
高寒沼泽湿地退化过程中土壤腐殖质变化特征的研究
2022-06-02 01:36林春英李希来孙海松孙华方马程彪李成一
草地学报 2022年5期
林春英, 李希来, 张 静, 孙海松, 孙华方, 马程彪, 李成一
(1. 青海省气象灾害防御技术中心,青海 西宁 81000; 2. 青海大学农牧学院,青海 西宁 810016;3. 青海省地理国情监测院,青海 西宁 81000)
湿地生态系统既可是碳源,又可是碳汇,到底是哪种取决于不同的植被类型和环境变量[1]。自20世纪90年代以来,在自然因素和人为因素的双重驱动下,全球湿地出现不同程度的退化,湿地面积减少了一半[2]。世界范围内湿地的退化和丧失增加了全球温室气体总量,湿地的温室气体排放已占全球总排放的十分之一[3]。高寒沼泽湿地指海拔高、温度低的湿地[4],因其气候寒冷,土壤有机质分解缓慢,长期积累数量巨大的有机碳。近年来,随着气候变暖和人为干扰,高寒沼泽湿地的退化速度加快,逐渐向高寒草甸演替[5]。冻融丘作为高寒沼泽湿地的一种特征,是以藏嵩草(Kobresiatibetica)为主高出积水区的草丘。冻融丘之间的低洼地称为丘间[6],优势种为苔草(Carexmoorcroftii)。研究报道,随着退化程度的加剧,冻融丘明显减小,直到重度退化完全消失,且有机碳减少[6],碳功能的下降。土壤腐殖质是土壤有机碳的主要组成部分[7],是土壤有机碳库中较稳定的组分,在固碳和调控养分供应等方面发挥着重要的作用[8-10]。土壤腐殖质主要由胡敏酸(Humic Acid,HA)、富里酸(Fulvic Acid,FA)、胡敏素(Humic)等组成,其形成与转化对于维持有机碳库的稳定和土壤碳平衡意义重大[11-12]。目前,土壤腐殖质的研究领域多集中在农田、草原、森林及低海拔湿地土壤腐殖质的空间分布特征、剖面分布及其组成特征[13-15],针对高寒湿地土壤腐殖质的研究少见报道。高寒沼泽湿地退化过程中植被凋落物分解和根系分泌等变化,对土壤腐殖质的含量产生影响[16-18],在高寒沼泽湿地不同退化阶段,土壤的腐殖化程度和过程也不尽相同。此外,土壤酶来源于凋落物、根系分泌物及微生物活动[19],是土壤中碳氮的转化主要依赖物[20],土壤腐殖质各组分的形成与土壤酶密切相关[8,21],高寒湿地的退化导致的土壤理化性质和酶活性等发生变化,必然会影响到土壤腐殖质组分的形成和转化。因此,本文以不同退化程度高寒沼泽湿地土壤为研究对象,研究了土壤腐殖质组分含量的分布特征,并探讨土壤腐殖质分布与土壤理化性质和酶活性的相关性,以期为高寒沼泽湿地土壤碳库的碳汇研究提供科学依据。
1 材料与方法
1.1 研究区概况和样地布置
2020年8月,选择黄河源果洛州玛沁县大武滩高寒沼泽湿地与外围退化区作为土壤腐殖质变化研究调查样地。玛沁县地处果洛藏族自治州东北部,年平均气温约为-3.5℃,年降水量约为500 mm。大武滩处于果洛州玛沁县东南角,面积大约十平方公里,其高寒沼泽湿地资源丰富,且出现不同程度的退化。研究区未退化(Undegradation,UD)的高寒沼泽湿地(图1a)以冻融丘(图2)藏嵩草为优势种,其盖度可达95%左右,丘间(图2)的苔草等植物是主要伴生的物种;轻度退化(Light Degradation,LD)高寒沼泽湿地(图1b)冻融丘藏嵩草和丘间苔草的盖度各为50%;重度退化(Heavy Degradation,HD)高寒沼泽湿地(图1c)无冻融丘,是以矮嵩草为主的高寒草甸。本研究中,不同退化程度的湿地样方布置,结合高寒沼泽湿地中藏嵩草优势度、植被盖度多少和冻融丘大小、数量等指标综合判断[6],将试验样地划分为未退化、轻度退化和重度退化共3个退化程度。样方的调查采用线样法,从湿地中心拉3条样线,样线长为150 m,每隔50 m设置取样样方(图3),每个退化阶段各设置1个1 m×1 m的样方(即不同退化程度各设置3次重复),进行植被调查和土壤采样。

图1 高寒沼泽湿地不同退化程度样地Fig.1 Sample plots of alpine swamp wetland with different degrees of degradation注:(a)未退化;(b)轻度退化;(c)重度退化Note:(a)Non Degradation;(b)Light Degradation;(c)Heavy Degradation

图2 高寒沼泽湿地冻融丘和丘间示意图Fig.2 Frozen-thaw patch and between the grass patchs of alpine marshland
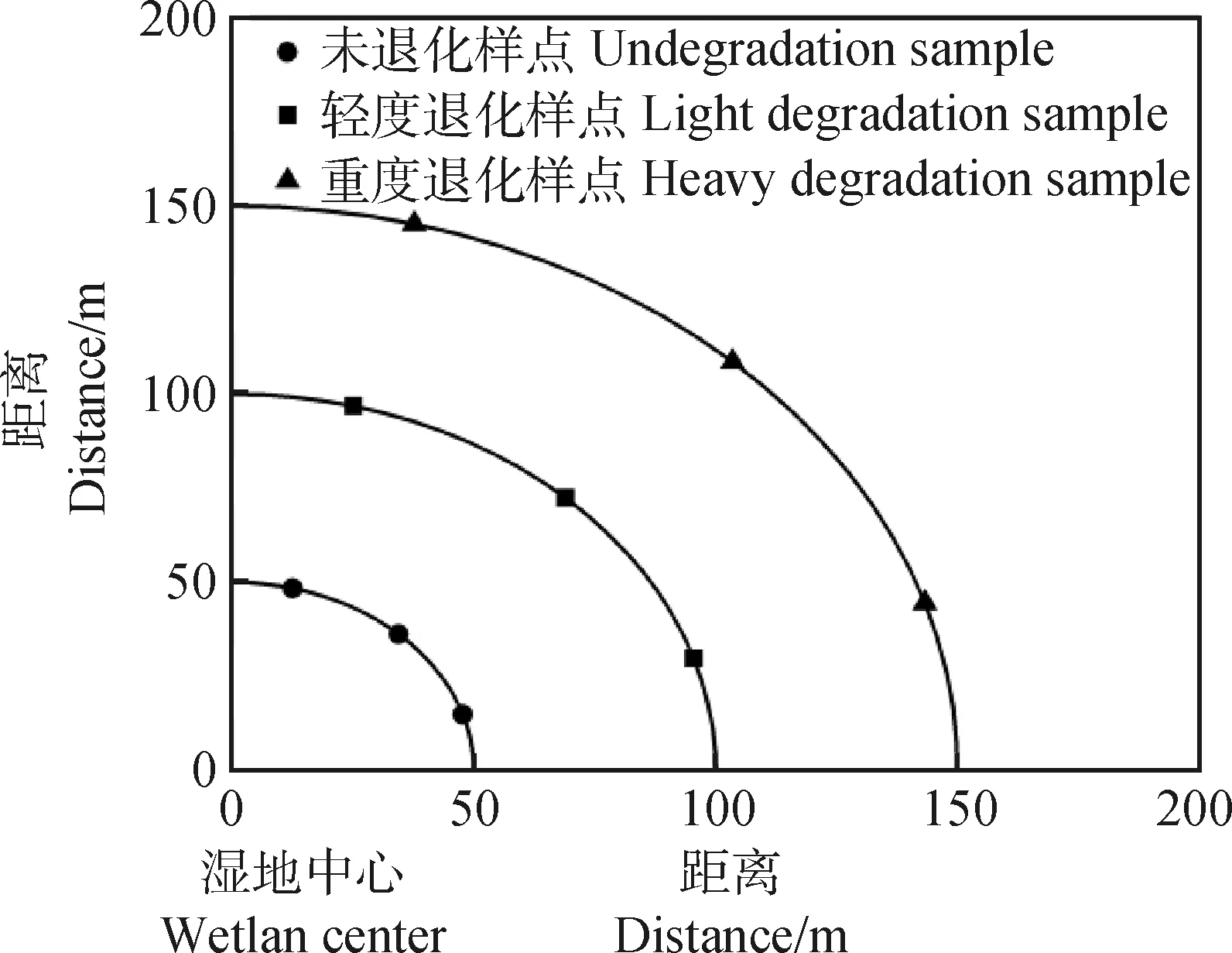
图3 高寒沼泽湿地不同退化程度样点选取图Fig.3 Sample pionts of alpine marshland with different degrees of degradation
1.2 土样样品采集
未退化和轻度退化阶段内的样方里有冻融丘和丘间,故分冻融丘和丘间采集土壤样品,因重度退化为高寒草甸,无冻融丘,故均一采样。自上而下用移除法分别采集0~10,10~20,20~30 cm三个深度的土样。将同一样地同一退化程度同一层的冻融丘和丘间土样分别装入自封袋,重复3次,共45个土样。每份土壤样品分为2份,一份自然风干后用于测定土壤腐殖质和全N、全P、全K,另一份鲜样(在4℃下储存)用于土壤微生物碳、纤维二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR)测定。
1.3 土壤指标测定
土壤pH采用pH计法测定;土壤容重采用环刀法测定;土壤含水量用便携式土壤水分测定仪TDR350测定;土壤腐殖质采用窦森法测定;土壤微生物碳采用氯仿熏蒸法测定;全N、全P、全K含量采用硫酸-过氧化氢混合消煮后分别以凯氏定氮仪和紫外分光光度计法测定;纤维二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)采用微孔板荧光法测定;脲酶(UR)采用比色法测定。
1.4 数据整理与统计分析
使用Microsoft Excel2010 对腐殖质数据进行处理,用SPSS19.0统计软件进行方差分析(one-way analysis)和相关性分析,利用Origin2018绘制不同退化程度高寒湿地土壤腐殖质变化图,使用R语言,制作土壤理化性质和腐殖质进行冗余分析(Redun dancy analysis,RDA)图。
2 结果与分析
2.1 不同退化程度高寒沼泽湿地土壤腐殖质变化特征
土壤腐殖质碳是胡敏酸和富里酸的总和,是土壤有机碳的主体,对提高土壤肥力有重要作用。冻融丘和丘间土壤腐殖质碳随着退化加剧呈减少趋势(图4),且冻融丘未退化与轻度退化、重度退化之间差异显著(P<0.05)。相对于未退化,轻度退化和重度退化0~10 cm冻融丘和丘间分别下降了28.73%,28.73%和14.52%,15.90%;10~20 cm分别下降了23.01%,21.87%和5.88%,0.01%;20~30 cm分别下降了31.18%,31.17%和4.30%,31.18%。
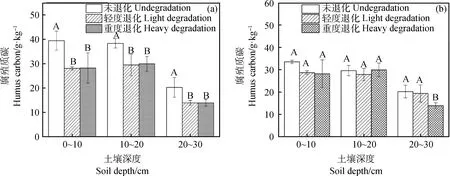
图4 不同退化程度高寒沼泽湿地不同土层腐殖质碳变化Fig.4 The change of humus carbon content of different soil layers in the alpine marshland注:图a为冻融丘,图b为丘间;不同大写字母表示相同土层不同退化程度间显著差异 (P<0. 05),下同Note:Fig. a,Frozen-thawing mound;Fig. b,Between the grass patches. The capital letters indicate significant difference among different degrees of degration in same soil depth at the 0.05 level,the same as below
由图5可知,冻融丘的胡敏酸含量较丘间高。随着退化加剧,冻融丘和丘间土壤胡敏酸呈减少趋势,且冻融丘0~10 cm、10~20 cm和20~30 cm未退化与轻度退化、重度退化之间差异显著(P<0.05)。相对于未退化,轻度退化和重度退化0~10 cm冻融丘和丘间分别下降了34.37%,34.29%和9.28%,17.72%;10~20 cm分别下降了21.12%,23.73%和7.46%,0.82%;20~30 cm分别下降了32.51%,35.24%和0.85%,30.66%。
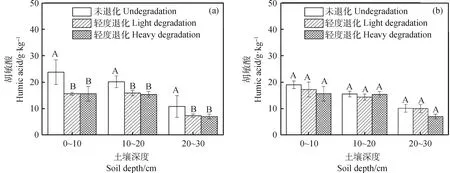
图5 不同退化程度高寒沼泽湿地不同土层土壤胡敏酸变化Fig.5 The change of humic acid content of different soil layers in the alpine marshland
随着退化加剧,冻融丘和丘间土壤富里酸呈下降趋势,但变化不明显(图6)。由表1可知,胡敏酸/富啡酸比值在冻融丘随退化程度的加剧呈减少趋势,且0~10 cm未退化与轻度退化、重度退化之间差异显著(P<0.05)。上述结果说明,未退化样地表层(0~10 cm)冻融丘土壤HA/FA比值均高于轻度退化和重度退化样地,土壤腐殖化程度较高,质量也较好,有利于HA的积累。随着退化程度的加剧,冻融丘HA/FA比值有所下降,土壤腐殖化程度较低。
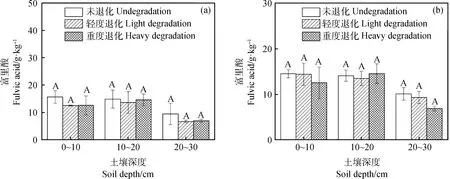
图6 不同退化程度高寒沼泽湿地不同土层土壤富里酸变化Fig.6 The change of fulvic acid carbon content of different soil layer in the alpine marshland
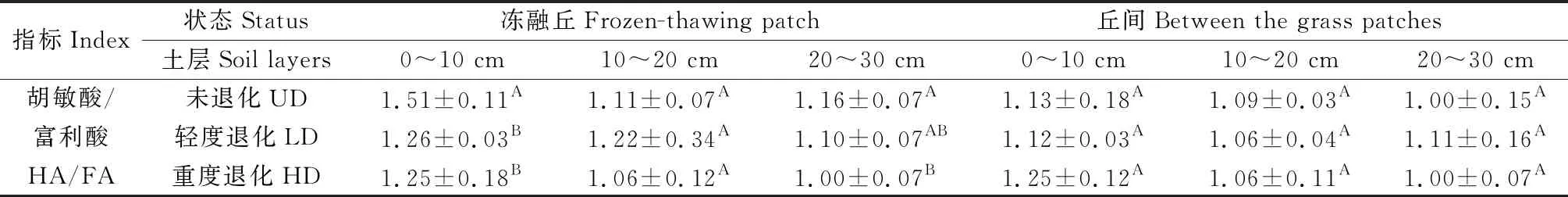
表1 不同退化程度下土壤胡敏酸与富里酸的比例Table 1 The ratio of soil humic acid to fulvic acid under different degradation degrees
如图7所示,冻融丘和丘间土壤胡敏素随着退化加剧呈减少趋势,且冻融丘和丘间各层未退化与轻度退化、重度退化之间差异显著(P<0.05)。相对于未退化,轻度退化和重度退化0~10 cm冻融丘和丘间土壤胡敏素分别下降了43.55%,46.44%和35.56%,38.16%;10~20 cm分别下降了33.66%,33.38%和51.92%,33.07%;20~30 cm分别下降了49.98%,45.54%和25.66%,53.03%。
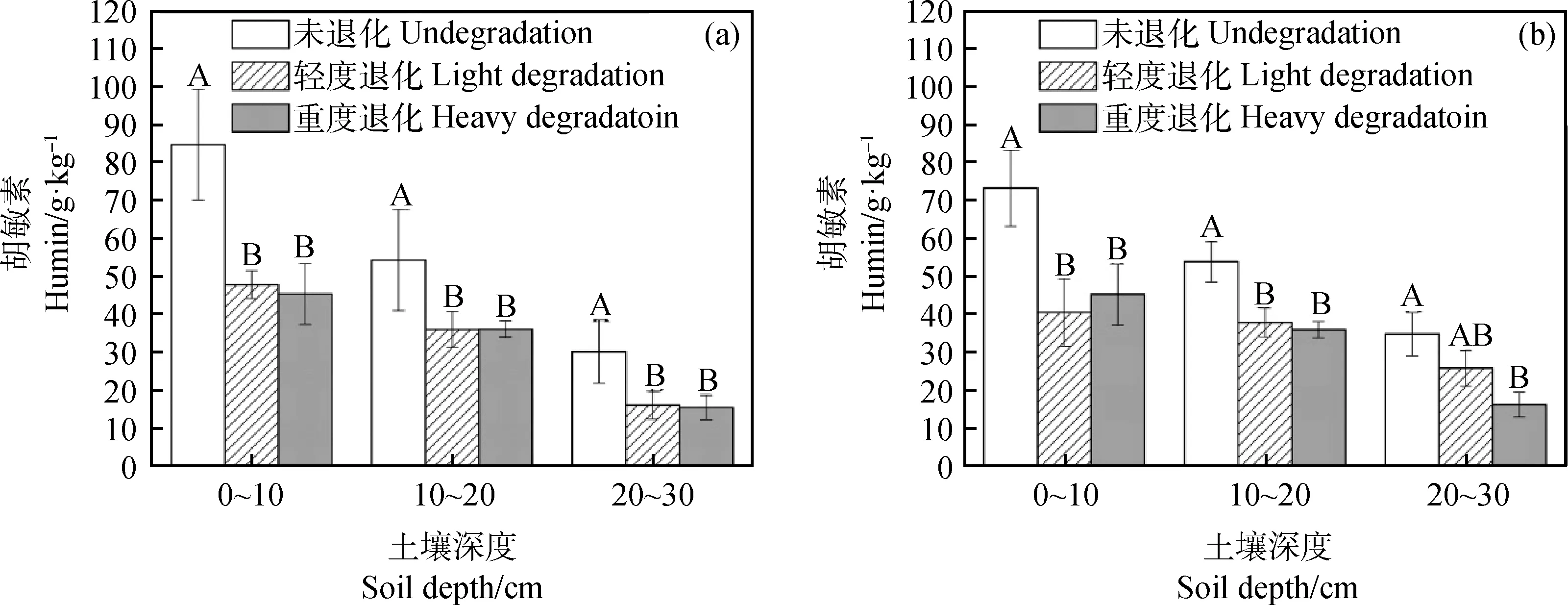
图7 不同退化程度高寒沼泽湿地不同土层土壤胡敏素变化Fig.7 The change of humin content of different soil layers in the alpine marshland
2.2 土壤腐殖质与土壤理化性质相关分析
土壤酸碱性、容重、含水量和土壤养分供应是限制微生物活动的重要因素,它们可通过微生物间接影响土壤腐殖质的形成和分解。本研究不同退化程度高寒沼泽湿地土壤理化性质见表2,土壤有机碳、土壤含水量、总氮、土壤腐殖质在未退化(UD)样地水平较高,随着高寒湿地退化程度的加剧逐渐减小;pH、TK则在重度退化(HD)样地较高,随着退化程度的加剧逐渐增大。从土壤理化性质与土壤有机碳及其组分排序看出(图8),在0~30 cm冻融丘和丘间土壤含水量、总氮与土壤腐殖质正相关,pH、TK与土壤腐殖质负相关。分析高寒沼泽湿地土壤理化性质与土壤腐殖质相关性发现(表略),冻融丘和丘间与腐殖质碳、胡敏酸、富里酸和胡敏素含量均呈显著正相关作用的指标为全N和土壤含水量;冻融丘和丘间pH和全K与腐殖质组分的含量均表现负相关关系,且pH表现显著的负相关(P<0.01);冻融丘所有腐殖质组分均与土壤容重呈显著负相关(P<0.01)。由此可见,土壤腐殖质各组分的形成与土壤氮素含量存在显著相互促进作用,能显著的提高土壤的保肥能力。
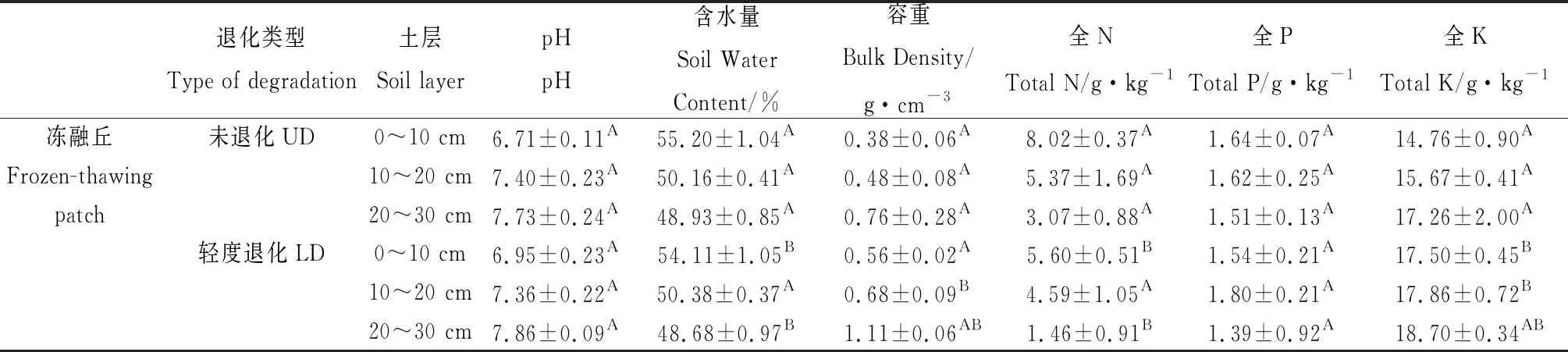
表2 不同退化程度高寒沼泽湿地土壤的理化性质Table 2 Physical and chemical properties of the soils at different soil layers in the alpine marsland
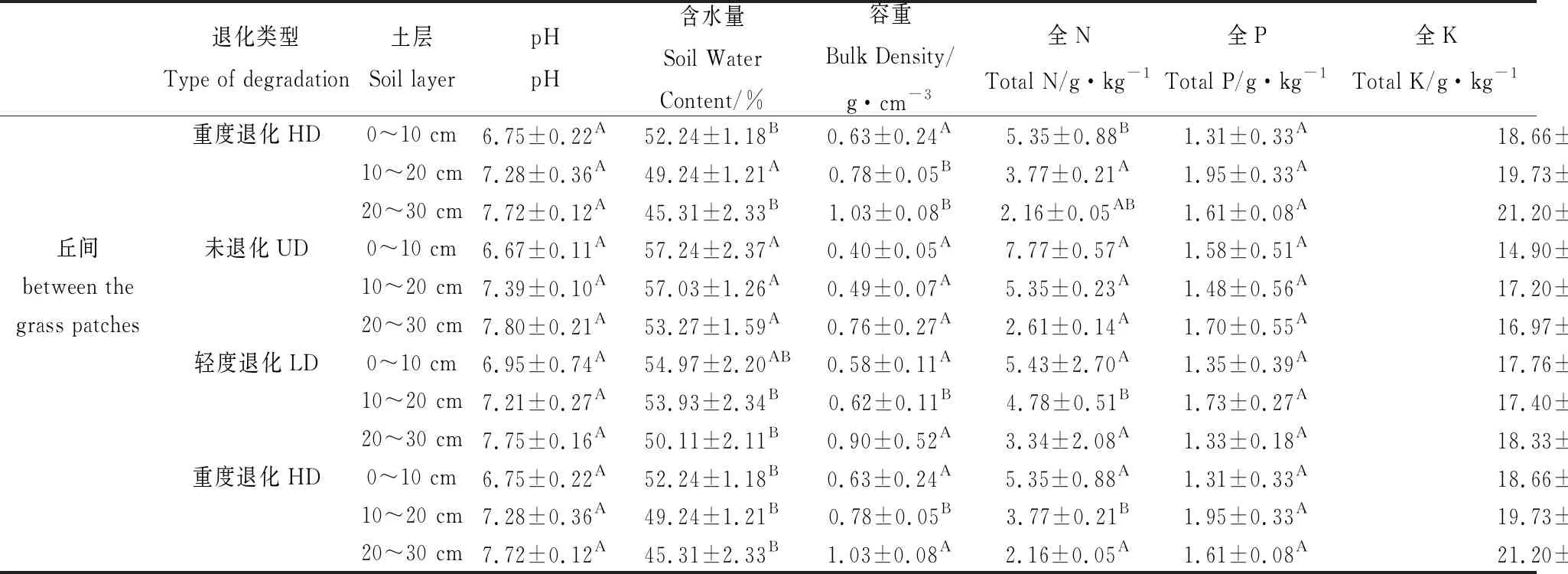
续表2
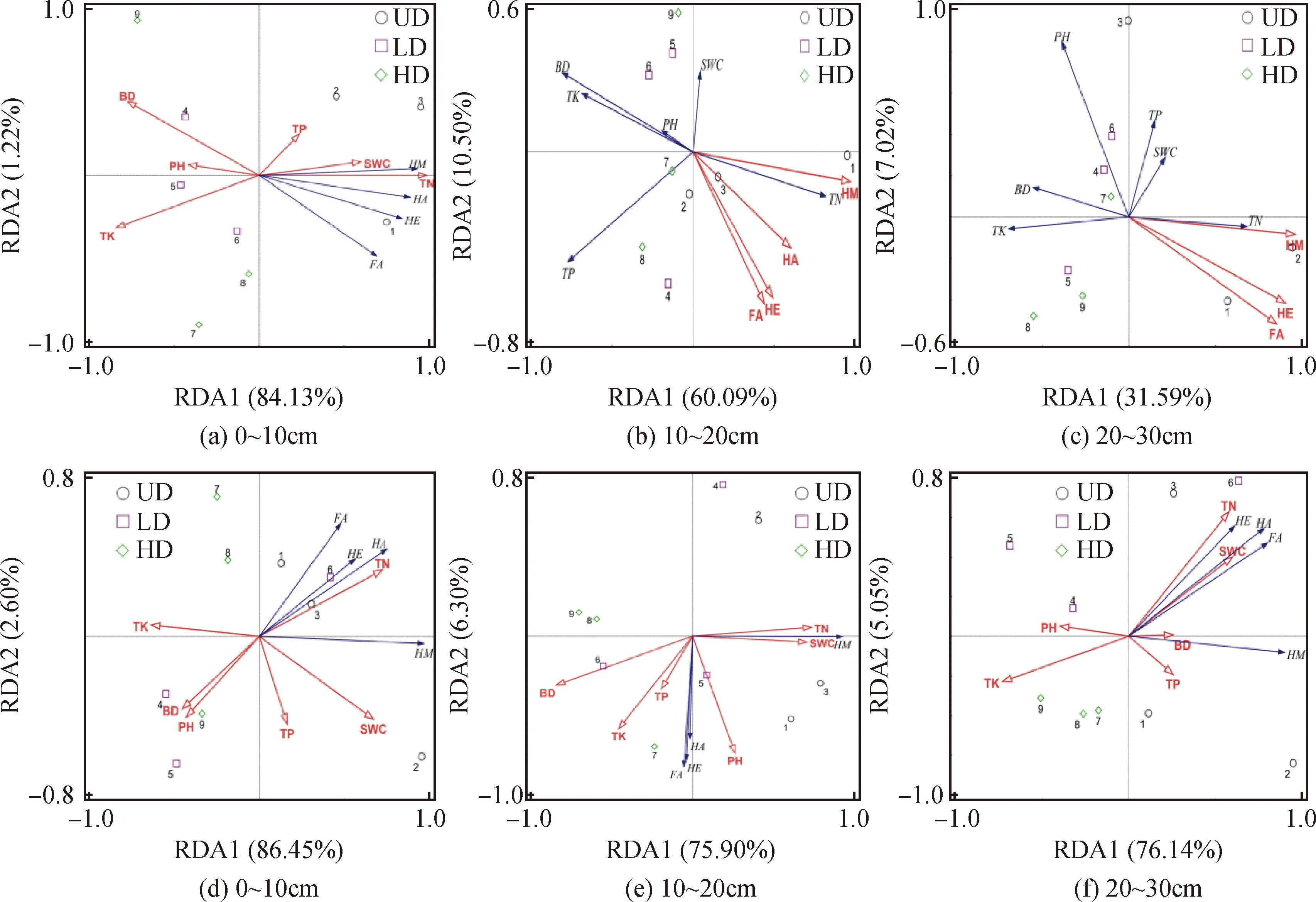
图8 土壤理化性质和土壤腐殖质之间的冗余分析二维排序图Fig.8 The ordination diagram of RDA analysis of vegetation and soil parameters注:BD,容重;SWC,土壤含水量;pH,酸碱度;TN,全氮;TK,全钾;TP,全磷;(a)(b)(c)为冻融丘,(d)(e)(f)为丘间Note:BD,Bulk density;SWC,Soil moisture content;pH,Potential of hydrogen TN,Total nitrogen;TK,Total potassium;TP,Total phosphorus. Fig. a,b,c,frozen-thawing mound;Fig. d,e,f,between the grass patches
2.3 土壤腐殖质与微生物碳、土壤酶活性的关系分析
土壤腐殖质是土壤有机质在微生物和酶促反应下逐步降解和转化的结果,同时土壤腐质化过程又会对微生物和土壤酶活性产生反馈作用[16]。研究区不同退化程度高寒沼泽湿地土壤生物学性质见表3。分析土壤腐殖质含量及构成与微生物碳、土壤酶活性之间的相互关系发现(表4),冻融丘中与腐殖质碳含量呈显著正相关的指标为β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)和β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG);与胡敏素含量呈显著正相关作用的指标为纤维二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)磷酸酶和脲酶(UR);与胡敏酸含量呈显著正相关的指标为纤维二糖水解酶(CBH)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR);而富里酸含量与微生物碳和土壤酶活性之间差异不显著。丘间中与腐殖质碳、胡敏素、胡敏酸和富里酸含量与纤维二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR)呈显著正相关,胡敏素与土壤微生物碳正相关(P<0.01),而腐殖质碳、胡敏酸和富里酸含量与微生物碳之间差异不显著。由此可见,丘间纤维二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR)与腐殖质的形成具有显著的促进作用,微生物碳对胡敏素形成有显著作用;而冻融丘中β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)和β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)对腐殖质碳形成具有显著的促进作用,纤维二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR)对胡敏素形成具有显著的促进作用,纤维二糖水解酶(CBH)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR)对胡敏酸形成具有显著的促进作用。
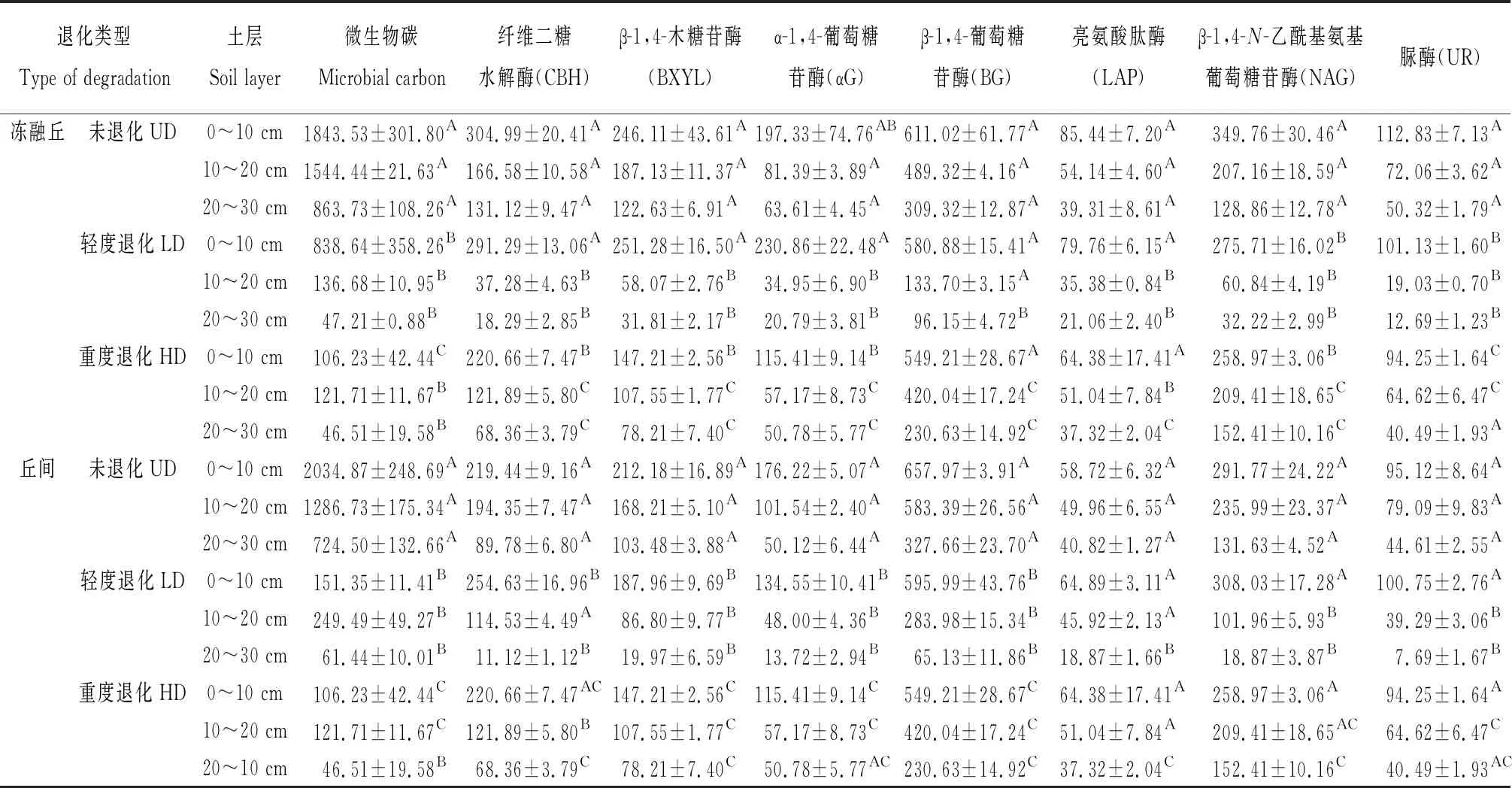
表3 不同退化程度高寒沼泽湿地土壤生物学性质Table 3 Biological properties of soil at different soil layers in the alpine marshland
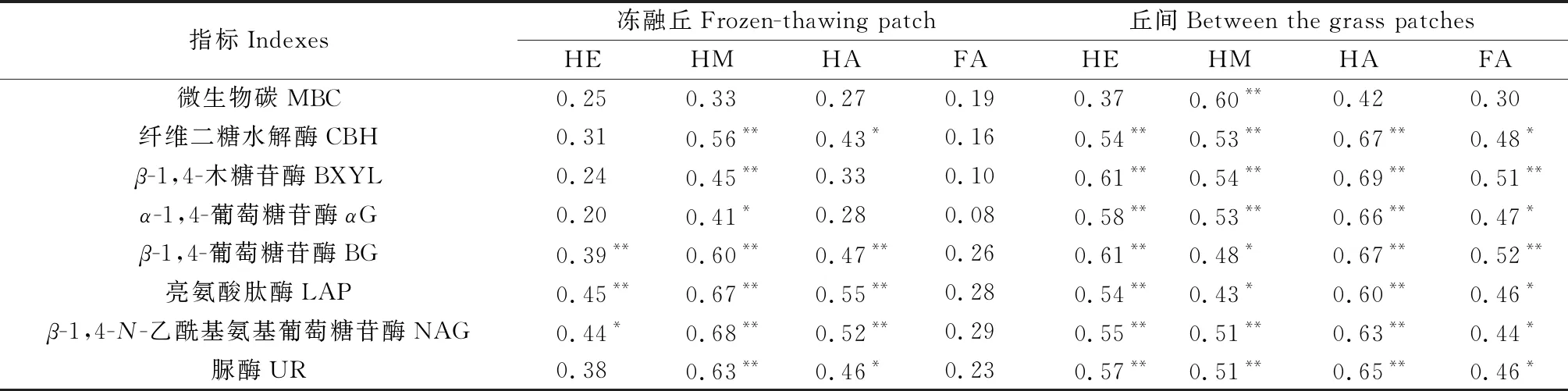
表4 不同退化程度高寒沼泽湿地土壤腐殖质与土壤生物学性质的相关性Table 4 Correlation coefficients between soil humus and soil biological properties in different degraded alpine marshlands
3 讨论
3.1 退化对高寒沼泽湿地土壤腐殖质的影响
本研究中,3种退化程度的高寒沼泽湿地土壤的腐殖质性质存在显著不同现象,土壤腐殖质含量高的未退化湿地土壤腐殖质稳定性较高,腐殖质含量较低的重度退化湿地土壤腐殖质稳定性较差(图4,5,6,7和表1),且冻融丘的腐殖质含量和稳定性较丘间高。各土层的土壤腐殖质组成大小均为:胡敏素>腐殖质碳>胡敏酸>富里酸。土壤腐殖质碳由土壤胡敏酸和富里酸组成,含量反映了土壤腐殖质的特性。高寒沼泽湿地随着退化程度的加剧,胡敏酸和富里酸含量均呈下降趋势,且冻融丘胡敏酸含量未退化与轻度退化、重度退化差异显著(P<0.05)。HA/FA反映土壤腐殖物质聚合程度,其值大表明土壤腐殖物质的聚合度高,土壤腐殖化的程度越高,质量也越好[22]。不同退化高寒沼泽湿地土壤腐殖质的组成中,HA/FA均大于1,这表明高寒沼泽湿地土壤为胡敏酸型土壤,土壤腐殖物质的聚合度较高。冻融丘土壤胡敏酸与富里酸之比随退化程度的加剧大致呈下降趋势,且0~10 cm未退化和轻度退化、重度退化差异显著(P<0.05),说明随着退化程度的加剧冻融丘土壤腐殖物质的聚合度下降,土壤腐殖化的程度下降。
研究区表层(0~10 cm)土壤腐殖质的含量变化幅度较大(图4,5,6,7),但随着土层的加深,含量差异逐渐减少,这主要与植物根系分布和凋落物数量及质量有关。研究区的未退化区冻融丘植物以莎草科植物藏嵩草为主,具有较高的地上生物量和盖度,每年回归土壤的丰富有机残落物增加了有机质的含量。随退化程度的加剧,植物种内竞争加剧,群落优势种更替,物种增多,植被盖度、高度和地上生物量持续降低,所生成的有机质较少。同时,冻融丘以莎草科植物藏嵩草为主,表层大量的凋落物和植物根系,为微生物生长提供了来源[6]。此外,冻融交替对高寒沼泽湿地土壤温度产生影响,增加土壤呼吸量,且杀死部分土壤微生物,死亡的微生物分解后释放出氨基酸和糖类等物质[23],提高了高寒沼泽湿地土壤中有机碳的含量,进而提高了高寒沼泽湿地土壤的腐殖质的含量。
3.2 土壤理化性质对腐殖质的影响
本研究中,冻融丘和丘间土壤全氮与腐殖酸碳、胡敏酸、富里酸和胡敏素表现为正相关关系,这与张晓曦[16]和党亚爱等[24]研究结果基本吻合。这由于土壤全氮提高了微生物的数量与活性,同时也为腐殖质合成提供了氮源。土壤水分条件作为决定高寒沼泽湿地植物群落分布和演化的主导因素[6],当高寒沼泽湿地退化后,水分条件发生变化,将影响高寒沼泽湿地植被特征和有机碳的含量,从而影响土壤腐殖质的含量。冻融丘和丘间土壤含水量与腐殖酸碳、胡敏酸碳、富里酸和胡敏素含量正相关关系,说明土壤水分是限制有机质的分解和土壤腐殖质的形成的主要环境因子。高寒沼泽湿地退化使土壤水分减少,土壤腐殖质的下降,不利于有机碳的积累,使碳功能下降。冻融丘和丘间土壤容重和pH与腐殖酸碳、胡敏酸、富里酸和胡敏素含量表现为负相关关系,土壤容重通过影响土壤孔隙度、通气状况直接对土壤微生物产生影响,而pH通过影响土壤中微生物种类、数量及其活性,从而对土壤腐殖质产生影响。冻融丘和丘间土壤腐殖质含量与全K呈负相关,由此可以推断,全K含量增加会造成土壤腐殖质含量降低。
微生物活动影响土壤腐殖质形成及转化,土壤的有机质经土壤微生物分解后产生土壤腐殖质,该过程有多种土壤酶参与[21]。由于高寒沼泽湿地退化过程中地上植被发生变化,致使土壤真菌和细菌群落结构发生变化,且对碳源利用率增加[5]。而土壤酶活性受土壤细菌和真菌群落结构的影响,从而导致土壤腐殖质积累量的不同。本研究表明,冻融丘中4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)和β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)有利于对腐殖质碳的形成;二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)、脲酶(UR)有利于对胡敏素的形成;二糖水解酶(CBH)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR)有利于对胡敏酸的形成;丘间纤维二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)、β-1,4-葡萄糖苷酶(BG)、亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)、脲酶(UR)和微生物量的增加有利于土壤腐殖质的形成,这与于江[25]和张晓曦[16]等的部分结论相吻合。
高寒沼泽湿地土壤有机碳和总氮的变化紧密相关[6,26],且随着退化程度的加剧它们呈减少趋势。本研究中参与碳循化的β-1,4-葡萄糖苷酶(βG)、二糖水解酶(CBH)、β-1,4-木糖苷酶(BXYL)、α-1,4-葡萄糖苷酶(αG)和参与氮循化的亮氨酸肽酶(LAP)、β-1,4-N-乙酰基氨基葡萄糖苷酶(NAG)和脲酶(UR)随着退化程度的加剧呈减少趋势(表4)。由此可以推断,随着退化程度的加剧,地上凋落物、根系分泌物及微生物活动减少,导致土壤酶活性减弱[27],减缓了土壤中碳的周转,不利于土壤有机碳的积累。
已有研究表明,随着退化程度的加剧土壤有机碳、轻组分有机碳、重组分有机碳、可溶性有机碳和微生物碳呈下降趋势[6,28],且冻融丘较丘间差异显著。本研究结果显示,土壤腐殖质含量随着高寒沼泽湿地退化程度的加剧也呈减少趋势,这意味着高寒沼泽湿地退化使得冻融丘的逐步消失和土壤有机碳含量的降低,以CO2等温室气体的形式向大气释放碳,温室气体会导致气温上升。青藏高原高寒沼泽湿地是高原生态系统有机碳密度最高的地区,也是气候变化最敏感的地区之一。研究区从未退化到重度退化每平方米有机碳减少了294.94 g·kg-1,而土壤腐殖质作为土壤有机碳的主要组成部分,占土壤总有机碳 75%~85%,由此可推算出青藏高原250万km2共损失有机碳73 667 g·kg-1,损失土壤腐殖质大约55 250~62 616 g·kg-1,对该地区的碳库、碳汇和实现碳达峰碳中和目标不利。
4 结论
高寒沼泽湿地退化后,冻融丘和丘间土壤腐殖质呈下降趋势,冻融丘土壤腐殖质碳、胡敏素和胡敏酸未退化与轻度退化和重度退化差异显著(P<0.05),对退化较丘间敏感。随着退化程度的加剧冻融丘土壤腐殖物质的聚合度下降,土壤腐殖化的程度下降。土壤含水量和总氮与土壤腐殖质密切相关,它们是限制高寒沼泽湿地土壤腐殖质形成的主要环境因子。冻融丘中土壤酶对胡敏素形成具有显著的促进作用,丘间土壤酶活性对土壤腐殖质的形成具有显著的促进作用,微生物碳对土壤胡敏素的形成具有显著的促进作用。因此,在加快退化高寒沼泽湿地生态恢复时重点考虑冻融丘的保护和加强土壤水分和有机肥的补充。
猜你喜欢
中草药(2022年20期)2022-11-15
食品科学(2022年20期)2022-10-31
现代食品科技(2022年9期)2022-10-09
冰川冻土(2022年2期)2022-06-14
科技创新导报(2021年33期)2021-04-17
当代化工(2019年10期)2019-12-02
丝路视野(2018年15期)2018-05-14
广东教育·高中(2017年11期)2017-12-04
现代营销·经营版(2010年5期)2010-05-14
女士(2009年2期)2009-03-07
