
胡麻生理生化代谢指标对干旱胁迫的响应及其模拟模型的研究
2022-05-30 08:35康亮河
干旱区地理(汉文版) 2022年3期
康 佳, 李 玥, 康亮河
(甘肃农业大学信息科学技术学院,甘肃 兰州 730070)
胡麻是一种具有抗寒、抗旱、耐瘠薄强的作物,又名油用亚麻[1]。在我国,甘肃省种植胡麻面积最大,约10.09×104hm2[2]。水资源不足一直是环境和资源的主要问题,影响作物生长,限制农业生产[3]。国内外就大豆、花生、小麦、油橄榄等[4-7]多种农作物的生理生化代谢指标对干旱胁迫的响应机制已经进行了大量研究。研究表明,由于作物类型多样、自身遗传基础等各种因素,作物适应干旱胁迫的反应在生理变化上存在着明显差异[8]。面对程度不同的干旱胁迫,作物所表现出第一反应特征为植株低矮、叶片数减低、叶面积指数降低和作物产量下降;其中,渗透代谢指标脯氨酸(Pro)含量、可溶性蛋白含量、丙二醛含量均增加;抗氧化代谢指标中的过氧化物酶(POD)、超氧化物歧化酶(SOD)活性增大。王利琴等[9]研究了胡麻不同品种生理特性受干旱的影响,赵利等[10]研究了受水分胁迫影响不同抗旱类型胡麻苗期的生理生化指标变化,他们的研究结果与上述油料作物研究结果基本一致。但以上研究均未与科学的模型相结合。
在我国,研究农业生产系统模拟器(APSIM)的学者,科研方向主要集中在:李广等[11-12]基于APSIM模型旱地春小麦的降水量、CO2、温度对产量的影响;王亚许等[13]利用APSIM模型对春玉米生育期旱灾损失敏感性进行了定量分析研究;沈禹颖[14]、古丽娜扎尔·艾力等[15]基于APSIM模型对黄土旱塬区苜蓿-小麦轮作系统进行了研究。然而,目前国内外对于胡麻作物产量与模型相结合的研究报道极少,有待继续深入。因此,本文以胡麻为研究对象,采用APSIM 模型,建立胡麻干旱胁迫模型,利用大田试验数据检验模型;揭示胡麻生理生化代谢指标对干旱胁迫的响应,为应用模型分析胡麻干旱胁迫提供理论依据。
1 材料与方法
1.1 研究区概况
试验长期在黄土高原的甘肃省定西市西巩驿镇(104°37′12″E,35°34′48″N)进行,试验地区为旱地,海拔高度1793 m,年日照时数2500 h,年均降水量小于400 mm,无霜期146 d,年均气温7 ℃,年均蒸发量1524.8 mm。试验土壤从大田间采取,为黑垆土。
1.2 试验设计
供试材料选用陇亚10 号(LY10)、陇亚11 号(LY11)、陇亚杂1 号(LYZ1),将胡麻种子播种于直径23 cm,深30 cm的塑料盆中。盆内装有大田供试土壤(黑垆土)与细土按1∶2 比例混合,并提前对基质进行灭虫,配260 g磷肥,混合搅拌均匀。试验设置4个干旱时期,分别为苗期、枞形期、现蕾期、终花期。试验处理分为3 组:试验土壤相对含水量设置为田间最大持水量的70%~75%,即为对照组(CK);田间最大持水量的50%~55%,即为中度胁迫组(MD);田间最大持水量的35%~40%,即为重度胁迫(SD)。采用称重法进行控制水分。为防止系统误差,随机区组排列,并在每盆上用标签纸进行标记。栽培管理同大田生产。
1.3 数据测定
1.3.1 气体交换测定气孔导度、蒸腾速率均采用美国公司Li-cor 便携式光合作用测定系统测定(型号为Li-6400型)[16-17]。
1.3.2 渗透调节、抗氧化代谢物质测定脯氨酸(Pro)含量测定采用“茚三酮法”[17],丙二醛含量测定采用的“硫代巴比妥酸法”[18],可溶性蛋白含量测定采用“考马斯亮蓝G-250 染色法”[17],CAT 活性测定采用紫外分光光度法[18],SOD 活性测定参照氮蓝四唑法[18],POD活性测定参照愈创木酚法[18]。
1.4 模型构建
在建模过程中,本模型以胡麻生长特点为依据,综合考虑辐射利用率,辐射截获对胡麻光合速率的影响,同时还添加了CO2因子、光合作用水分胁迫因子、蒸腾效率系数。参数中的相关系数均跟生物学相关,在模型中都具有一定的意义,在描述胡麻生理生化指标响应干旱胁迫的反应机理方面也能够更准确,使模型更具有普适性。
农作物模拟模型(APSIM),即农业生产系统模拟器,是一种农业系统建模框架,由澳大利亚科学家研制[19]。通过内含的各个模块(土壤、作物、肥料、水、牧场等)模拟农业生产系统中各环节[20]。作物在干旱地区会受到土壤、降水、光照、季节等条件不同程度的影响,其中光照强度、昼夜温差和季节变化都会对实际的光合作用产生关键性作用[21]。在20 世纪80 年代,SUCROS[22]、MACROS[23]和ORYZA[24-25]模型的出现,因它们较强的机理性,使得光合生产模型研究得到飞速发展。随后出现的CERES[26-27]系列模型和GOSSYM[28]模型因搭建结构和参数的复杂繁多,忽略了实际的生产管理。现有相关的油料作物模型,如油菜作物模型通过模拟光合作用及光辐射,较为成熟的有APSIM-Canola[29-30]、CERES-Rape[31-32]和LINTUL-BRASNAP[33-34]等,其中生物量的累计通过辐射利用率计算生物量来进行[16]。
本模型所需要的试验区2013—2014 年气象数据由甘肃省气象局气象中心提供。胡麻生理生化对干旱胁迫的响应中,光合作用进程对干旱尤为敏感。光合作用、生物量累计干旱胁迫因子计算如下:

式中:fW,photo为水分胁迫反应函数;Wu为实际水分吸收量(mm);Wd为土壤水分需求量(mm);Ws为潜在土壤水分供应量(mm);ΔQr为由辐射截获决定的潜在生物累积量(kg·hm-2);R为呼吸率;TE 为蒸腾效率,与平均日蒸汽压差VPD(kPa)和CO2因子的倍数有关;KL 为根吸水值(mm);SW 为土壤含水量(mm);LL为凋萎系数(mm·mm-1);fc,TE为用于蒸腾效率的CO2因子,是CO2浓度函数,当CO2浓度从350增加到700 时(μL·L-1),fc,TE从1 线性增加到1.37;fTE为蒸腾效率系数;VPD为蒸汽压差;I为辐射截获量(MJ·m-2);RUE为辐射利用率(g·MJ-1·m-2);fd、fs、fc分别为散射因子、胁迫因子和CO2因子;fv为春化因子;Tmax、Tmin分别为日最高气温、日最低气温(℃)。

式中:FW为水分胁迫光合作用;Fp为潜在光合作用;Et和Et,p分别为实际蒸腾和潜在蒸腾作用(mm);Ei和Ei,p分别为截获水的实际和潜在蒸发量(mm)。
水分供给(水分需求)和水分胁迫因子如图1所示。一般来说,在现有的模型中,都会计算一个应力因子即水分胁迫,在0 和1 之间变化(0 代表受到完全胁迫,1 代表没有受到胁迫),而干旱胁迫可能会降低植物的发育速率[35-36]。

图1 APSIM模型中水分供给(水分需求)和水分胁迫因子的关系Fig.1 Relationship between water supply(water demand)and water stress factors in APSIM model
1.5 模型参数校正
APSIM 模型对作物种植的影响因素、生理生态机理等方面具有比较好的模拟能力,应用广,精度高。但由于APSIM 模型本身以南半球的各地理环境因素为主要研究依据,与北半球的研究区存在着显著差异,模型在进行模拟前需要进行本地化参数输入,用科学的方法对模型模拟参数进行不断校准、调整、优化[37]。本研究通过运行本地化APSIM模型参数模拟胡麻光合作用和干旱胁迫,模型所需要的水量来源于本课题组研究成果《胡麻目标产量指标化调控技术规程》,并结合2013—2014年实测大田试验数据验证构建模型。胡麻模型参数分为2种,一种是胡麻物种参数,数据来源于前人研究[38-39],另外一种是可以调整的遗传参数(表1),通过编程运行BPPPAR 投影寻踪自回归模型[40],并使用RAGA 优化投影指标函数的方法[40]调整表1 参数,使得模型的模拟数据尽可能的接近实测数据。

表1 胡麻生理生化代谢指标对干旱胁迫响应的模型参数Tab.1 Model parameters of response of flax physiological and biochemical metabolic indices to drought stress
1.6 模型评价方法
模型本地化的关键在于使用试验站定西地区2013—2014年的气象数据,供试土壤黑垆土的土壤资料,以及在该环境中种植本地供试品种的作物管理数据来运行模型,通过调整模型参数使得模型模拟数据与实际数据之间的差距逐渐变小,从而实现模型的校准与调整;然后使用另外一年的数据检验模型模拟数据与实测数据的差异。差异越小,模型精确度越高。
本研究针对模型的可行性检验,是通过与研究区大田数据的实际产量值进行对比。检验方法采用2 种指标进行检验,分别为利用均方根误差(RMSE)和决定系数(R2)[41]。一般认为,模拟结果与实际测量值之间的差距,即模拟精度。其中RMSE数值越小,精度越高;R2的值越趋近于1,模拟效果就越理想。
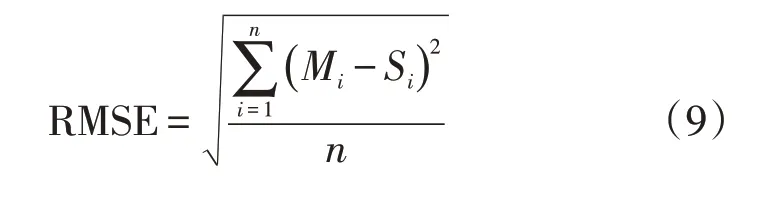

式中:Mi为实测值;Si为模拟值;n为观察值数目;Mˉ为Mi的平均值;-S为Si的平均值。
1.7 数据统计分析
本文所有数据均采用软件Excel for Mac 和SPSS 26.0 进行分析处理。建模语言使用C#编程语言。
2 结果与分析
2.1 模型检验
利用定西市西巩驿镇研究区2013—2014 年试验中的实际产量,验证模型模拟胡麻生理生指标响应干旱胁迫时,其中光合指标对产量影响的有效性。结果表明(图2),产量模拟值的RMSE 为41.3159~155.6685 kg·hm-2,平均值为80.1837 kg·hm-2。产量的实测值与模拟值R2为0.8929~0.9894,平均值为0.9387。可见,模型能够较好地模拟胡麻生理生化指标响应干旱胁迫的胡麻产量。

图2 胡麻产量的实测值与模拟值Fig.2 Measured and simulated values of flax yield
2.2 胡麻生理生化指标对干旱胁迫的响应
2.2.1 干旱胁迫下气体交换的变化蒸腾速率(Tr)是用来调整各类的作物机制,还能应对环境变化[42]。由图3a 可知,胡麻受到干旱胁迫的程度越大,Tr 受到的抑制尤为明显,3 个胡麻品种的Tr 也均为下降趋势。其中,LYZ1 和LY10 面对干旱胁迫表现更为敏感。在中度胁迫处理下,LY11 的Tr 平均降幅最小,较对照组降低25.00%;重度胁迫处理下,LY10的Tr 平均降幅最大,较对照组下降86.67%。因此,干旱胁迫的加深给胡麻Tr带来了不利影响。
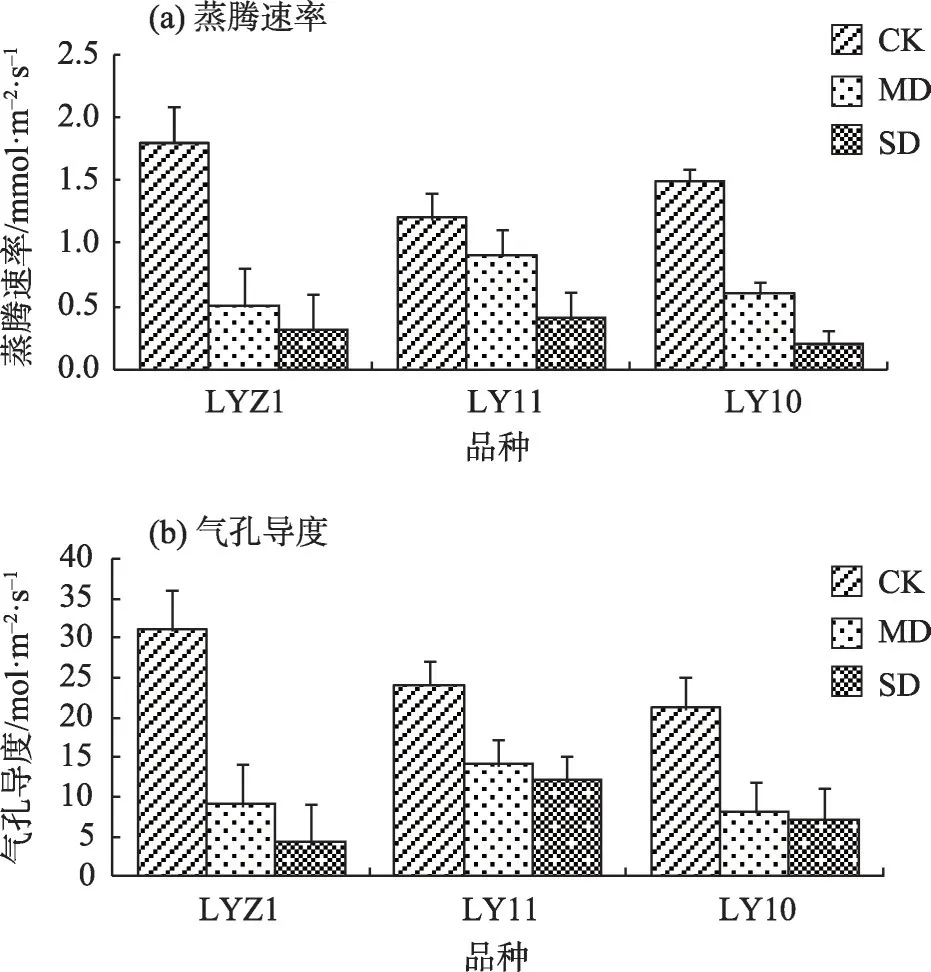
图3 干旱胁迫对气体交换的影响Fig.3 Effects of drought stress on gas exchange
气孔导度(Gs)可以反映作物遭受外部不利影响受损的程度[43]。由图3b 可知,干旱胁迫程度越大,3 个胡麻品种的Gs 差异越显著,都呈现下降趋势。在中度和重度胁迫处理下,LYZ1 的Gs 较对照组平均降幅最为明显,分别为70.97%和87.10%。由此表明,干旱胁迫的加重会使胡麻叶片的Gs 降低,蒸腾失水也同时下降。
2.2.2 渗透调节代谢指标对干旱胁迫的响应在不同干旱胁迫下渗透调节物质(脯氨酸、可溶性蛋白和丙二醛)会从叶片内产出,而大量脯氨酸的积累有助于保护植物细胞结构和功能,维持植株渗透势(图4a)。在胡麻的不同生育时期干旱胁迫均使胡麻叶片脯氨酸含量有较大幅度增加,在重度胁迫处理下,LYZ1、LY11 和LY10 现蕾期叶片脯氨酸含量最高且增长幅度较大,分别为116.85%、236.22%、81.22%。由此可证明,胡麻不同品种脯氨酸含量的增加会使其表现出较强的干旱适应能力。
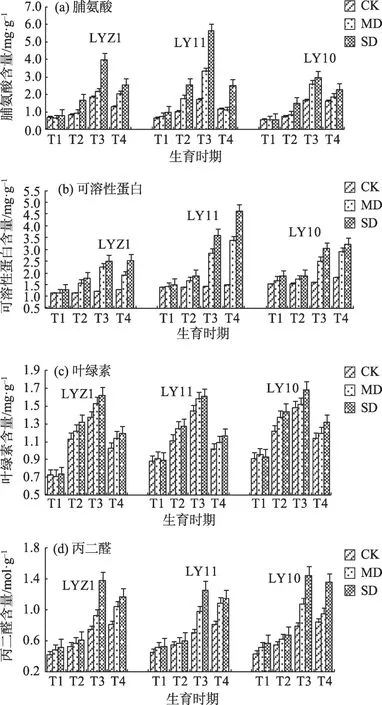
图4 干旱胁迫下渗透可调节物质含量变化特征Fig.4 Variation characteristics of osmotic adjustable substance content under drought stress
3个胡麻品种的可溶性蛋白含量在干旱胁迫下的不同生育时期有明显增加,且增加幅度由大到小为:现蕾期>终花期>枞形期>苗期(图4b)。在重度胁迫处理下,LYZ1、LY11 和LY10 现蕾期可溶性蛋白增幅最大,分别为107.17%、154.33%、91.12%;而苗期可溶性蛋白含量较对照组增幅最小,分别为12.78%、9.31%、22.90%。
胡麻响应干旱胁迫时,不同生育期叶绿素的含量增加明显。在重度胁迫处理下,LYZ1、LY11 和LY10 苗期叶绿素含量较对照组升高,增幅分别为1.39%、1.94%、1.75%,但差异不明显;而较对照组显著增加的3 个时期是现蕾期、枞形期和终花期;其中,在重度胁迫处理下叶绿素含量变化最高值出现在现蕾期,LYZ1 叶绿素含量较对照组升高了18.11%(图4c)。植株叶片由于受到干旱胁迫的影响,叶片叶绿素含量增加,叶片相对含水量降低,导致叶绿素浓度的增加。
植物响应干旱胁迫时发生膜质过氧化,其器官的主要产物为丙二醛。在不同干旱胁迫下,使胡麻叶片在不同生育时期的丙二醛含量增加(图4d)。在重度胁迫处理下,LYZ1、LY11 和LY10 现蕾期叶片丙二醛含量都呈大幅度增长,分别为84.00%、80.00%、83.54%。干旱胁迫程度较低时,3个胡麻品种体内的丙二醛含量也较低,但超过植物的生理极限,会导致丙二醛的大量产生。
2.2.3 抗氧化代谢指标对干旱胁迫的响应CAT可以将H2O2转变为活性较低的物质,从而使膜质不发生过氧化作用而起到保护作用。在不同干旱胁迫下的胡麻生育时期,叶片CAT 活性有所下降(图5a)。其中,在重度胁迫处理下,LYZ1、LY11和LY10苗期胡麻叶片CAT活性比对照组降幅最少,分别为13.28%、12.43%、15.38%;现蕾期比对照组降幅最大,分别为28.09%、20.44%、27.00%,低于对照组明显。植株CAT 活性的降幅越小,抗氧化能力就越强。
SOD通过催化反应并与POD协同组织脂质过氧化物的积累,主要作用是维持植物体内代谢平衡[44]。在重度胁迫处理下,LYZ1、LY11和LY10枞形期、现蕾期增幅较大分别为14.08%、21.33%、11.76%和21.33%、24.61%、19.46%;苗期和终花期在同样处理下CAT活性也显著增加(图5b)。
POD 是保护性酶,对环境变化最为敏感[45]。不同生育时期在干旱影响下均显著增加了3个胡麻品种叶片的POD 活性。在重度胁迫处理下,LYZ1、LY11 和LY10 终花期叶片POD 活性较对照增幅最大,分别为28.00%、26.90%、26.09%;现蕾期次之;苗期POD 活性增幅最小,分别为12.50%、27.91%、13.42%(图5c)。

图5 干旱胁迫下抗氧化代谢物质含量变化特征Fig.5 Variation characteristics of antioxidant metabolite content under drought stress
2.2.4 干旱胁迫对胡麻生理指标的相关性分析作物的各类生理指标受到干旱胁迫时,会产生一个复杂的生理生化过程,它们在此反应过程中,呈现不同的相关性、相互作用和影响(表2)。
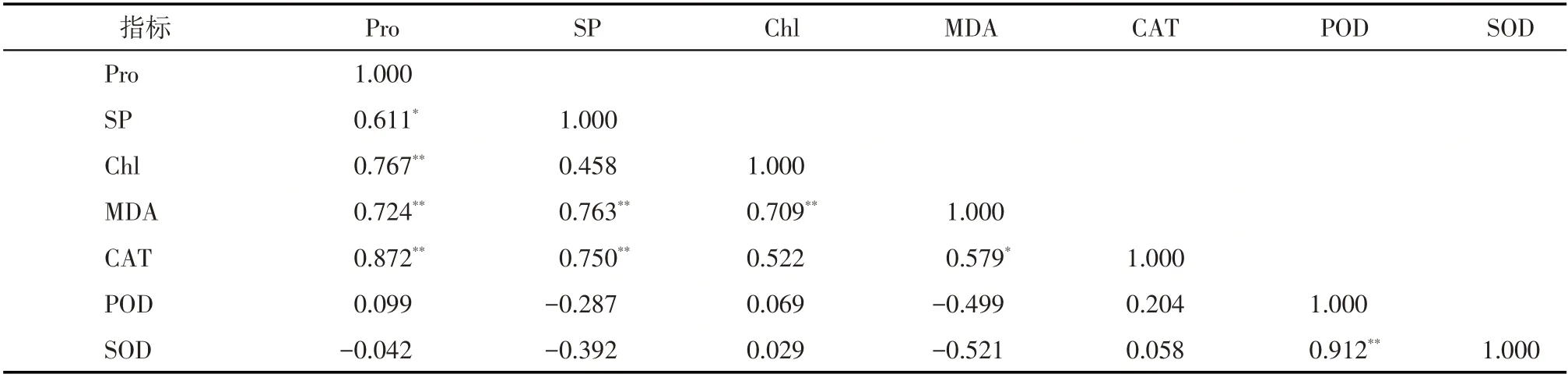
表2 干旱胁迫下胡麻生理生化代谢指标的相关性分析Tab.2 Correlation analysis of physiological and biochemical metabolism indices of flax under drought stress
3 讨论
李玥等[46]以胡麻为研究对象,通过不同肥料、不同播种方式以及不同种植密度等初步检验,构建其光合生产与干物质积累模拟模型有初步成效。王钧等[47]利用APSIM 模型对旱地春小麦的不同生育期、不同干旱程度胁迫对小麦生育期、籽粒数、千粒重及产量进行了模拟,适宜性较好。汤亮等[30]构建油菜地上部器官干物质分配动态模拟模型,结合品种遗传特性和播期试验资料,检验结果发现具有较好预测性。本研究在APSIM模型的基础上,模型模拟产量值的R2为0.8929~0.9894,表明模拟胡麻生理生化在不同生育时期、不同程度干旱胁迫条件下的产量有较高精度,且具有一定的适宜性。
在本研究中,3个胡麻品种在干旱胁迫下株高、茎粗和根系生长受到抑制,品种不同在不同的胁迫时期受到抑制程度也不同。叶绿素的多少直接反应作物光合能力的强弱[48]。一般研究认为,干旱所造成的植株叶片失水,相对含水量降低会抑制叶绿素的合成,还会造成其分解加速,最终使植物叶片内的叶绿素含量降低[49]。同时,部分研究表示植物在响应干旱胁迫时,叶绿素含量会增加显著[50-51]。本研究结果表明,胡麻在响应干旱胁迫时,不同生育时期胡麻叶片叶绿素含量上升,其中在重度胁迫处理下,LYZ1、LY11 和LY10 都在现蕾期上升幅度最大。这可能是因为胡麻叶片在受到干旱胁迫时,叶片相对含水量降低,使其中的叶绿素无法分解或分解的少部分产生浓缩,导致叶绿素含量逐步上升[52]。
渗透调节物质(脯氨酸、丙二醛、可溶性蛋白)是调节生理机制和响应干旱最直接、最重要的方式之一[53]。本研究发现,在重度胁迫处理下,这3种物质的含量均显著上升,LYZ1、LY11 和LY10 现蕾期和终花期脯氨酸和可溶性糖增幅最大,反映了胡麻通过以上指标在不断适应不同干旱情况。由于这些指标对干旱胁迫响应机制有所不同,研究还发现脯氨酸含量增幅高于可溶性糖含量,且水平显著。胡麻在响应干旱胁迫时,丙二醛含量增加;其中,苗期和枞形期积累相对较少,现蕾期和终花期含量增加明显,说明受到干旱胁迫时,胡麻苗期有一定程度的耐受性。
抗氧化代谢指标(SOP、POD、CAT)作为活性氧的清除者,可衡量作物的抗逆性的强弱[54]。本研究发现,将POD 活性维持在较高水平,胡麻能更好适应胁迫和带来的危害,在重度胁迫处理下,LYZ1、LY11和LY10终花期POD活性增幅最大,SOD、POD活性提高显著,CAT 活性下降,说明胡麻抵御干旱时,可以调动抗氧化保护酶,提高抵御能力,这也是作物在遇到干旱胁迫时所做出的一种生理反应。
4 结论
本研究以陇亚10号(LY10)、陇亚11号(LY11)、陇亚杂1号(LYZ1)为主要试验材料,基于APSIM模型建立胡麻干旱胁迫模型,研究了胡麻不同生育时期生理生化代谢指标对干旱胁迫的响应。主要结论如下:
(1)基于APSIM 模型,利用2013—2014 年甘肃省定西市西巩驿镇试验数据,建立胡麻干旱胁迫模型,模型检验结果表明模型能够模拟不同胁迫下胡麻生理生化指标的变化,具有较好的拟合度,可靠性较高。
(2)重度干旱胁迫下,生理生化指标脯氨酸、可溶性蛋白、丙二醛受影响最大。渗透调节代谢指标均呈增长趋势,且在现蕾期增幅最大;抗氧化代谢指标CAT 活性呈现下降,且现蕾期降幅最大,SOD和POD活性有所增加,且终花期增幅最大。
(3)3个胡麻品种LYZ1、LY11和LY10中,生理生化指标受干旱胁迫影响最大的是LY11,受影响最小的是LY10。
猜你喜欢
——基于CMS 模型的实证分析
市场周刊(2022年9期)2022-09-29
昆明医科大学学报(2021年10期)2021-12-02
农业科技与信息(2021年19期)2021-11-30
农业科技与信息(2020年20期)2020-12-18
音乐研究(2019年5期)2019-11-22
江苏农业学报(2019年1期)2019-09-10
意林(2018年20期)2018-10-31
食品与生活(2018年1期)2018-02-07
Coco薇(2017年5期)2017-06-05
环球时报(2015-05-13)2015-05-13
