
白刺叶片解剖结构性状异速生长与生态适应性研究
2022-06-02 02:01:58任尚福
草地学报 2022年5期
任尚福
(1. 喀什大学生命与地理科学学院,新疆 喀什 844000; 2. 新疆帕米尔高原生物资源与生态重点实验室,新疆 喀什 844000)
植物个体发育过程中,不同器官(如根、茎、叶、种子等)相互协调发展是植物生长的一种生活史策略,在其协调发展过程中,其生物学特征都直接或间接地对某种特定的环境产生相应的权衡策略[1]。植物在长期的适应进化过程中逐渐形成了与环境相适应的叶片结构[2]。植物性状反映了植物与环境相互作用,逐渐形成了内在生理和外在形态方面的适应对性,减小环境的不利影响[3]。叶片是植物进化过程中对环境变化较敏感且可塑性较大的器官,在不同选择压力下已经形成各种适应类型,其结构特征最能体现环境因子的影响或植物对环境的适应[4]。植物的叶片性状与植物利用资源的能力关系比较紧密,能够反应植物适应环境变化所形成的生存对策[5]。根据张海娜对荒漠区植物叶片厚度和叶肉(栅栏组织和海绵组织)等光合作用器官结构特征的研究,叶片解剖性状沿着有利于提高水分利用效率的方向发展[6]。相关学者研究表明植物叶片解剖结构受海拔[7-8]、降水量[9]、蒸发量[9]、日照[10]等环境因子的显著影响,主要表现为叶形的变化、叶片的大小、厚度及解剖结构的差异。
植物叶片功能性状间多呈异速生长关系,各叶片性状因子间异速生长关系存在明显差异[11]。异速生长指生物体某2个性状的相对生长速率不相同的现象[12],反映了生物体不同性状间的相关性,是植物生长发育过程中器官与组织呈现的一个重要规律。异速生长由物种遗传物质所决定[13],同时受环境因素的影响,是植物对环境做出的生态对策[13-15]。植物的异速生长是对异质生境适应的重要体现,可以揭示植物生物学特征与尺度无关的内在规律,而且已扩展到从基因组水平到生态系统水平的各个方面[16],其中植物叶内的生物量分配及生长关系是近年来的研究热点之一[17-18]。
当今植物叶片性状异速生长规律的研究对大尺度种间水平上关注比较多,但对种内水平叶片解剖性状之间的异速生长研究鲜有报道,John等对生活在湿润和干燥生境的14个不同被子植物物种的叶片进行解剖研究,发现各组织间的细胞壁厚度均呈等速生长关系;实验材料中的叶片厚度与栅栏组织厚度间呈等速生长关系,而与其他组织之间均呈异速生长关系[19]。白刺作为南疆干旱荒漠区的优势种植物,中国境内分布的白刺有6种,在改良盐碱地方面的效果显著。有关学者对西伯利亚白刺(NitrariasibiricaPall.)、唐古特白刺(N.tangutorumBobr.)、大果白刺(N.roborowskiiKom.)等进行了大量生理生化及种子休眠萌发的研究,但对不同居群的3种白刺叶片解剖性状叶片异速生长规律研究未见报道。因此,本研究选取来自南疆不同生境的大果白刺、西伯利亚白刺和唐古特白刺等3种优势种植物,研究3种不同白刺叶片解剖性状对环境变化的响应以及叶片解剖性状之间的异速生长关系,探讨其叶片解剖结构特征对不同生境的生态适应性。通过对生长在南疆荒漠区不同生境(主要包括海拔、平均日照、年降水量、年蒸发量)下的3种白刺科植物的叶片解剖性状进行分析,对叶片各解剖性状之间的异速关系采用异速方程Y=βXα描述,试图探讨白刺科植物解剖指标与环境因子的关系,拟解决以下2个科学问题:(1)3种白刺科植物叶片解剖性状在不同生境下的异速生长规律是怎样的?(2)其异速生长规律是否可以揭示白刺科植物在不同生境下的生态适应性?
1 材料与方法
1.1 研究材料与研究区概况
白刺属植物原为蒺藜科的一个古老小属,属于第三纪孑遗植物,是地中海—西亚至中亚分布类型[20]。白刺生长在西北干旱区的沙漠、戈壁和低山,是重要的防风固沙植物,在半干旱至干旱区草原、荒漠植被中具有相当重要的位置[21]。在《Flora of China》第11卷[22]中,对蒺藜科植物的系统关系进行了重新划分:将白刺属列入单属科白刺科(Nitariaceae),约6~10种,我国共有5~6种[23]。
研究区位于塔克拉玛干沙漠西南部,具有典型的大陆性暖温带荒漠气候,根据当地气象局提供的数据资料(2015—2017年),当地年降水量68~172 mm,年平均降水量78.6 mm,8月份降雨量最高为35.2 mm,1月和12月降水量最低为1 mm,但蒸发比较强烈,一般年蒸发量达到2 500 mm以上,日照时间较长,平均日照时间为2 600~3 000 h,昼夜温差比较大,年平均气温为14℃,7月最高气温为28℃,当地最低气温出现在1月。无霜期时间长约为180~240 d。南疆荒漠区除了白刺科植物以外主要伴生种有骆驼刺、柽柳、野西瓜、罗布麻、甘草、沙棘、沙拐枣等。
1.2 研究方法
1.2.1样品采集 根据3种不同白刺植物所处的经度、纬度和海拔等不同的生境各选取3个研究区,在每个研究区内选择3个3 m×3 m的样方,在每个样方内随机选取3株生长健壮、长势一致的植株(不同生境实验材料的采集其具体地理分布及生境信息见表1)。在每棵白刺的中部取大致发育程度相同的成熟叶片,并且用锋利的刀片在叶脉中间位置切取3~5 mm的小块,立刻放在装有FAA固定液(70%无水酒精90 mL+福尔马林5 mL+冰醋酸5 mL)的瓶中密封并且编号,并且用医用注射器抽取瓶中的空气,使瓶中形成部分真空,固定材料可以下沉到固定液瓶底部[24]。
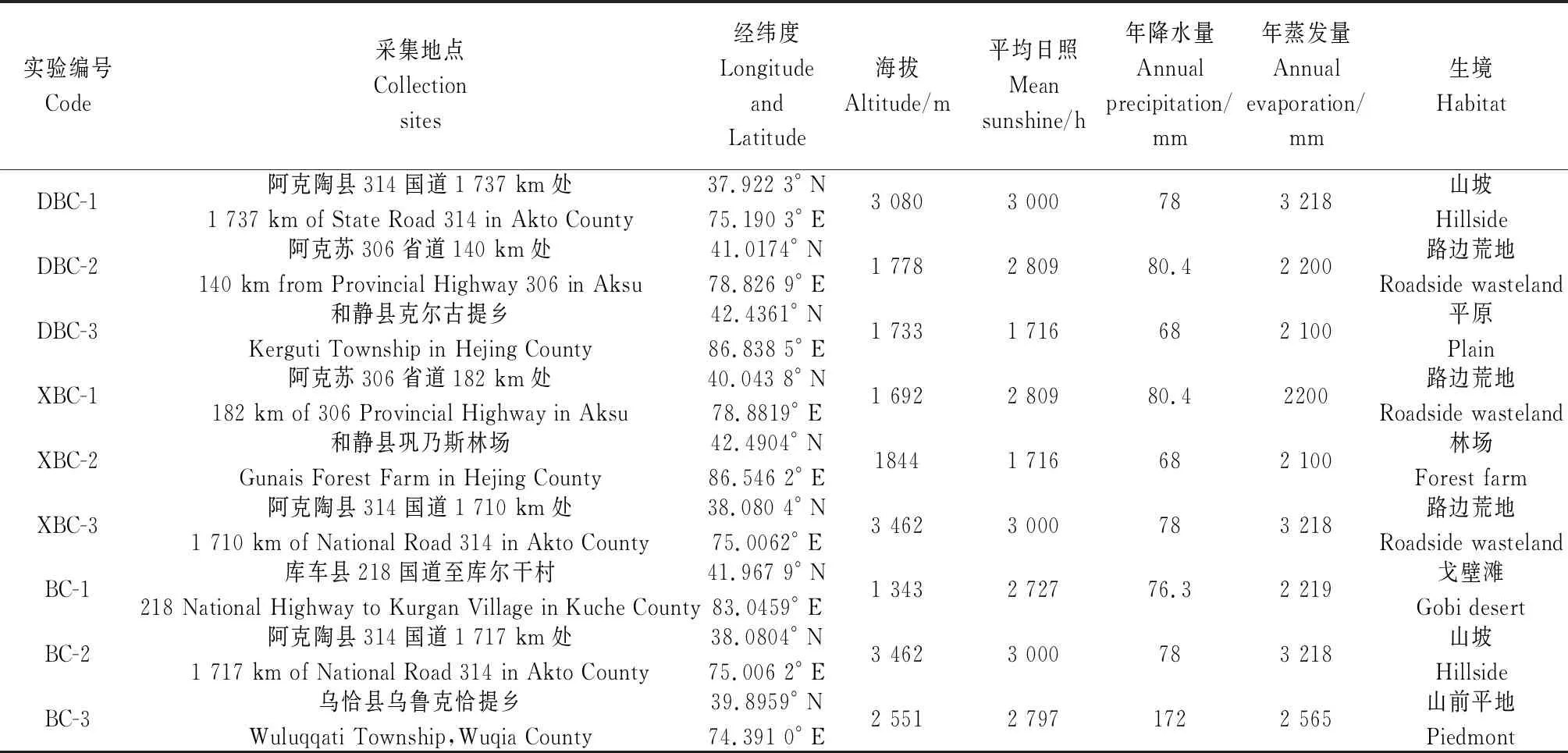
表1 叶片采集地点及气候特征Table 1 Leaf collection location and climate characteristics
1.2.2研究方法 每种白刺均从不同居群中各选取3个样本,共采取9个样本,在FAA固定溶液中取出小块叶片,采用改良的常规石蜡切片法制成叶片横切面永久切片[24]。
1.2.3数量性状测定方法 每株叶片样本选3张切片,每个样片切片选取5个观察视野(10个细胞以上),用DMB5-223I-5图像处理显微镜照相,并用显微测量软件Arcview3.3(ESRI,USA)对图像上的各种组织的厚度进行测量,测定指标包括叶片厚度、上角质层厚度、上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度及下角质层厚度等。
1.2.4数据分析 植物叶片解剖各性状之间的异速生长关系计算采用幂次方程
Y=βXα
式中:Y为植物某一性状;X为另一性状;α为异速生长指数;β为异速生长常数[25]。当α的95%置信区间包含1时,将Y和X之间的标度关系指定为等距关系,否则标度关系则为异速生长关系。α大于1表明,Y的线性增加比X更快;而α小于1表明Y的线性增加比X缓慢[5,7,19](表2)。
数据处理用R软件R语言INLA程序包,采用完全嵌入拉普拉斯近似方法(Integrated Nested Laplace Approximations Algorithm,INLA)进行近似贝叶斯推断[26]。检验各性状之间生长指数和异速生长系数的差异显著性。根据测量数据,计算各指标均值、标准差以及变异系数。


表2 异速生长指数值判断标准Table 2 Judging criteria for the value of allometric growth coefficient
2 结果与分析
2.1 南疆荒漠区不同生境的环境因子的异质性
在大尺度范围植被分布的主导因素是气候和地貌,在小尺度范围下对植被分布起关键作用的是微环境。从表1可知,不同生境下的各种环境因子(海拔高度、平均日照、年降水量、年蒸发量)的差异均达到极显著水平;计算不同生境的各环境因子的变异系数,并进行分析,其结果可以反映各生态因子的变异特点(表3),经计算各生态因子的变异系数均在0.200~0.374之间,按照变异系数的划分等级(CV=0.1~1.0属于中等变异),即在本实验中各生态因子均存在着中等程度的变异[27],表明同一生态因子在不同生境中是异质性的。

表3 各生态因子变异系数(CV)Table 3 Coefficient of variation of each ecological factor(CV)
2.2 不同生境唐古特白刺叶片性状的异速生长规律
变异系数表示实验各观测性状的变异程度,反映某物种群体与个体之间的差异。不同生境下大果白刺叶片解剖性状的变异系数在0.10~0.36之间(表4),表示不同生境唐古特白刺叶片解剖结构的变异性为中等变异,其中栅栏组织厚度变异系数最小,下表皮厚度变异系数最大,根据蔡永立等的研究,变异系数较小的被认为是相对稳定的系统演替性状[2,28]。
观察3个不同生境下唐古特白刺叶片解剖石蜡切片,记录叶片解剖性状指数。分析唐古特白刺叶片厚度与上角质层厚度等21组性状间的异速生长关系,结果表明:所有21组性状之间关系均呈显著相关(P<0.05)(表5),其中上角质层厚度与海绵组织厚度、上表皮厚度与下表皮厚度、栅栏组织厚度与下角质层厚度之间的生长指数(95%置信区间)包含1,说明这些性状可能会以相同的速率生长,根据异速生长系数值判断标准(表3)这些生长指数之间属于同速生长关系。除此之外,其余所有的2个解剖指标厚度之间的生长指数(P<0.05)与1差异显著并且都小于1,Y的线性增加比X缓慢,说明这些性状之间存在异速生长,并且呈现非等比例增长,增长速率前者小于后者,根据异速生长系数值判断标准(表3),这些生长指数之间属于负异速生长关系。决定系数(R2)位于0.710 5~0.909 6之间。
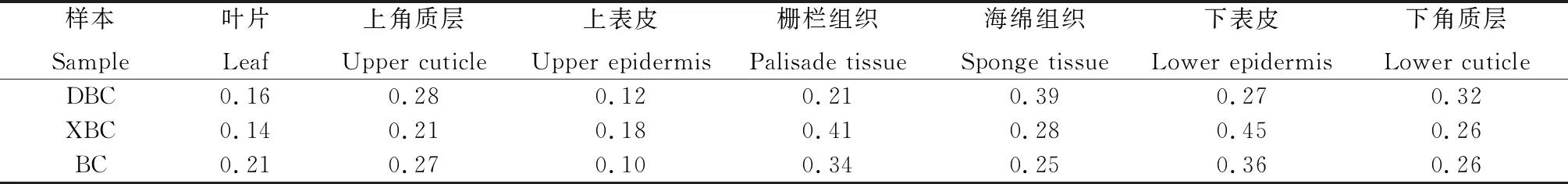
表4 3种白刺植物叶片解剖结构性状变异系数Table 4 Coefficients of variation of leaf anatomical structure traits in three Nitariaceae
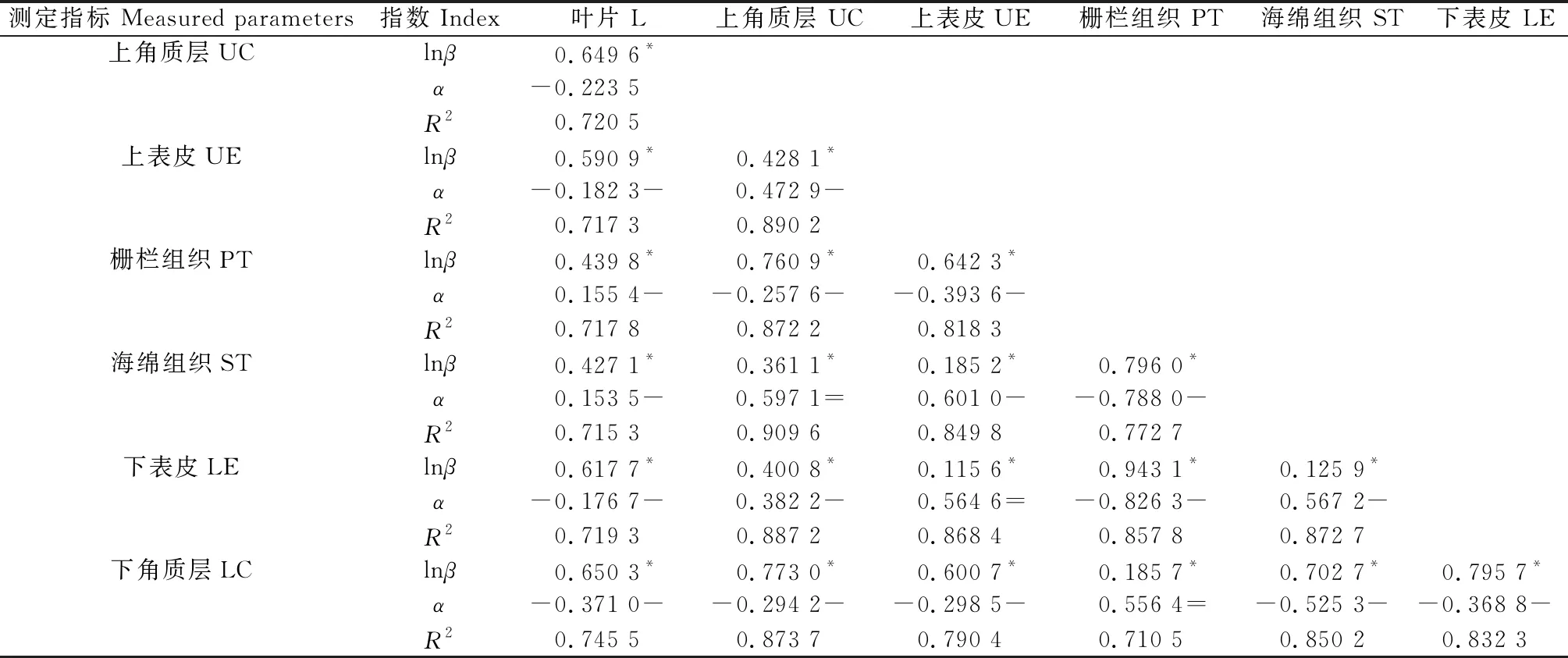
表5 不同生境唐古特白刺叶片性状异速生长方程参数Table 5 Parameters of allometric growth equations of leaf traits in different habitats of Nitraria tangutorum Bobr.
2.3 不同生境大果白刺叶片性状的异速生长规律
不同生境下大果白刺叶片解剖性状的变异系数在0.12~0.39之间(表4),根据变异系数的划分等级,3个不同居群大果白刺叶片解剖结构的变异性为中等变异,其中栅栏组织变异系数最小,上角质层变异系数最大。
以大果白刺叶片解剖指标为分组变量,用异速方程描述叶片解剖性状之间的关系,结果表明:所有21组性状之间关系显著相关(P<0.05)(表6),其中叶片厚度与栅栏组织厚度、上角质层厚度与栅栏组织厚度、上表皮与栅栏组织厚度、上表皮厚度与海绵组织厚度、栅栏组织厚度与海绵组织厚度、上角质层厚度与下表皮厚度、上角质层厚度与下角质层厚度、上角质层厚度与海绵组织厚度、上表皮厚度与下表皮厚度、栅栏组织厚度与下角质层厚度之间的生长指数(P<0.05)与1差异显著并都小于1,Y的线性增加比X缓慢,说明这些性状之间存在异速生长,并且呈现非等比例增长,生长速率前者小于后者,根据异速生长系数值判断标准(表3),这些生长指数之间属于负异速生长关系。除此之外,其余所有的2个解剖指标厚度之间的生长指数(P<0.05)包含1,说明这些性状可能会以相同的速率生长,根据异速生长系数值判断标准(表3)这些生长指数之间属于同速生长关系。决定系数(R2)位于0.449 7~0.898 4之间。
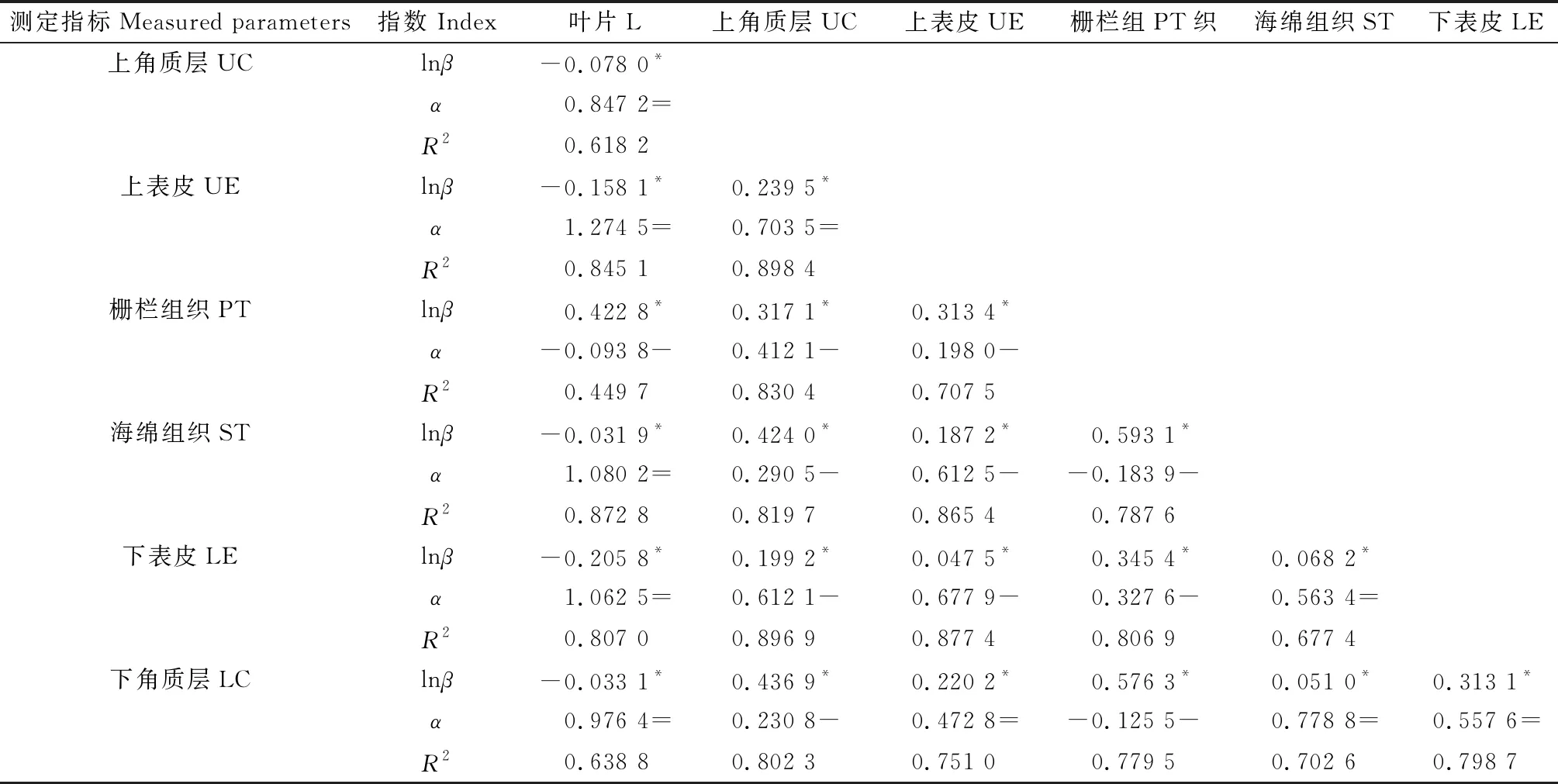
表6 不同生境大果白刺叶片性状异速生长方程参数Table 6 Parameters of allometric growth equations of leaf traits in different habitats of Nitraria roborowskii Kom.
2.4 不同生境西伯利亚白刺叶片性状的异速生长规律
不同生境下西伯利亚白刺叶片解剖性状的变异系数在0.14~0.45(表4)之间,不同生境西伯利亚白刺叶片解剖结构的变异性为中等变异,其中叶片厚度变异系数最小,下表皮厚度变异系数最大。
以西伯利亚白刺叶片解剖指标为分组变量,用异速方程描述叶片解剖性状之间的关系,结果表明:所有21组性状之间关系显著相关(P<0.05)(表7),其中叶片厚度与下表皮厚度、海绵组织厚度叶下表皮厚度、上表皮厚度与下角质层厚度、上表皮与栅栏组织厚度、上表皮厚度与海绵组织厚度、栅栏组织厚度与海绵组织厚度、上角质层厚度与下表皮厚度、上表皮厚度与下表皮厚度、栅栏组织厚度与下表皮厚度之间的生长指数(P<0.05)与1差异显著并都小于1,Y的线性增加比X缓慢,说明这些性状之间存在异速生长,并且呈现非等比例增长,生长速率前者小于后者,根据异速生长系数值判断标准(表3),这些生长指数之间属于负异速生长关系。除此之外,其余所有的2个解剖指标厚度之间的生长指数(P<0.05)包含1,说明这些性状可能会以相同的速率生长,根据异速生长系数值判断标准(表3)这些生长指数之间属于同速生长关系。决定系数(R2)位于0.521 8~0.954 3之间。
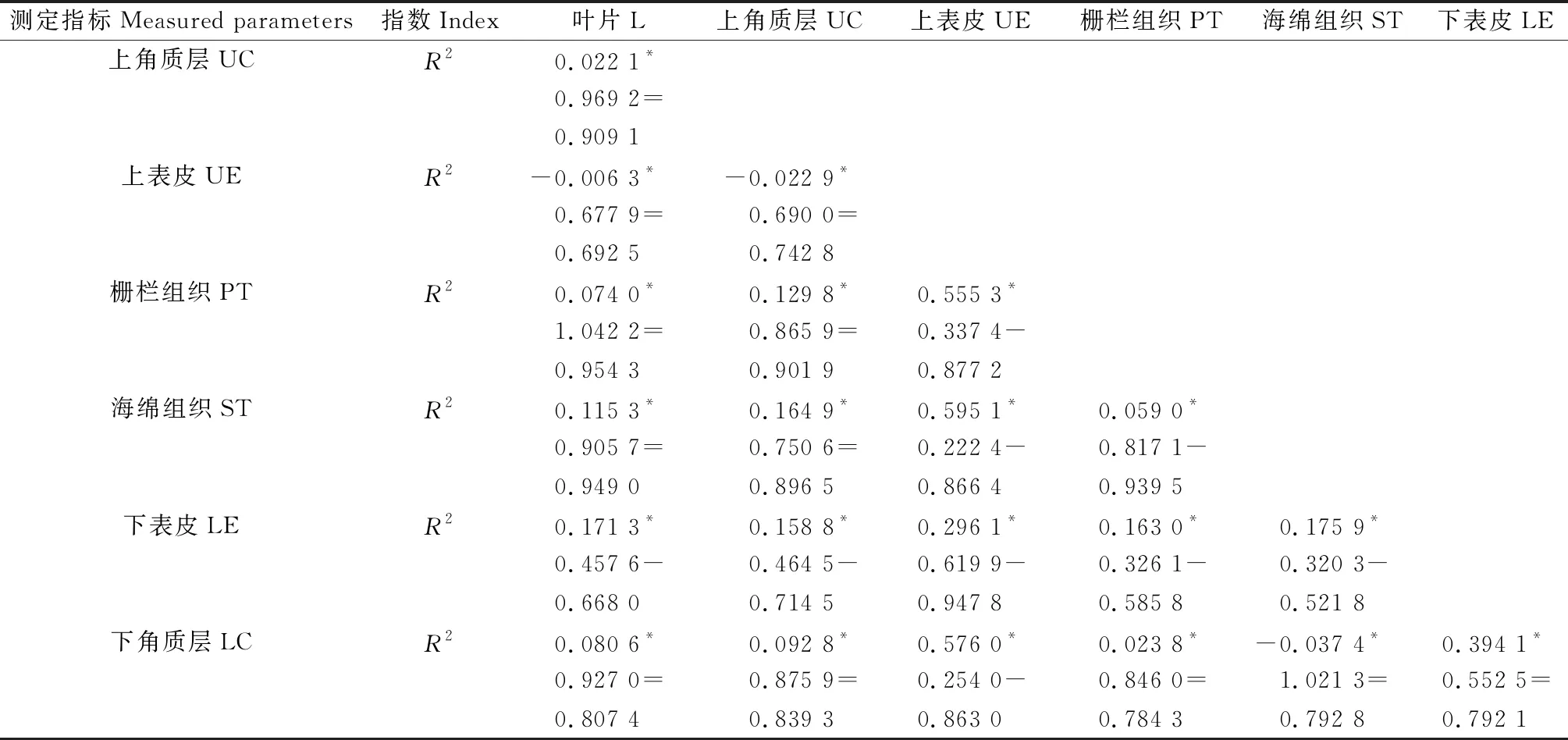
表7 不同生境西伯利亚白刺叶片性状异速生长方程参数Table 7 Parameters of allometric growth equations of leaf traits in different habitats of Nitraria sibirica Pall.
2.5 不同生境3种白刺科植物叶片解剖特征与环境因子之间的关系
对不同生境白刺科植物的7个叶片解剖指标(上角质层厚度、下角质层厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度及叶片厚度)与相应的地理气候生态因子(平均气温、平均日照、年降水量、年蒸发量、纬度、经度与海拔等)进行线性回归分析,结果表明:叶片、栅栏组织、海绵组织与平均气温、平均日照、年降水量存在极显著的正相关变化关系,上表皮和下表皮与平均气温、平均日照、年降水量存在较弱的负相关关系。叶片、栅栏组织、海绵组织与年蒸发量存在极显著的正相关变化关系,上表皮、下表皮和下角质层与年蒸发量存在较弱的负相关变化关系。上角质层、栅栏组织、下表皮、下角质层与纬度存在极显著的负相关变化关系。叶片、栅栏组织、海绵组织与经度存在极显著的正相关变化关系,但上表皮和下表皮与经度存在极显著的负相关变化。叶片、上角质层、上表皮、栅栏组织、海绵组织、下角质层与海拔高度存在极显著的正相关变化关系(表8)。
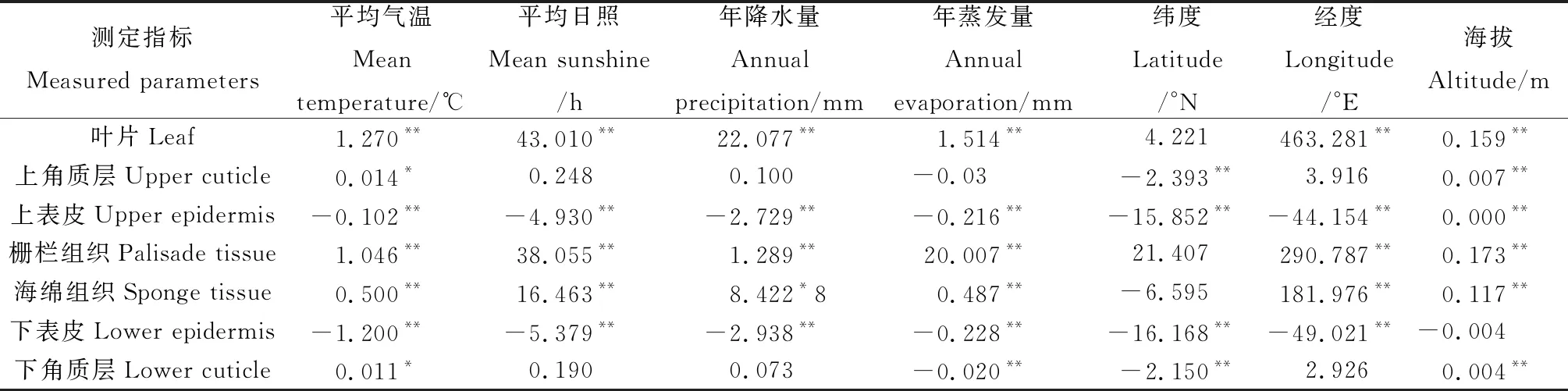
表8 不同居群3种白刺科植物叶片解剖结构参数与环境因子的线性回归分析Table 8 The linear regression analysis of leaf anatomical structure parameters and environmental factors of three Nitrariaceae plants in different populations
3 讨论
叶片解剖结构可以反映外界环境因子对植物的影响,以及植物自身对环境的适应[28]。国内外学者已通过叶表皮解剖特征对不同植物进行系统分类学研究[29]。本实验所有白刺科植物叶片解剖结构均可分为表皮、叶肉与叶脉3部分,为等面叶,等面叶是指叶片上、下两面都有栅栏组织,属于旱生植物的特征。白刺叶片在上下表皮处有2~3层长柱形的栅栏组织,细胞排列紧密,胞间隙较大。对于叶片解剖而言,变异系数大于50%的性状被认为是生态适应性状,变异系数较小的被认为是相对稳定的系统演替性状[2,30],本实验所测量计算的叶片解剖指标的变异系数均在10%~45%之间,基本属于系统演替性状。植物不同性状间的生长关系为异速生长关系是普遍承认的生长规律[31-32]。在本研究中不同生境白刺叶片解剖性状之间存在典型的异速生长关系,可以更好的对进行植物叶片进行功能性状评价,同时表明白刺叶片解剖性状对不同生境具有较强的适应性,其异速生长规律遵循幂函数关系,异速系数为Y轴上的截距,是异速生长起始点的表征,而斜率异速指数可以反映叶片性状对环境的敏感性[33]。
3.1 同种白刺叶片解剖性状异速生长关系
3.1.1叶片厚度与栅栏组织厚度 叶片厚度通常被认为是一个非常有价值的性状,可能与资源获取、水分保存和同化作用有关[22]。蔡永立等对冈青叶片的研究显示,叶片厚度与温度呈负相关关系,即随着温度的降低,叶片厚度呈现增加的趋势[30]。荒漠植物具有发达的栅栏组织,可避免强光对叶肉细胞的损伤[34],同时整个叶片厚度或同化枝直径增加直接影响了水分散失效率[6],对可利用的水资源具有生理调节的作用。通过对不同居群下唐古特白刺叶片样本数据进行分析后得出,叶片厚度与栅栏组织厚度之间的异速生长指数小于1,即栅栏组织的厚度增长速率没有叶片厚度增长速率快,这组性状间的异速生长关系为负向,即存在资源分配权衡(trade-off),指生物将资源(物质和能量)不能同时对繁殖、生长、维持等3方面进行分配,因为生物可利用的资源是有限的,投入到某一功能或性状上的增多必然会减少对另一功能性状的投入[35],虽然栅栏组织厚度增加,但经过分析后发现海绵组织增长速率为负向,造成了叶片厚度与栅栏组织厚度之间的异速生长指数小于1的结果。不同居群大果白刺和西伯利亚白刺叶片厚度与栅栏组织厚度之间的异速生长指数等于1,这组性状间的生长关系为同速生长。说明不同种白刺之间对环境适应策略不同,均向有利于自身生长的方式进化。与John等部分研究叶片厚度与栅栏组织厚度间呈等速生长关系,而与其他组织之间均呈异速生长关系具有一致性[19],但John等研究得出叶片厚度与上表皮厚度间相关性不显著,与本研究结果不一致,原因可能是John等研究对象仅仅包括木本与草本,但本研究中白刺科植物均属于灌木。
3.1.2叶片厚度与海绵组织厚度 通过对不同居群的唐古特白刺样本数据进行分析后得出,叶片厚度增长速率比海绵组织的厚度增长速率快,二者之间的异速生长指数小于1,根据表2异速生长系数值判断标准,其异速生长关系为负向,即二者之间存在着资源分配权衡。不同居群的大果白刺和西伯利亚白刺叶片厚度与海绵组织厚度之间的异速生长指数约等于1,说明叶片厚度与栅栏组织厚度同速率增长,这组性状间的生长关系为同速生长,2个性状之间同速生长关系可能与它们的功能特性具有一致性有关。
3.1.3海绵组织厚度与栅栏组织厚度 叶肉是叶片解剖性状变异系数最大的部分,叶肉组织中具有明显的栅栏组织和海绵组织的分化[2]。海绵组织对植物光合作用和水分利用效率的提高有着积极作用。通过对不同居群唐古特白刺、大果白刺、西伯利亚样本数据进行分析后得出,海绵组织厚度的增长速率没有栅栏组织厚度增长的快,二者之间的异速生长指数小于1,根据表2异速生长系数值判断标准,其异速生长关系为负向,即也存在资源分配权衡。与刘艳芳[2]研究的海绵组织增加的幅度大于栅栏组织的增大不一致,这种差异表明,植物为满足自身生长需求达到最优的组织分配,在不同环境条件下不同类型的植物叶片的栅栏组织与海绵组织的分配策略方面存在明显的差异。
3.2 不同种间白刺叶片解剖性状异速生长关系
有学者研究胡杨叶片解剖结构时发现,所有叶片解剖结构指标均与土壤含水量无显著相关性[29],且由于研究区域均属于干旱荒漠区,土壤含水量比较低,故本实验没有考虑土壤含水量对植物叶片解剖结构的影响。本实验采用完全嵌入拉普拉斯近似方法(INLA)进行近似贝叶斯推断,其原因是采用拉普拉斯近似方法(INLA)可以包含1,即在实验中可以检验1(α=1)。植物异速生长分配理论认为,各器官一般按照异速生长的关系进行发育,异速生长依赖于物种,在物种内异速生长是相对稳定的,产生异速生长的原因是物种对不同环境适应的特异性。Price等通过贝叶斯分析,发现植物的异速指数是变化的[36],因为本实验中3种植物都属于同一科,亲缘关系相对比较近,并且由于不同植物种在形态结构上往往以不同的形式去适应于大致相同的环境条件,同种植物由于长期适应于不同的生境,其形态结构表现出明显的趋异现象[37],一般植物朝着最佳的有利于自身生存的异速生长曲线进化[38]。同种或不同种白刺科植物解剖性状等速或异速生长是植物体在特定生境下所采取的生态对策,反映了植物叶片解剖性状之间的相互关联程度[39]。
3种白刺不同居群的叶片解剖性状均有显著性差异(P<0.05),不同居群下白刺科植物叶片解剖结构性状均呈显著的幂函数异速生长关系且异速系数呈现显著性差异(P<0.05),根据表2异速生长系数值判断标准,异速指数表可分两大类,一类是≈1,即叶片在不同生境下各性状生长速率基本一致,呈等速生长;另一类是<1,即叶片在不同生境下性状呈负异速生长,即叶片解剖性状之间存在权衡关系[40]。权衡策略可以更好地理解植物在资源分配上的外观可塑性和表型可塑性[41]。异速生长的原因可能通过细胞优化生长理论可以解释,当某物种仅仅通过增加细胞数量实现个体生长时,个体的新陈代谢速率和其个体大小成正比(即幂指数为1的比例生长),而当其仅仅通过增加体内每个细胞的大小实现生长时,则呈幂指数异速生长[42]。
同种白刺由于长期不在相同生境下产生了变异,而不同种白刺在同一生境下长期共存,产生了一致的叶片解剖性状,这与白潇等研究相一致:生长在外界环境基本相同的不同种植物,其形态解剖结构往往表现趋同现象,但生长在环境条件差异较大的同一种植物,其形态解剖结构常表现趋异现象[43]。植物某些形态解剖结构对不同环境因子的响应具有一定的相似性,可能与环境因子的综合效应有关[44]。Huang研究发现,在种间和种内水平综合分析生态学代谢理论(基于生物个体大小和环境温度对个体新陈代谢过程的影响,使用尺度推移(scaling)的方法来解释多种生态学格局和过程)的假设与预测,表明生态代谢理论不符合种间水平的结果,但符合种内水平的结果,这是由于在种内水平不同物种具有不同的异速系数,导致种间水平的异速指数发生偏移[45]。相同性状之间生长速率的差异,除环境资源状况不同原因之外,可能由于植物本身的个体差异性造成的,植物在特定的环境下会采取不同的适应可塑性对策增加其生存适合度[46]。
4 结论
在不同生境下的白刺叶片解剖性状均有显著性差异(P<0.05),白刺叶片解剖性状均呈显著的幂函数异速生长关系,其异速系数均表现显著性差异(P<0.05),其异速指数表现两类完全不同的规律,一类是≈1,即叶片解剖性状的生长速率大致相同;另一类是<1,即叶片解剖性状呈负异速生长规律,存在着资源分配权衡。白刺叶片解剖性状的变异系数均在10%~45%之间,基本属于相对稳定的系统演替性状。生长在环境条件差异较大的同一种白刺,其形态解剖结构常表现趋异现象,白刺通过长期的外界环境因子的驱动改变叶片结构性状可以使其在不同的生境下朝着最佳的异速生长曲线进化。研究证明海拔、降水、蒸发量是本研究影响叶片性状的主要因素。
猜你喜欢
数学小灵通(1-2年级)(2021年10期)2021-11-05 07:20:38
落叶果树(2021年6期)2021-02-12 01:29:10
农学学报(2018年10期)2018-11-01 01:19:30
中成药(2018年7期)2018-08-04 06:04:06
数学小灵通(1-2年级)(2017年9期)2017-10-13 08:10:11
西南农业学报(2016年5期)2016-05-17 05:42:28
体育科技(2016年2期)2016-02-28 17:06:07
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
第二课堂(小学版)(2014年4期)2014-08-02 17:39:17
东北林业大学学报(2012年1期)2012-06-13 06:20:40
