
盐胁迫对3种白刺渗透调节物质的影响1)
2012-06-13 06:20:40倪建伟张华新
东北林业大学学报 2012年1期
武 香 倪建伟 张华新 刘 涛 张 丽
(中国林业科学研究院,北京,100091)
土壤盐渍化是一个世界性的重大资源问题和生态问题。全世界盐碱地面积近10亿hm2,约占陆地面积的7.6%。我国各类盐碱地面积0.346亿hm2,仅海涂土壤就占海岸带土壤总面积的17.35%,是世界盐碱地大国[1]。由于工业污染加剧、灌溉农业的发展和化肥使用不当等原因,次生盐碱化土地面积还在扩大[2]。筛选和培育耐盐植物用以改良盐碱地环境与其他改良盐碱地方法相比更为切实可行,因而研究植物的耐盐机理和耐盐能力、培育筛选优良耐盐碱植物具有重要的理论和现实意义。植物在盐渍环境中会表现出一系列的生长和生理反应,以提高自身的抗性或减轻盐胁迫产生的伤害,不同植物由于其耐盐能力的差别,对盐胁迫的响应也不尽相同。目前,树木耐盐性研究主要集中在不同树种对盐胁迫的响应、树木耐盐机理、耐盐树种筛选与培育、耐盐基因工程等方面[3-9],对于同一属的不同种植物对盐胁迫的生长和渗透调节物质含量变化的研究较少。白刺属(NitrariaL.)属蒺藜科(Zygophyllaceae),为多年生落叶小灌木,全世界有12种,中国有8种,主要分布于我国西北部、北部地区[10]。白刺属植物具有耐干旱、耐高温、耐严寒、耐盐碱、耐贫瘠、抗风沙等特性,是用于防风固沙和盐碱地改良的优良植物[11]。本试验以唐古特白刺(Nitraria tangutorumBobr)、西伯利亚白刺(Nitraria sibiricaPall.)、齿叶白刺(Nitraria roborowskiiKom.)为试材,研究其在不同浓度盐胁迫下的生长及渗透调节物质含量变化的差别,综合评价其耐盐能力,为利用白刺改良盐碱地和选育优良耐盐种质资源提供理论依据和指导。
1 材料与方法
试验设计:试验在北京市房山区各庄村的大棚温室内进行。供试材料为唐古特白刺、西伯利亚白刺和齿叶白刺的2年生实生苗,产自吉林白城。2009年5月初,将供试材料定植在30 cm×30 cm的底部带托盘的塑料盆中。定植所用基质为V(腐殖质土)∶V(草炭)∶V(珍珠岩)=1∶3∶1,用多菌灵进行消毒处理。定植后进行常规管理。7月,选择长势相对一致的苗木进行盐胁迫(NaCl)处理。试验采用完全随机区组设计,共4个盐浓度处理,分别为 100、200、300、400 mmol/L,以不加 NaCl为对照,3次重复,每个小区10株苗木。采取分次浇灌的施盐方式。盐胁迫期间,定期定量浇水,漏于花盆托盘中的盐溶液或水倒回盆中,防止盐分流失。盐胁迫处理历时45 d。
测定指标与方法:盐胁迫结束后,每小区选择长势中等的苗木9株,在105℃下,杀青1 h,然后在80℃下烘干至恒质量,进行称量,测定生物量、根冠比,其中,根冠比=地下部分干质量/地上部分干质量。采用硫代巴比妥酸法测定[12]丙二醛,采用蒽酮比色法[13]195-197测 定 可 溶 性 糖,采 用 考 马 斯 亮 蓝法[13]184-185测定可溶性蛋白,采用磺基水杨酸比色法[13]258-260测定脯氨酸。矿质元素参照王宝山等[14]和於丙军等[15]的方法测定,Na+、K+和 Ca2+用原子吸收分光光度计法测定,Cl-用自动滴定仪测定。
数据处理:用SPSS分析软件进行方差分析和多重比较。采用隶属函数法[16],对3种白刺各项耐盐碱指标的隶属值进行累加,求取平均值,用以评定其耐盐性,方法如下:①求出各指标的隶属函数值。如果某一指标与耐盐性呈正相关,则,X(u)=(XXmin)/(Xmax-Xmin);如果某一指标与耐盐性呈负相关,则,X(u)=1-(X-Xmin)/(Xmax-Xmin)。式中:X为各白刺某一指标的测定值;Xmax为3种白刺中某一指标测定值中的最大值;Xmin为该指标中的最小值。②把每种白刺各耐盐性指标的隶属函数值进行累加,计算其平均值。
2 结果与分析
2.1 盐胁迫对3种白刺生长指标的影响
不同浓度NaCl胁迫对3种白刺生物量的影响结果(表1)表明,盐浓度大于200 mmol/L时,唐古特白刺和西伯利亚白刺的生物量与对照差异显著,大于100 mmol/L的盐浓度对齿叶白刺的生物量影响显著(P<0.05)。唐古特白刺、西伯利亚白刺、齿叶白刺在100 mmol/L NaCl胁迫下的生物量比对照分别增加了 4.57%、5.56%、5.61%。而其他盐浓度胁迫下植株的生物量与对照相比都有所降低,盐浓度为400 mmol/L时生物量最少,齿叶白刺生物量减少幅度最大,为24.54%,西伯利亚白刺次之,为23.43%,唐古特白刺减少幅度最小,为22.60%。NaCl胁迫45 d对3种白刺根冠比的影响各不相同(表1)。唐古特白刺在盐浓度为400 mmol/L时与对照差异显著(P<0.05),盐胁迫后的根冠比均小于对照,西伯利亚白刺和齿叶白刺各处理间根冠比差异不显著(P>0.05),其中,西伯利亚白刺则随着盐浓度的增加而降低。齿叶白刺在盐浓度为100 mmol/L时略下降,之后随着盐浓度的增加而增加。

表1 不同浓度盐胁迫对3种白刺生物量、根冠比的影响
2.2 盐胁迫对3种白刺渗透调节物质的影响
2.2.1 对有机渗透调节物质的影响
经45 d盐胁迫后,3种白刺MDA质量摩尔浓度的变化规律不同(表2),盐浓度为100、200 mmol/L时唐古特白刺相比对照差异显著,西伯利亚白刺白刺仅在100 mmol/L时比对照显著降低,齿叶白刺盐胁迫后的MDA质量摩尔浓度显著高于盐胁迫前(P<0.05)。唐古特白刺和西伯利亚白刺在盐浓度为100 mmol/L时MDA质量摩尔浓度较低,分别比对照减少了34.00%、19.55%,盐浓度大于100 mmol/L时其质量摩尔浓度增加。齿叶白刺的MDA质量摩尔浓度随着盐度的增加呈现先升后降的趋势,在盐浓度为300 mmol/L时最大,是对照的1.07倍。低浓度盐胁迫对唐古特白刺和西伯利亚白刺的可溶性糖质量分数的影响显著,齿叶白刺在高盐浓度下相比对照显著增加(P<0.05)(表2),除唐古特白刺在盐浓度200 mmol/L时低于对照外,其它处理均高于对照。西伯利亚白刺和齿叶白刺在盐浓度为200 mmol/L时的可溶性糖质量分数最大,分别比对照增长了45.89%和61.31%,唐古特白刺在盐浓度为100 mmol/L时最大,增长了21.07%。3种白刺受盐胁迫后,可溶性蛋白质量分数均高于对照,盐胁迫对齿叶白刺影响不显著(P>0.05),唐古特白刺盐胁迫后的质量分数显著高于对照,西伯利亚白刺除在100 mmol/L时与对照差异不显著外,其他浓度盐处理与对照相比差异显著(表2)。唐古特白刺和西伯利亚白刺随盐浓度的增加而增加,在盐浓度400 mmol/L时可溶性蛋白质量分数最高,分别比对照增加了95.30%和34.75%。齿叶白刺可溶性蛋白的最大值在盐浓度为200 mmol/L时,比对照增加了67.00%,当盐浓度大于200 mmol/L时虽相比200 mmol/L时有所下降但依然高于对照。盐胁迫45d后,3种白刺的脯氨酸质量分数变化有所不同(表2)。唐古特白刺和西伯利亚白刺盐胁迫后脯氨酸质量分数均高于对照,唐古特白刺除在100 mmol/L时与对照差异不显著(P>0.05)外,其他浓度盐处理均显著高于对照,呈明显的增加趋势。盐胁迫对西伯利亚白刺的影响显著(P<0.05)。齿叶白刺各盐浓度处理间的脯氨酸质量分数变化不显著(P>0.05)。当盐浓度为400 mmol/L时唐古特白刺和西伯利亚白刺的脯氨酸质量分数达到最高,分别增长了82.49%和45.46%,齿叶白刺在200 mmol/L时增长4.66%达到最高。
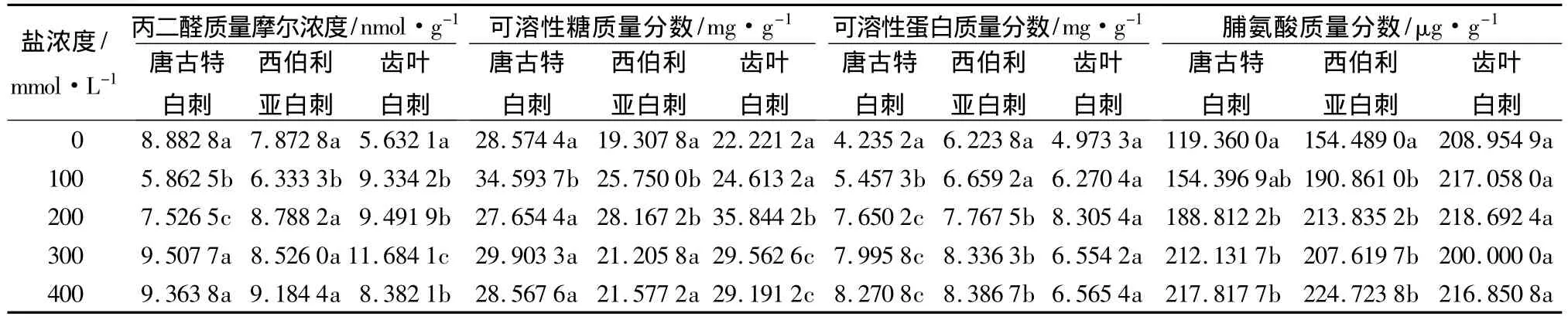
表2 不同浓度盐胁迫对3种白刺丙二醛、可溶性糖、可溶性蛋白、脯氨酸的影响
2.2.2 对3种白刺矿质元素质量分数的影响
由表3可见,盐胁迫后3种白刺叶片中Na+质量分数变化趋势不同。唐古特白刺仅在300 mmol/L时与对照差异显著,为69.1625 mg/g,达到最大值,其他处理与对照差异不明显(P>0.05);西伯利亚白刺的Na+质量分数盐胁迫后变化显著(P<0.05),均高于对照,且随盐浓度的增加呈明显的先增后减趋势,在盐浓度为200 mmol/L时质量分数最大,增加了39.47%。齿叶白刺各处理间差异不显著(P>0.05),在100 mmol/L时齿叶白刺增加了3.54%,达到最大值。盐胁迫对西伯利亚白刺叶片中K+质量分数影响不显著(P>0.05),唐古特白刺在盐浓度大于 100 mmol/L时与对照差异显著(P<0.05),齿叶白刺除200 mmol/L时差异不显著外,其他处理均比对照显著增加(P<0.05)。其中唐古特白刺的K+质量分数随着盐浓度的增加先降低后增加,盐浓度为400 mmol/L时质量分数最大,增加了22.60%。西伯利亚白刺盐胁迫后的K+质量分数均低于对照,盐浓度为400 mmol/L时最小,降低了14.18%。齿叶白刺盐胁迫后的K+质量分数均高于对照,在100 mmol/L时增加了43.54%,达到最大。盐胁迫后,3种白刺叶片中的Ca2+质量分数除唐古特白刺在200 mmol/L时与对照差异不显著外,其他处理均变化显著(P<0.05);唐古特白刺在盐浓度为200 mmol/L时的Ca2+质量分数高于对照外,其它均低于对照。唐古特白刺和西伯利亚白刺在盐浓度为400 mmol/L时Ca2+质量分数最低,分别下降了15.59%和45.93%。齿叶白刺在盐浓度为300 mmol/L时最低,下降了45.95%,400 mmol/L时略有升高。胁迫后,3种白刺叶片中的Cl-质量分数均高于对照,盐胁迫对唐古特白刺和西伯利亚白刺影响显著(P<0.05),齿叶白刺在低盐浓度下无明显变化,在高盐浓度下与对照差异显著(P<0.05)。唐古特白刺和齿叶白刺在300 mmol/L时Cl-质量分数最高,分别增长了20.29%和30.56%,盐浓度大于300 mmol/L时下降,西伯利亚白刺在200 mmol/L时Cl-质量分数最高,增长了31.54%,盐浓度大于200 mmol/L时下降。盐胁迫45d后,3种白刺叶片中的钾钠比变化各不相同。唐古特白刺呈明显的先降后升趋势,在400 mmol/L时钾钠比比对照增长了10.87%,达到最大,其他盐浓度处理均低于对照。西伯利亚白刺盐胁迫后的钾钠比均低于对照,在盐浓度200 mmol/L时钾钠比最小,为0.32。齿叶白刺盐胁迫后的钾钠比均高于对照。
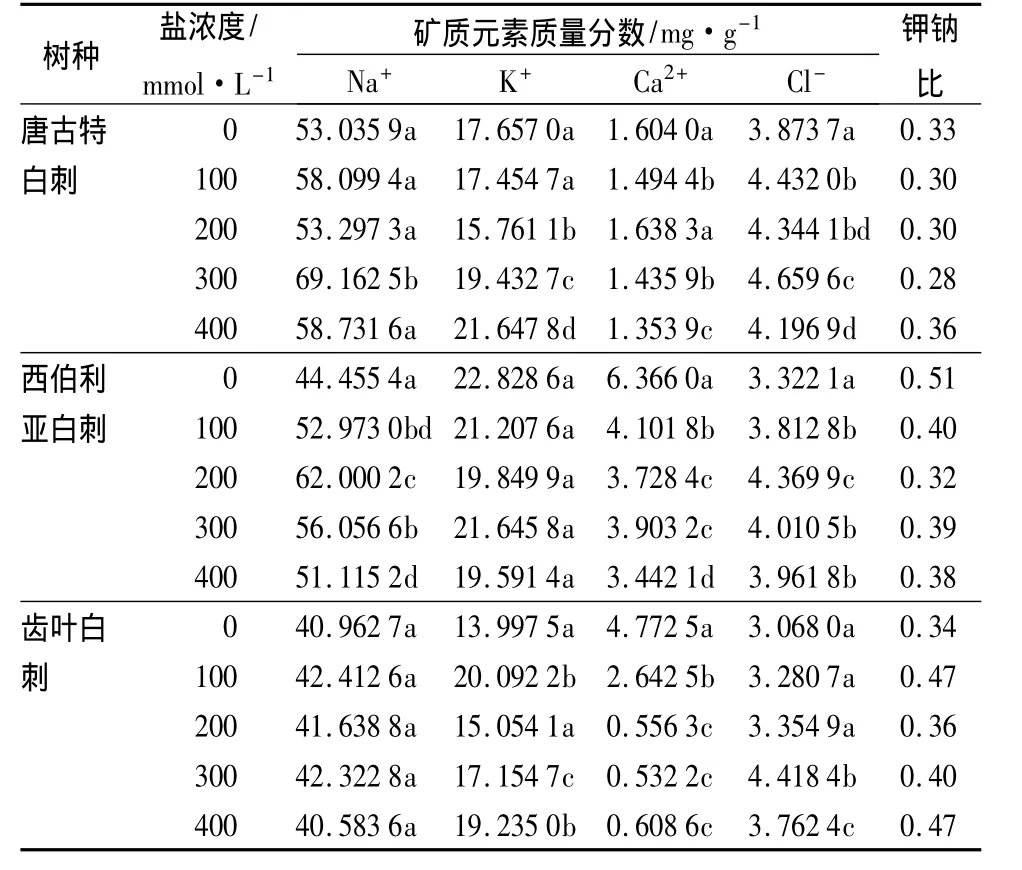
表3 不同浓度盐胁迫对3种白刺Na+、K+、Cl-、Ca2+质量分数和钾钠比的影响
2.3 3种白刺耐盐性的综合评价
以生物量、根冠比、丙二醛、脯氨酸、可溶性蛋白、可溶性糖、Na+、K+、Ca2+、Cl-、钾钠比 11 个指标对3种白刺进行综合评价(表4),结果表明,西伯利亚白刺的隶属度最大,为0.604 0,唐古特白刺次之,为0.466 6,齿叶白刺最小,仅为 0.395 6。故 3 种白刺耐盐能力由强到弱依次为:西伯利亚白刺、唐古特白刺、齿叶白刺。

表4 3种白刺的耐盐性综合评价结果
3 结论与讨论
植物的生长过程对盐胁迫非常敏感,生物量是评估盐胁迫程度和植物抗盐能力的可靠标准[17],高浓度盐胁迫会抑制盐生植物根、茎的生长,改变根系的形态学特征[18],因此盐胁迫下根冠比的变化对植物的耐盐性具有重要意义。本试验中,3种白刺生物量在低盐浓度下略有上升,高盐浓度下降,可见低盐胁迫没有对白刺的生物量积累造成实质性的伤害,高盐胁迫明显抑制了白刺的正常生长。在高浓度盐胁迫下,唐古特白刺和西伯利亚白刺的根冠比小于对照,齿叶白刺大于对照,显然高盐胁迫对唐古特白刺和西伯利亚白刺根的伤害较大,对齿叶白刺地上部分的伤害大于根。
盐渍条件下,植物都要受到渗透胁迫的伤害,它们只有通过在细胞液中积累无机离子或合成有机溶质等方式进行渗透调节以减轻或避免伤害[19],参与渗透调节的有机溶质主要有氨基酸及其衍生物、有机酸类、可溶性碳水化合物类和糖醇类[20]。盐胁迫后,唐古特白刺和西伯利亚白刺的可溶性蛋白质量分数随盐浓度增加而增加,齿叶白刺变化不明显。唐古特白刺除在100 mmol/L盐浓度时可溶性糖质量分数明显高于对照外,其它处理与对照无显著差异,西伯利亚白刺和齿叶白刺呈先增后降趋势。唐古特白刺和西伯利亚白刺盐胁迫下的脯氨酸质量分数有所增加,齿叶白刺无明显变化。可见不同种类白刺在不同盐浓度胁迫下用于渗透调节的主要有机渗透调节物质不同,其变化趋势也有所差别。
决定林木耐盐能力的关键之一在于林木对Na+、K+、Cl-等离子的吸收调节能力,即限制 Na+和Cl-进入体内,选择性吸收K+,才能提高耐盐能力,所以叶片中Na+和Cl-的质量分数是反映林木耐盐特性的良好指标[21]。在本试验中,唐古特白刺和西伯利亚白刺盐胁迫后的Na+质量分数增加,齿叶白刺盐胁迫前后变化不显著。唐古特白刺高盐浓度下的K+质量分数明显高于对照,西伯利亚白刺变化不显著,齿叶白刺各盐浓度处理后均高于对照。盐胁迫后,3种白刺叶片中积累Cl-。Ca2+质量分数除唐古特白刺在200 mmol/L盐胁迫时略高于对照外,其他均低于盐胁迫前。可见矿质元素含量变化对白刺适应盐胁迫环境有重要意义。唐古特白刺除400 mmol/L外的其他盐浓度和西伯利亚白刺的全部盐浓度下的钾钠比均比盐胁迫前小,齿叶白刺相比对照呈现不同程度的增加。这可能是由于种间差别,植物对盐的耐受程度和根系对离子的选择性吸收不同造成的。
渗透调节是植物抵御盐胁迫的重要生理机制,渗透调节物质复杂多样,各种渗透调节物质对植物耐盐性的重要性存在差别。唐古特白刺所测定的渗透调节物质对其耐盐性均有重要作用;K+相对其他物质反映西伯利亚白刺耐盐性的作用较小;在盐胁迫后的齿叶白刺叶片中,Na+、脯氨酸、可溶性蛋白的质量分数与其他渗透调节物质相比变化不明显,可见不同白刺抵御盐胁迫的的主要渗透调节物质各不相同。目前,还没有统一确定的鉴定指标评价植物的耐盐性,笔者对测定的与白刺渗透调节相关的11个指标对白刺的耐盐性进行了综合评价,结果显示西伯利亚白刺的耐盐性最强,唐古特白刺次之,齿叶白刺最差。但植物的耐盐性是一个复杂的综合性状,除受渗透调节物质的影响外还受多种因素的影响。因此在以后的试验中应尽可能多地考虑到与植物耐盐性相关的性状,以便更好地评价植物的耐盐性。
[1]王越,赵辉,马凤江,等.盐碱地与耐盐碱牧草[J].山西农业科学,2006,34(1):55-57.
[2]史湘华,殷鸣放,赵辉,等.盐碱地与耐盐碱树种的选育[J].中国林副特产,2005(2):60-62.
[3]Yeonghoo K,Joji A,Takuji N,et al.Antioxidative responses and their relation to salt tolerance inEchinochloa oryzicolaVasing andSetaria virdis(L.)Beau[J].Plant Growth Regulation,2004,44:87-92.
[4]Corney H J,Sasse J M,Ades P K.Assessment of salt tolerance in eucalypts using chlorophyll fluorescence attributes[J].New Forests,2003,26(3):233-246.
[5]孙海菁,王树凤,陈益泰.盐胁迫对6个树种的生长及生理指标的影响[J].林业科学研究,2009,22(3):315-324.
[6]张华新,刘正祥,刘秋芳.盐胁迫下树种幼苗生长及其耐盐性[J].生态学报,2009,29(5):2263-2271.
[7]赵可夫,范海.盐胁迫下真盐生植物与泌盐植物的渗透调节物质及其贡献的比较研究[J].应用与环境生物学报,2000,6(2):99-105.
[8]任丽丽,任春明,赵自国.植物耐盐性研究进展[J].山西农业科学,2010,38(5):87-90.
[9]张华新,宋丹,刘正祥.盐胁迫下11个树种生理特性及其耐盐性研究[J].林业科学研究,2008,21(2):168-175.
[10]李双福,张启昌,张起超,等.白刺属植物研究进展[J].北华大学学报:自然科学版,2005,6(1):78-81.
[11]王彦阁,杨晓晖,于春堂,等.白刺属植物现状、生态功能及保护策略[J].水土保持研究,2007,14(3):74-79.
[12]郝建军,康宗利,于洋.植物生理学实验技术[M].北京:化学工业出版社,2007.
[13]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[14]王宝山,赵可夫.小麦叶片中Na、K提取方法的比较[J].植物生理学通讯,1995,31(1):50-52.
[15]於丙军,罗庆云,曹爱忠,等.栽培大豆和野生大豆耐盐性及离子效应的比较[J].植物资源与环境学报,2001,10(1):25-29.
[16]孙彩霞,沈秀瑛.作物抗旱性鉴定指标及数量分析方法的研究进展[J].中国农学通报,2002,18(1):49-51.
[17]Larcher W.植物生态生理学[M].翟志席,译.北京:中国农业大学出版社,1997.
[18]王东明,贾媛,崔继哲.盐胁迫对植物的影响及植物盐适应性研究进展[J].中国农学通报,2009,25(4):124-128.
[19]武维华.植物生理学[M].北京:科学出版社,2003.
[20]王劲,杜世章,刘君蓉.植物耐盐机制中的渗透调节[J].绵阳师范学院学报,2006,25(5):56-61.
[21]汪贵斌,曹福亮,游庆方,等.盐胁迫对4树种叶片中 K+和Na+的影响及其耐盐能力的评价[J].植物资源与环境学报,2001,10(1):30-34.
猜你喜欢
食品与健康(2021年9期)2021-09-10 16:26:04
小读者(2019年20期)2020-01-04 02:13:32
武汉冶金管理干部学院学报(2018年3期)2018-11-15 08:16:06
农学学报(2018年10期)2018-11-01 01:19:30
中成药(2018年7期)2018-08-04 06:04:06
西夏学(2018年1期)2018-04-29 09:09:02
西夏研究(2017年1期)2017-07-10 08:16:55
西南农业学报(2016年5期)2016-05-17 05:42:28
天然产物研究与开发(2014年6期)2014-04-27 14:15:58
探索地理(2013年3期)2014-01-09 06:37:28
