空气温度是显著影响高寒湿地生态系统水分利用效率变化的主要影响因子(P<0.05)。两者直接和间接效应值分别为0.05,0.08和0.02,0.01。

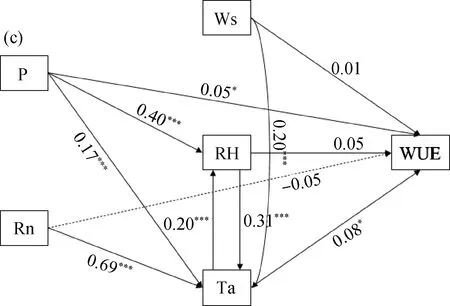
图3 环境因子对生态系统CO2净交换量、生态系统呼吸速率和水分利用效率影响Fig.3 Structural equation diagram of the influence of environmental factors on ecosystem net exchange of CO2,ecosystem respiration rate and water use efficiency注:实线表示正相关,虚线表示负相关;“***”和“*”表示在P<0.01和P<0.05达到显著性水平,P表示降水量,Ws表示风速,Rn表示净辐射,Ta表示空气温度,RH表示相对湿度Note:The solid line represents the positive correlation,the dashed line represents the negative correlation;“***” and “**” indicate significant levels at P<0.01 and P<0.05,P indicates precipitation,Ws indicates wind speed,and Rn indicates net radiation,Ta means air temperature,RH means relative humidity
3 讨论
本研究中,2004—2009连续6年间生态系统呼吸速率、生态系统CO2净交换量和水分利用效率均呈极显著上升趋势。季节尺度上,6年间非生长季生态系统碳排放总量大于生长季碳吸收总量。这与对寒区滨河湿地对碳通量的季节变化研究结果一致[30]。究其原因可能是因为在植物生长季昼夜温差大,有助于植物光合和光合产物的积累[31]。对于生态系统呼吸速率和水分利用效率均呈现出生长季值大于非生长季值。可能是因为在植被生长季,温湿度条件适宜时,其光合作用较强,植被生长良好,故植物生态系统呼吸速率和水分利用效率在植被生长季较高[32]。
植被自身和微生物的活动也会影响生态系统CO2净交换量[16]。本研究中,净辐射和相对湿度、风速对生态系统CO2净交换量存在极显著负效应影响。净辐射通过影响植物光合作用来影响高寒湿地夏季生态系统CO2净交换量[16]。青海隆宝滩湿地生态系统CO2净交换量与土壤湿度之间存在显著负相关关系,湿地水位降低有利于氧气的进入,加速了土壤有机物质分解,进而导致CO2排放增加[4]。
生态系统呼吸速率是草地生态系统进行碳交换的主要环节[11]。本研究中,生态系统呼吸速率主要受到相对湿度和气温的影响,且均呈正相关影响。气温的变化会引起植物物候和植被生产力的变化,进而伴随着生态系统呼吸速率的变化[33]。神农架大九湖泥潭湿地土壤温度通过影响植物和微生物呼吸酶活性、对气体和底物扩散速率等影响生长季夜间生态呼吸速率[34]。
降水量和温度是调控水分利用效率变化的主要因子,且与两者之间均呈正相关。大九湖泥潭湿地生态系统夏季水分利用效率变化与土壤温度之间呈现正相关关系,其余季度呈负相关关系,究其原因是因为土壤温度直接影响地表水分蒸发和通过影响酶活性和影响气体的扩散速度来影响水分利用效率变化[15]。降水量的变化对高寒湿地生态系统水分利用效率的影响存在阈值效应,在未达到阈值时,两者之间呈正相关关系,超过阈值后,水分利用效率变化基本不变,这是因为生态系统生产力不仅受到降水量的控制[35],还受到温度的控制,进而影响水分利用效率值[26]。
4 结论
年际尺度上,生态系统CO2净交换量、生态系统呼吸速率和水分利用效率均呈极显著上升趋势;季节尺度上,连续6年间,非生长季生态系统碳排放总量大于生长季碳吸收总量。针对于生态系统呼吸速率和水分利用效率均呈现出生长季值大于非生长季值。相对湿度和净辐射、相对湿度和空气温度、降水量和空气温度分别是影响青海海北高寒湿地生态系统CO2通量、生态系统呼吸速率和水分利用效率的主要气象因子。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
北京大学学报(自然科学版)(2022年3期)2022-06-17
农业灾害研究(2022年1期)2022-05-07
环境(2021年5期)2021-06-20
大气科学学报(2018年4期)2018-09-10
物联网技术(2016年11期)2017-01-12
中学化学(2016年10期)2017-01-07
中学化学(2015年8期)2015-12-29
