
不同利用方式对大针茅草原生态系统多功能性的影响
2022-06-02 02:01郑佳华赵萌莉
草地学报 2022年5期
郑佳华,赵萌莉*,王 琪,张 峰,张 彬,张 军
(1. 内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010019; 2. 内蒙古农业大学草地资源教育部重点实验室,内蒙古 呼和浩特 010019; 3. 内蒙古农业大学理学院,内蒙古 呼和浩特 010019)
生物多样性与生态系统功能的关系一直是生态学研究的热点问题[1],生态系统的价值在于其能提供多种功能和服务的能力,即生态系统多功能性(Ecosystem multifunctionality,EMF)[2]。土壤养分循环相关指标通常用于评价生态系统多功能性,主要是因为土壤和植被是高度复杂且相关的系统,在很大程度上反映了多种生态系统功能,如碳、氮、磷储存以及支持植物生长的能力[3-4]。虽然已有研究表明植物多样性是影响生态系统多功能性的重要因素[3,5],但非生物因素也起到重要的中介作用,生物多样性对生态系统多功能性的影响可能通过土壤非生物因素传递[6]。目前,不同利用方式下生物和非生物因素对典型草原生态系统多功能性的影响尚不明确,开展此方面的研究对草地生物多样性和生态系统功能的保护以及探索草地可持续利用方式具有重要意义。
人类活动导致的土地利用变化是导致生物多样性丧失和多种生态系统功能改变的重要因素[7]。利用方式的改变可以通过改变土壤理化性质直接影响生态系统过程[8],也可以通过改变植物以及土壤微生物群落结构和多样性间接影响[9]。放牧、刈割和围封是内蒙古草原最常见的草地管理措施[10-11]。放牧通过选择性采食、践踏和家畜的排泄物等多种机制影响植物群落和生态系统功能[12]。一般来说,重度放牧对典型草原生态系统有负面影响,有研究表明,放牧的增加导致优势种重要值降低,同时显著降低了土壤养分含量[13]。汤永康等[14]的研究也发现,土壤含水量、草地蒸散速率、土壤有机碳含量和土壤呼吸速率均与放牧强度呈负相关关系,说明重度放牧降低了典型草原生态系统服务及功能。刈割主要通过非选择性去除植物地上部分影响植物群落和生态系统功能,这可能会增强植物的补偿性生长和土壤养分的矿化,显著增加土壤中的有机碳以及微生物生物量碳和氮含量[15]。也有研究表明,一年一次的刈割是维持内蒙古典型草原土壤肥力和微生物群落功能的最佳措施,有利于维持或改善土壤功能[16-17]。围封是内蒙古恢复退化草地的主要方式[18]。然而,围封可能对草地生态系统功能和服务产生一系列负面影响,其对退化草地的恢复效果以及草地生态系统功能的影响仍然存在争议[19]。因此,研究不同利用方式对生态系统多功能性的影响有重要意义。
大针茅草原作为典型草原最具代表性的类型,长期受到放牧及刈割等人为活动的干扰,对草地生态系统造成了深远的影响,然而,不同利用方式对大针茅草原生态系统多功能性的影响尚不明确。本试验以大针茅草原为研究对象,研究不同利用方式对大针茅草原生态系统多功能性的影响以及植物多样性和土壤非生物因素与生态系统多功能性的关系,为指导当地生产实践以及草地合理利用提供理论依据。
1 材料与方法
1.1 研究区概况
本研究在内蒙古锡林浩特市毛登牧场(116.03°~116.50° E,44.80°~44.82° N)进行。该地区属温带干旱大陆性气候;年平均气温2.25℃,月平均气温1月最低(-19.6℃),7月最高(21.8℃);年平均降水量281.39 mm,大部分降水集中在6~9月,占全年降水量的80%以上;年蒸发量2 000~2 700 mm;光照、热、降水同期出现。大针茅典型草原的土壤类型主要是栗钙土。天然植被以大针茅(Stipagrandis)为建群种,优势种有羊草(Leymuschinensis)和知母(Anemarrhenaasphodeloides),主要伴生种有黄囊苔草(Carexkorshinskyi)、黄花黄芪(Astragalusluteolus)、冰草(Agropyroncristatum)以及冷蒿(Artemisiafrigida)等[20]。
本研究包括的利用方式为放牧(Grazing)、刈割(Mowing)以及围封(Enclosed),各个样地在地形地貌、土壤组分、气候及原生群落组成与类型上基本一致。放牧样地位于毛登牧场的牧民草场,全年自由放牧,主要放牧家畜为绵羊,2014—2020年通过在家庭牧场设置9个移动围笼测定样地利用率为63.5%,属于重度放牧,该样地放牧年限超过20年。其中刈割样地和围封样地于2014年开始围建,采用随机区组试验设计,各3次重复,每个小区面积为50 m×30 m,每年的8月31号使用打草机对刈割样地进行打草并禁牧,留茬高度为5 cm。该试验样地在围建之前与放牧样地的利用方式和强度完全相同。
1.2 植物取样与分析
于2020年8月中旬对不同利用方式下的样地进行了植物群落的采样。在刈割和对照样地各个重复小区内随机设置3个1 m×1 m的样方,在放牧样地内的移动围笼内设置1个1 m×1 m的样方,分种剪取样方中出现的所有物种并记录其高度、盖度和密度,装入信封中带回实验室。将植物地上部分放在65℃的烘箱中烘干48 h至恒重,称重后使用球磨仪磨成草粉,用于测定植物营养元素含量,植物磷含量使用钼锑抗比色法测定,植物氮和植物碳含量使用元素分析仪测定。在每个样方内使用内径7 cm的根钻收集3个0~20 cm的土壤样品计算植物的地下生物量,使用2 mm的筛子冲洗样品,将植物根在65℃烘干48 h后称重,作为地下生物量。
1.3 土壤样品采集与分析
在每个样方中剪去植物地上部分后,使用土钻采集3个0~20 cm的土壤样品,并混合成一个样品。使用2 mm的筛子筛去根和石头,然后分成2份保存运回实验室,一份放在4℃冰盒中用于土壤速效氮的测定,另一份带回实验室风干后用于其他土壤理化性质指标的测定。完成土壤样品采集后,在每个小区内使用快速检测温度计随机测定3处20 cm深度的土壤温度,计算平均值记为该小区的土壤温度。使用烘干称重法测定土壤含水量。
土壤全磷含量使用钼锑抗比色法测定;土壤铵态氮、硝态氮含量使用连续流动分析仪AA3测定;土壤有机碳含量使用重铬酸钾容量法-外加热法测定;土壤有效磷含量使用0.5 mol·L-1NaHCO3法测定;土壤电导率使用电导仪测定;土壤pH使用pH计测定。以上指标均严格按照《土壤农化分析》手册中的实验步骤测定[21]。
1.4 植物多样性计算
本研究中的植物物种多样性指数包括:物种丰富度指数(Margalef指数)、Simpson指数、Shannon-Wiener指数和物种均匀度指数(Pielou指数),计算公式如下:
Margalef指数:M=(S-1)/lnN
Pielou指数:SE=H′/lnS
上式中,S是样方内的物种数目是第i个物种的相对密度,N为样方中所有物种的个体数。
1.5 生态系统多功能性量化
本研究共选择10个生态系统功能指标计算生态系统多功能性,这10个指标分别与碳循环(土壤有机碳含量、植物碳含量),氮循环(土壤铵态氮含量、土壤硝态氮含量、植物氮含量),磷循环(土壤总磷含量、土壤有效磷含量、植物磷含量)以及生产力(地上生物量、地下生物量)有关(表1)。利用单功能法和平均值法量化生态系统多功能性[4,6]。
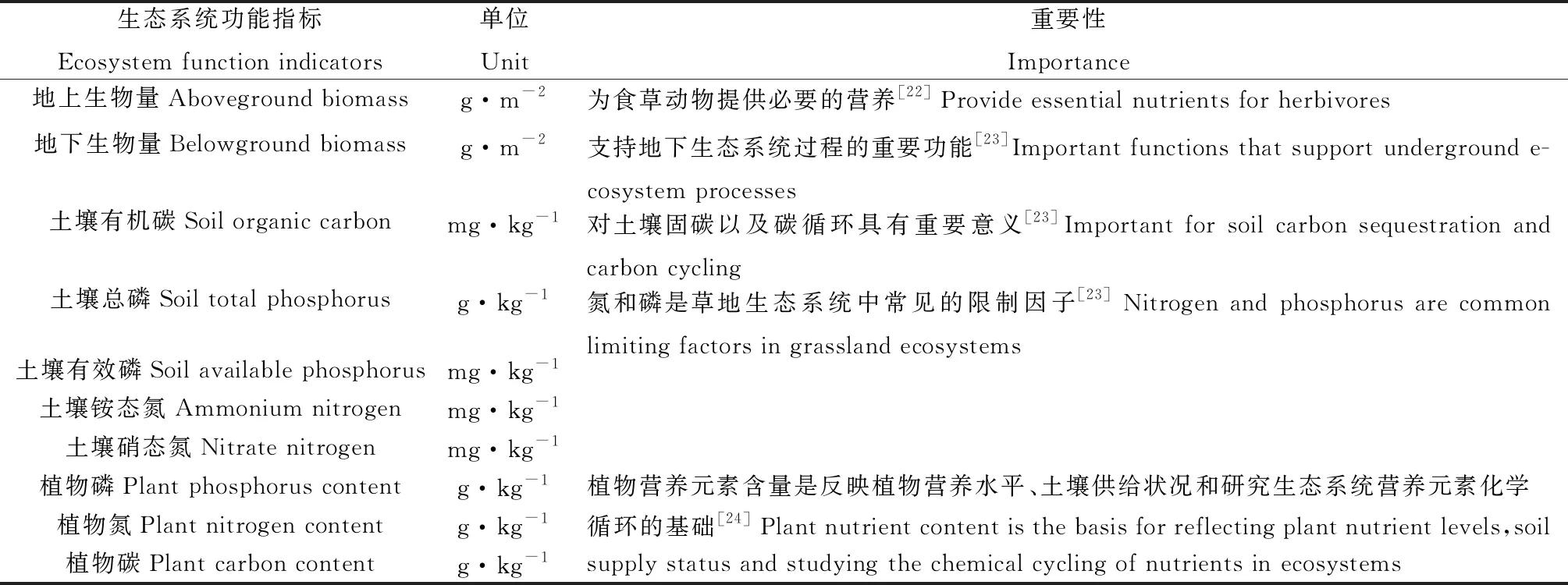
表1 生态系统功能指标及其重要性Table 1 Ecosystem function indicators and their importance
使用Z得分法标准化样方内每个生态系统功能指标,即用测定值减去平均值后除以标准差,具体计算公式为:
Zij=(Xij-ui)/σj
式中,Zij为第i个样方第j种生态系统功能指标的Z得分;Xij为第i个样方第j种生态系统功能指标的测定值;uj为第j种生态系统功能指标在所有样方内的平均值;σj为第j种生态系统功能指标在所有样方内的标准差。
单个功能指数计算公式为:
式中,Fij为第i个样方第j种生态系统功能指标的功能指数;n为该功能包含的生态系统功能指标数量。
生态系统多功能性指数为样方内所有单个功能指数的平均值;单一生态系统功能指数为其所包括的单个功能指数的平均值。即生产力指数为地上生物量和地下生物量的单个功能指数的平均值;碳循环指数为土壤总碳含量、土壤有机碳含量和植物碳含量的单个功能指数的平均值;氮循环指数为土壤总氮含量、土壤铵态氮含量、土壤硝态氮含量和植物氮含量的单个功能指数的平均值;磷循环指数为土壤总磷含量、土壤有效磷含量和植物磷含量的单个功能指数的平均值。
1.6 数据处理
对试验数据进行正态分布检验和方差齐性检验,本试验的数据均符合正态分布且通过了方差齐性检验,采用方差分析法分析不同利用方式对植物多样性及生态系统功能指标的影响,并进行差异显著性检验(Duncan法,α=0.05);采用皮尔逊相关分析法分析单一生态系统功能与植物多样性的关系;采用一元线性回归分析植物多样性和土壤环境因子与生态系统多功能性的关系;建立线性混合效应模型分析生态系统多功能性的主要影响因素。以上数据分析均采用SPSS 23.0进行,采用Origin 2019作图。
2 结果与分析
2.1 利用方式对土壤理化性质及植物多样性的影响
与围封和刈割相比,放牧显著降低了土壤硝态氮、有效磷、有机碳含量,但显著提高了植物磷含量(P<0.05)(表2)。与围封和刈割相比,放牧显著降低了土壤含水量、温度和pH,刈割下土壤pH显著高于围封和放牧(P<0.05)(表3)。
刈割下Shannon-Wiener指数和Margalef指数显著高于放牧,Shannon-Wiener指数表示植物群落的复杂程度,Margalef指数表示植物群落的丰富度,说明刈割导致植物群落丰富度显著增加且群落复杂程度增高。与围封和刈割相比,放牧显著降低了Simpson指数(P<0.05),Simpson指数表示植物群落中优势植物的优势度大小,说明放牧显著降低了群落中优势植物的优势度(图1)。
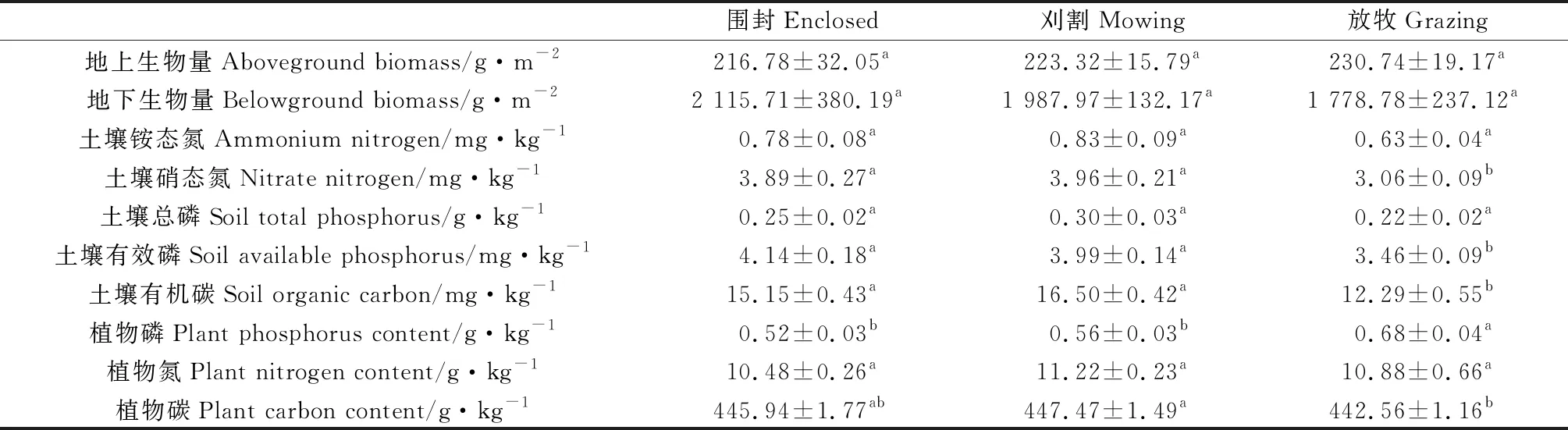
表2 不同利用方式对生态系统多功能性指标的影响Table 2 The effects of different management regimes on the variables used to compute ecosystem multifunctionality
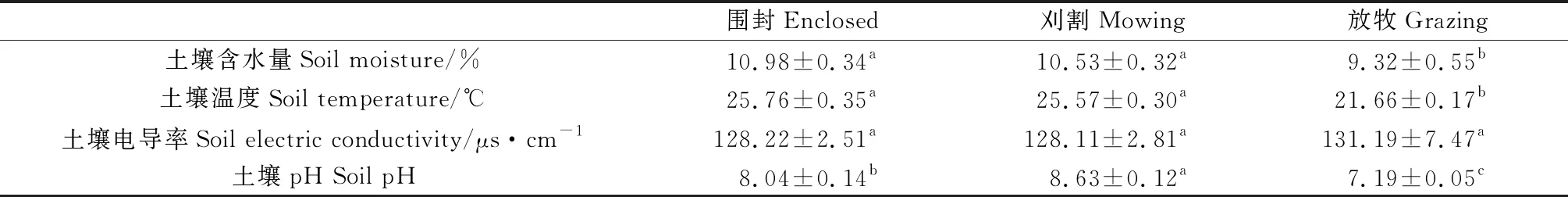
表3 不同利用方式对土壤环境因子的影响Table 3 The effects of different management regimes on the soil environment factor
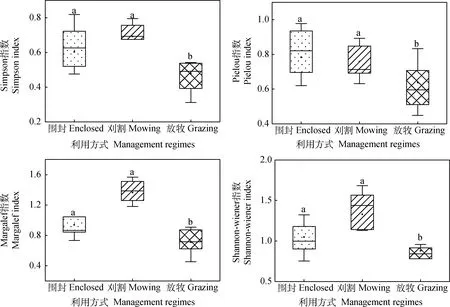
图1 不同利用方式对植物多样性的影响Fig.1 The effects of different management regimes on the plant diversity注:不同小写字母表示处理间差异显著(P<0.05),下图同Note:Diferent small letters indicated significant difference at 0.05 level,the same as below
2.2 利用方式对生态系多功能性的影响
围封和刈割下生态系统多功能性显著高于放牧(P<0.05)(图2)。对于单一生态系统功能而言(图3)围封和刈割下碳循环指数和氮循环指数显著高于放牧(P<0.05)。说明放牧降低了碳循环功能和氮循环功能以及生态系统多功能性。
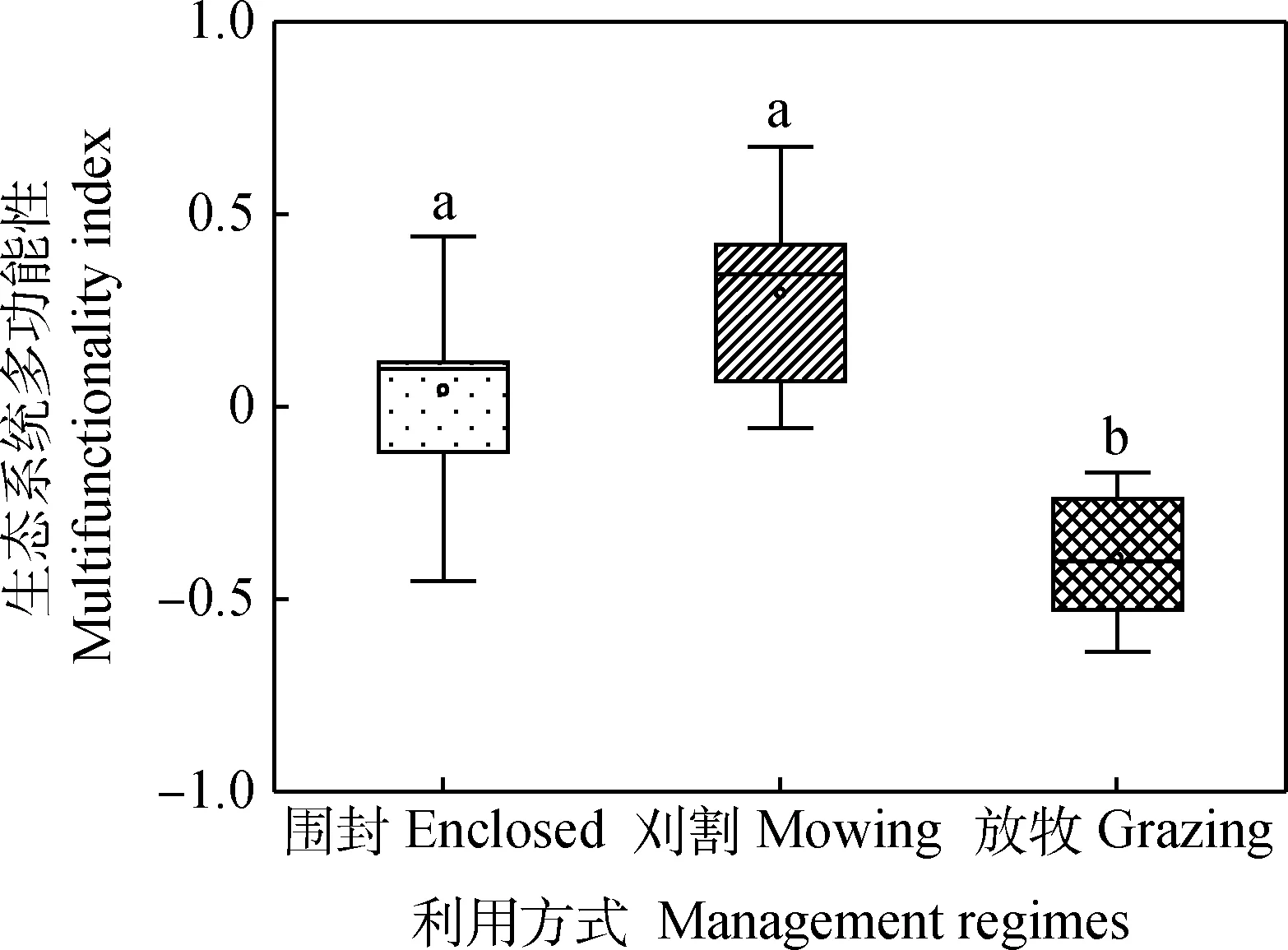
图2 不同利用方式对生态系统多功能性的影响Fig.2 The effects of different management regimes on the multifunctionality index
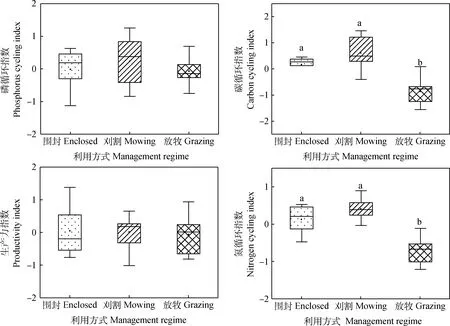
图3 不同利用方式对与C,N,P循环及植物生产力有关功能的影响Fig.3 The effects of different management regimes on the different functional indices related to the C,N,P cycles and plant productivity
2.3 植物多样性与生态系统多功能性的关系
相关分析的结果表明,氮循环指数和植物多样性指数呈显著正相关关系,碳循环指数和Simpson指数及Shannon-Wiener指数呈显著正相关关系(P<0.05)(图4)。植物多样性指数与生态系统多功能性指数线性回归的结果表明,Shannon-Wiener指数、Simpson指数及Pielou指数均与生态系统多功能性呈显著的正相关关系(P<0.05)(图5),其中,Shannon-Wiener指数与生态系统多功能性拟合的方程结果最好,解释量达到了57.5%(P<0.001)。
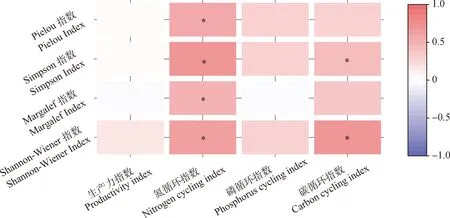
图4 C,N,P循环及生产力指数与植物多样性的关系Fig.4 Relationships between C,N,P cycling and Productivity indices and the diversity of plant注:“*”表示显著相关(P<0.05)Note:" * " indicates significant correlation at the 0.05 level
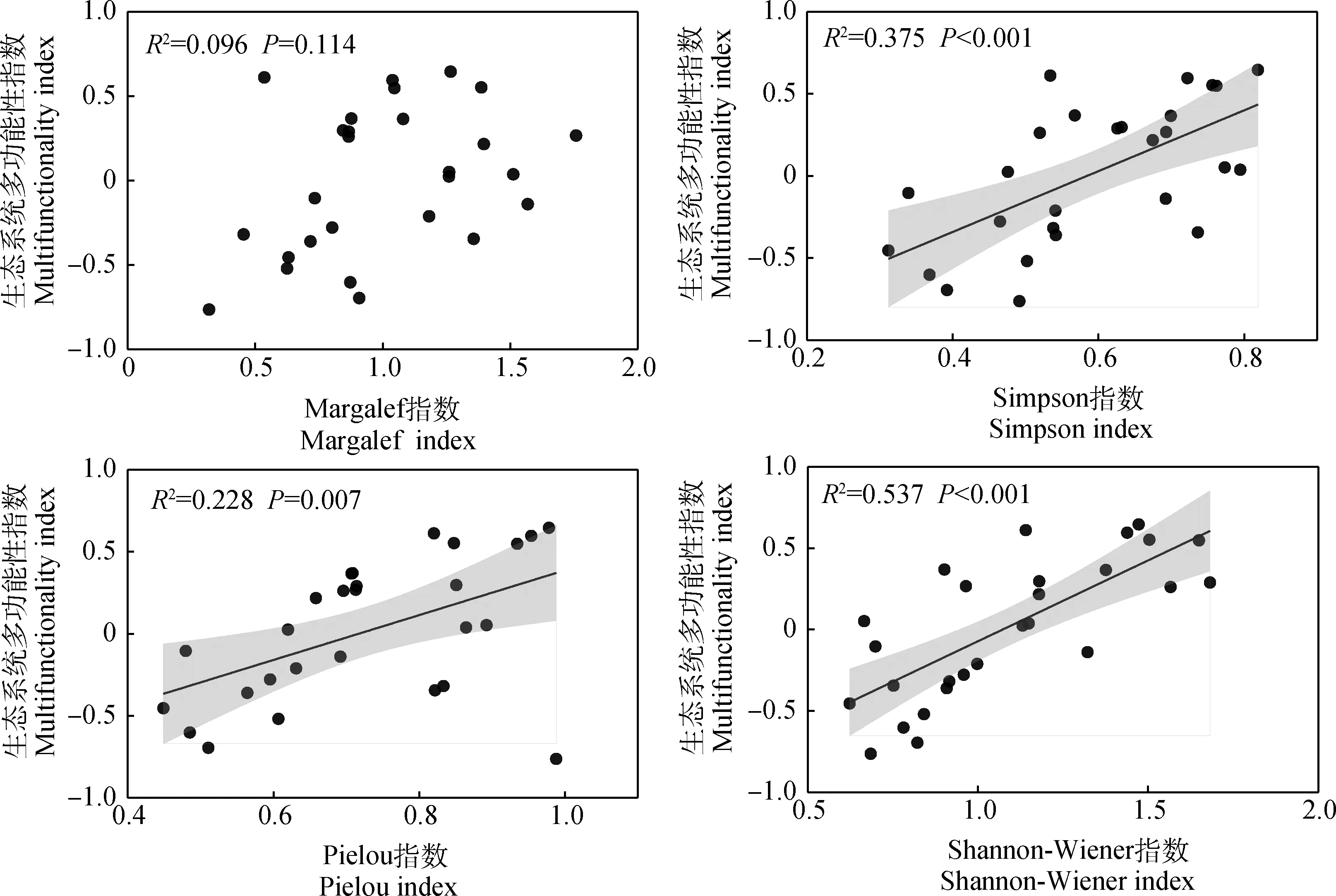
图5 生态系统多功能性指数与植物多样性的关系Fig.5 Relationships between grassland multifunctionality and plant diversity
2.4 生态系统多功能性影响因素
使用植物多样性指数和土壤环境因子与生态系统多功能性建立线性混合效应模型。模型整体R2为0.727 5,说明植物多样性和土壤环境因子共解释了生态系统多功能性72.75%的变化。结果表明,Shannon-Wiener指数对生态系统多功能性有显著的正向影响(P<0.05)。且回归系数绝对值最大(0.722),说明Shannon-Wiener指数对生态系统多功能性的影响最大。同时,Pielou指数(0.481)和土壤含水量(0.093)也对生态系统多功能性有显著的正向影响,此外,土壤pH(-0.312)对生态系统多功能性有显著的负向影响(P<0.05)(表4)。
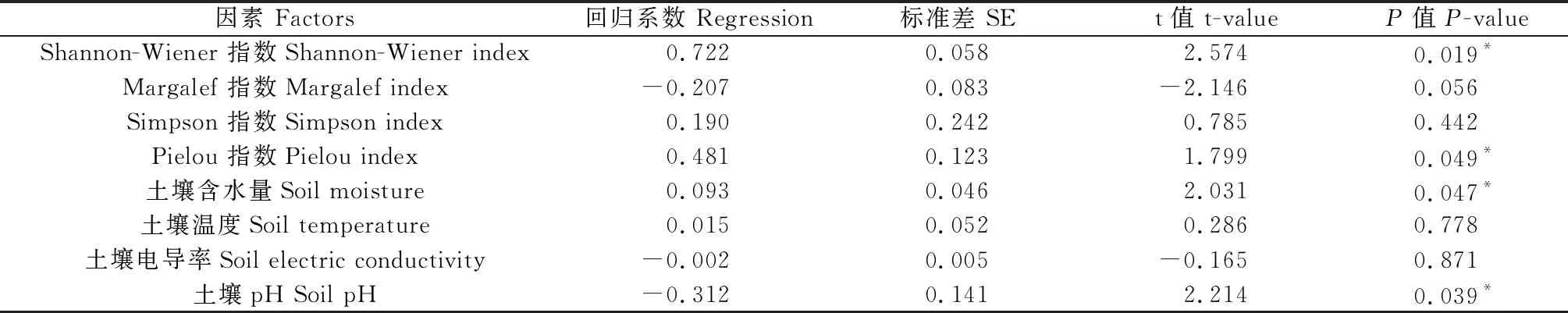
表4 植物多样性和土壤环境因子对多功能性影响的线性混合效应模型Table 4 Summary of the linear mixed-effects model for the effects of plant biodiversity and soil environment factors on grassland multifunctionality
3 讨论
3.1 利用方式对生态系统多功能性和植物多样性的影响
本研究表明,重度放牧显著降低了生态系统多功能性,Zhang等[25]对我国短花针茅荒漠草原的研究表明,重度放牧显著降低了生态系统多功能性,Ren等[26]的研究也表明,内蒙古典型草原生态系统多功能性随放牧压力的增加而下降。上述研究与本研究结果一致,这主要归因于重度放牧条件下碳、氮养分循环显著降低。植物根系和凋落物分解是土壤有机碳的主要来源,一方面,放牧家畜对地上植被持续的采食作用导致植物将更多资源分配到地上部分,导致地下生物量的减少,从而减少了对土壤的碳输入[27];另一方面,放牧导致土壤呼吸作用增强,加速向大气释放二氧化碳[28],进而导致土壤有机碳含量降低。同时,本研究表明,重度放牧显著降低了土壤硝态氮含量(P<0.05),这与顿沙沙等[15]的研究结果一致。可能是由于放牧破坏了土壤表层结构,地表暴露,最终由于风蚀加剧了土壤养分的流失[29],此外,植物主要吸收利用土壤中的硝态氮,放牧刺激下植物对土壤中硝态氮的吸收加剧,导致放牧下土壤硝态氮含量显著降低[30]。本研究表明,围封和刈割间生态系统多功能性以及各生态系统单一功能均无显著差异,这是由于刈割导致仅有很少的枯落物归还于土壤,土壤的资源流失得不到补充,长期围封也导致地面堆积过厚的枯落物,不利于凋落物的分解进而补充土壤养分[31]。同时,有研究表明,与围封相比,刈割下具有更高的土壤养分循环速率,所以有更高的生态系统多功能性[32],这说明与围封和重度放牧相比,刈割可能更有利于维持草地生态系统多功能性。
在本研究中,刈割下植物多样性指数显著高于围封和重度放牧,说明在刈割条件下,植物群落丰富度最大,群落最复杂。刈割移除了地上植被和立枯物,从而增加了光有效性,有利于种子萌发,并且刈割主要收获高大的优势种,为低矮的植物提供生长和繁殖的空间,从而提高植物的多样性和生物量[33-34]。同时,植物多样性和生物量增加也通过增加植物根系、根系分泌物和光合产物的有机碳和氮输入提高了土壤碳和氮储量[18],本研究中土壤碳、氮循环指数与植物多样性指数显著正相关也证明了这一观点。植物多样性的增加导致C4植物种类增加,有利于土壤氮矿化和积累[32],这表明植物多样性对于维持与碳和氮循环相关的生态系统功能,进而维持碳氮固存和土壤肥力尤为重要[35]。此外,放牧显著降低了植物多样性指数,主要是由于本研究的放牧强度属于重度放牧,导致植物低矮,分蘖减少,一些物种大量减少甚至消失,进而导致多样性指数降低。
3.2 植物多样性和土壤环境因子对生态系统多功能性的影响
目前,大量研究表明植物多样性是生态系统多功能性的主要驱动因素,且往往与生态系统多功能性正相关[4,5,32,36],与本研究结果一致。植物多样性指数均与生态系统多功能性呈显著的正相关关系(P<0.05),其中Shannon-Wiener指数的拟合结果最好,解释量达到了57.5%,说明了植物多样性对于维持生态系统多功能性的重要性。这一结果可能与植物群落生态位互补效应有关[37],植物多样性的增加有利于植物群落物种生态位的分化[38],有利于植物更有效地利用生态系统中的资源和空间,进而维持高水平的生态系统多功能性。
本研究表明,土壤含水量对生态系统多功能性有显著的正效应。一项对欧洲森林生态系统的研究也表明,生态系统多功能性存在强烈的环境依赖性,水分可用性等环境因子起到至关重要的作用[39]。较高的土壤水分通常与较高的地上净初级生产力和较高的养分有效性相关,这有利于植物对资源的利用,进而维持更高的生态系统多功能性[10]。此外,土壤pH对生态系统多功能性有显著的负效应。这与Jing等[10]的研究结果一致,土壤酸化会抑制土壤有机碳的分解,从而增强草地生态系统的多种功能[40]。本研究中多元线性回归方程结果表明,当综合考虑植物多样性及土壤环境因子时,生态系统多功能性的解释量达到了72.75%,高于任一单独因素的解释量。因此,在评价草地生态系统多功能性时,必须综合考虑植物多样性和土壤环境因子,以促进草地生态系统的保护和可持续管理。
4 结论
本研究表明,持续重度放牧显著降低了生态系统多功能性,导致草地生态系统功能加剧恶化;长期围封下草地略有恢复;刈割下草地生态系统功能和多样性恢复最佳。植物多样性是生态系统多功能性的主要驱动因素,但是结合土壤环境因子有利于提高对生态系统多功能性的解释量,能够更有效地预测生态系统多功能性。
猜你喜欢
绿色天府(2022年9期)2022-11-19
世界热带农业信息(2022年8期)2022-07-19
纺织标准与质量(2022年1期)2022-07-12
贵州畜牧兽医(2022年3期)2022-06-28
当代水产(2022年4期)2022-06-05
福建轻纺(2022年4期)2022-06-01
中国药学药品知识仓库(2022年8期)2022-05-09
疯狂英语·初中版(2019年4期)2019-09-10
小太阳画报(2018年6期)2018-05-14
青少年日记·小学生版(2009年1期)2009-03-09
