
六个葡萄砧木品种(系)的抗寒性评价
2022-05-29 08:08郭艳兰牟德生赵连鑫王鑫马宗桓张兆铭张利年
中外葡萄与葡萄酒 2022年3期
郭艳兰,牟德生,赵连鑫,王鑫,马宗桓,张兆铭,张利年
(1. 武威市林业科学研究院,甘肃武威 733000;2. 甘肃农业大学园艺学院,甘肃兰州 730070)
武威市地处河西走廊东端,其独特的地理位置和气候条件,为酿制优质葡萄酒提供了绝佳的先天环境,使得武威成为全国酿酒葡萄九大产区之一。但武威产区冬季寒冷干燥,低温冻害发生频繁,葡萄需要下架埋土才能越冬,增加生产成本的同时也限制了该区域酿酒葡萄产业的健康可持续发展。因此,探索提高葡萄抗寒能力的行之有效的措施成为当前武威地区酿酒葡萄产业发展急需解决的关键问题。
葡萄砧木的抗寒性是其实现利用价值的关键之一,采用抗寒砧木进行嫁接不仅能提高葡萄的抗寒能力,还可以提高树体营养、改善果实品质,对产业发展具有重要的促进作用[1-3]。目前,有关酿酒葡萄砧木的抗寒性已开展了较多的研究,如牛铁荃等[4]以不同葡萄砧木的实生苗叶片为研究对象,测定了相对电导率及相关生理生化指标,并利用灰色关联分析、隶属函数、聚类分析等方法对其耐寒性开展了评价;李妍琪等[5]对葡萄砧木的4个杂交组合的亲本及后代开展了抗寒性鉴定;李鹏程等[6]以8种葡萄砧木为试材,研究了低温胁迫下的失水速率、萌芽率等,并比较了LT50与隶属函数法综合评价结果的一致性;钟海霞等[7]对7个葡萄砧木根系的抗寒性进行了研究。而武威产区有关葡萄砧木抗寒性的综合分析及评价较少。本文以从法国和意大利引入的6个酿酒葡萄砧木品种(系)为材料,在人工模拟低温条件下,测定其相对电导率、丙二醛(MDA)等相关生理生化指标,并采用隶属函数法对其抗寒性进行综合分析及评价,以期筛选出抗寒性强的砧木品种(系),为该地区葡萄抗寒砧木的选择及利用提供理论依据。
1 材料与方法
1.1 供试材料
供试砧木品种(系)为SO4-762、SO4-102、CFS57-34、P40-151、101-141、1103P-113,均采自武威市民勤县甘肃夏博岚葡萄种植基地,2015年从法国和意大利引入。树体5年生,栽植密度0.5 m×3 m,厂字形架式, 田间管理水平一致,植株生长良好。2020年10月冬剪时采集粗度1 cm、生长健壮的一年生成熟枝条,带回武威市林业科学研究院恒温库沙藏备用。
1.2 试验处理
将沙藏枝条取出冲洗干净,剪成10 cm左右小段,分别装入自封袋中并做好标记,置于高低温试验箱(型号:LRHS-101A-LJ)中进行低温处理,先以4 ℃·h-1的速率缓慢降至目的温度,保持24 h后再以相同速率升至4 ℃,取出置于4 ℃冰箱保存待测。共设置-14、-18、-22、-26、-30 ℃ 5个低温处理,以4 ℃为对照。
1.3 测定指标及方法
相对电导率:用修枝剪将低温处理过的枝条避开芽眼剪成约1 mm厚的薄片,混合均匀后称取1 g置于25 mL试管中,加入20 mL蒸馏水,加盖于室温静置4 h,用DDSJ-308A型电导率仪测定初始电导率,测前摇匀;于沸水中煮30 min,冷却至室温后测定终电导率,计算相对电导率(相对电导率=初始电导率/终电导率×100%)和低温半致死温度(LT50)[8],并进行曲线拟合,拟合方程如下:

其中,Y代表相对电导率,a、b为方程参数,K为常数,代表最大电解质渗出率,由于Y是累积频率,此处K取100,LT50即方程拐点,计算方式为:
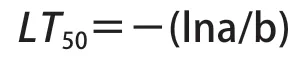
生理生化指标:将各处理枝条剪成2~3 mm薄片,用液氮冷冻后迅速将枝条粉碎,置于-80℃冰箱保存用于测定生理生化指标。MDA、脯氨酸、可溶性糖、可溶性蛋白含量及过氧化氢酶(CAT)活性均采用武汉纯度生物酶联免疫分析试剂盒(ELISA)进行测定。
1.4 数据处理与分析
用Microsoft Office进行数据整理和制图、SPSS 25.0进行数据分析。
抗寒性的综合分析与评价采用隶属函数法[9-10]进行,计算公式如下:
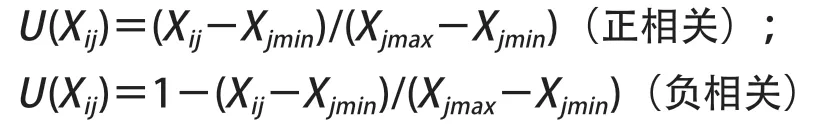
式中,U(Xij)为测定i品种j指标的抗寒隶属函数值,Xij为i品种j指标的测定值,Xjmin为j指标的最小值,Xjmax为j指标的最大值,i表示品种,j表示抗寒指标。
2 结果与分析
2.1 低温处理对葡萄砧木枝条相对电导率的影响
低温处理后各砧木品种(系)的相对电导率变化如图1所示,随着温度的降低,各砧木枝条的相对电导率均呈上升趋势,且升高趋势各有不同。在-18~4 ℃时,各品种(系)相对电导率随处理温度的降低表现缓慢上升,其中,101-141上升幅度最小,1103P-113次之,P40-151上升幅度最大;随着温度的继续降低,各品种(系)的相对电导率上升幅度增加,-22 ℃时变化最为明显;-30~-26 ℃温度处理时,各品种(系)相对电导率随温度降低继续升高但幅度变缓;在低温处理过程中,P40-151的相对电导率随温度降低的增幅最大,CFS57-34的相对电导率随温度降低的增幅最小,且-30 ℃温度处理后,101-141的相对电导率最大,P40-151次之,而CFS57-34的相对电导率最小。

图1 低温处理下不同砧木品种枝条的相对电导率Figure 1 Relative conductivity in branches of different rootstock under low temperature treatments
2.2 Logistic模型建立及低温半致死温度
根据试验温度范围内各砧木的相对电导率建立Logistic方程,并根据方程计算半致死温度LT50,如表1所示。各砧木Logistic方程拟合度R2在0.7272~0.8961,表明所拟合方程具有较高的可靠性,由此计算的LT50真实可用。各砧木的LT50在-25.86~-21.85℃,LT50值越小,说明抗寒性越强。
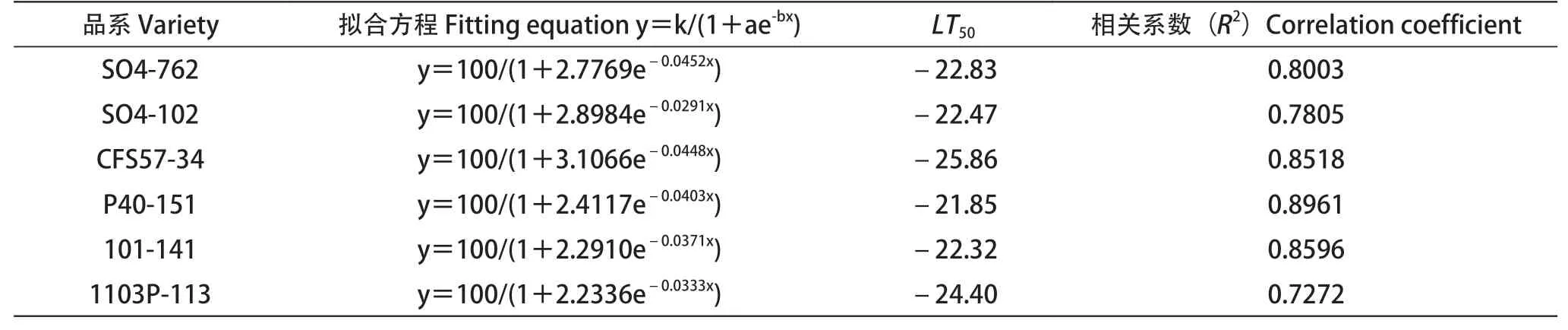
表1 低温处理下不同砧木品种枝条的低温半致死温度和拟合方程Table 1 LT50 and fitting equation in branches of different rootstock varieties under low temperature treatments
2.3 低温处理对葡萄砧木枝条生理指标的影响
2.3.1 低温处理对MDA的影响
低温处理后各砧木品种(系)枝条的MDA含量变化如图 2所示,随着温度的降低,各砧木枝条的MDA含量均呈上升趋势。-30~4 ℃低温处理过程中,P40-151枝条MDA含量升高幅度最快,而CFS57-34枝条MDA含量升高幅度最慢,其余4个品种(系)枝条的MDA含量变化介于二者之间;-30 ℃低温处理后,P40-151枝条的MDA含量显著高于其他品种(系),而CFS57-34枝条的MDA含量显著低于其他品种(系)。

图2 低温处理下不同砧木品种枝条的MDA含量Figure 2 MDA content in branches of different rootstock under low temperature treatments
2.3.2 低温处理对游离脯氨酸影响
如图3所示,随着温度的降低,各砧木品种(系)枝条的游离脯氨酸含量均呈先上升后下降的趋势,但变化幅度各不相同。4 ℃时1103P-113枝条的游离脯氨酸含量最高,而SO4-762枝条的游离脯氨酸含量最低;当处理温度降至-22 ℃,101-141、CFS57-34枝条的游离脯氨酸急剧升高且达到最高,而其余各品种(系)枝条的游离脯氨酸随着温度的继续降低持续增加,至-26 ℃时,SO4-762、SO4-102、P40-151、1103P-113枝条的游离脯氨酸含量达到最高,后随温度的继续降低,各砧木的游离脯氨酸含量持续降低。低温处理过程中,101-141、CFS57-34枝条游离脯氨酸含量随温度降低的增幅较大,SO4-102、P40-151枝条游离脯氨酸含量随温度降低的增幅较小。

图3 低温处理下不同砧木品种枝条的游离脯氨酸Figure 3 Freee proline content in branches of different rootstocks under low temperature treatments
2.3.3 低温处理对可溶性糖的影响
由图4可知,不同砧木枝条的可溶性糖含量随着处理温度的降低持续增加。4 ℃时CFS57-34枝条的可溶性糖含量最低,其次为1103P-113,而P40-151枝条的可溶性糖含量最高。随着处理温度的不断降低,各砧木枝条可溶性糖含量快速升高,至-30 ℃时达到最高水平,且CFS57-34枝条的可溶性糖含量最高,且显著高于其他砧木品种(系),101-141枝条的可溶性糖含量最低,但与其余4个品种(系)无明显差异。
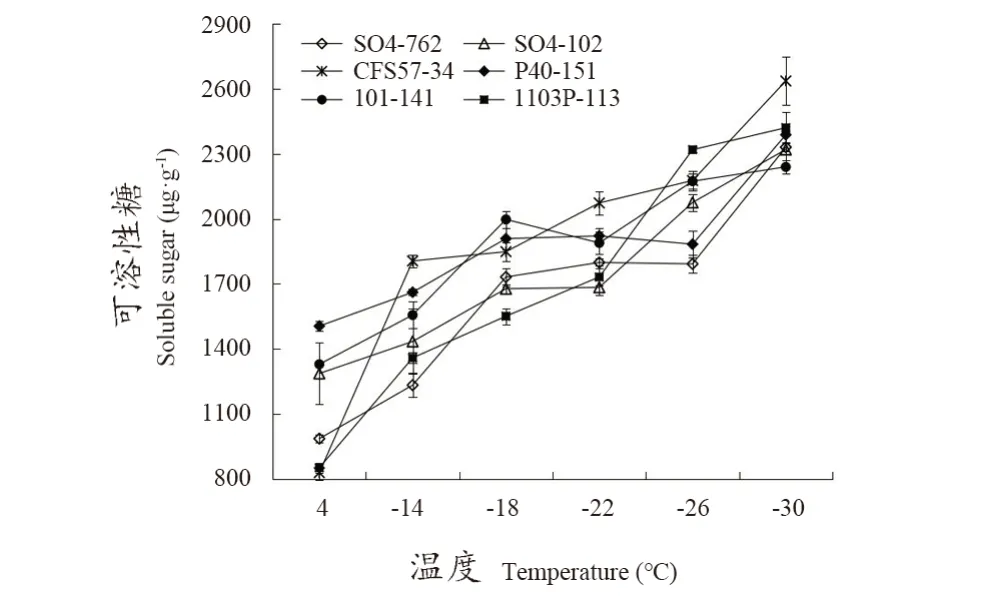
图4 低温处理下不同砧木品种枝条的可溶性糖含量Figure 4 Soluble sugar content in branches of different rootstocks under low temperature treatments
2.3.4 低温处理对可溶性蛋白的影响
由图5所示,各品种(系)枝条的可溶性蛋白含量随温度降低逐渐升高。4 ℃时,CFS57-34枝条的可溶性蛋白含量最高,其次为SO4-762,而SO4-102的可溶性蛋白含量最低,且显著低于其余各品种(系)。随着温度的降低,各品种(系)枝条的可溶性蛋白含量出现不同程度的增加,至-30 ℃时增至最高,其中,1103P-113的可溶性蛋白含量最高,其次为CFS57-34,二者无明显差异,P40-151的可溶性蛋白含量最低。
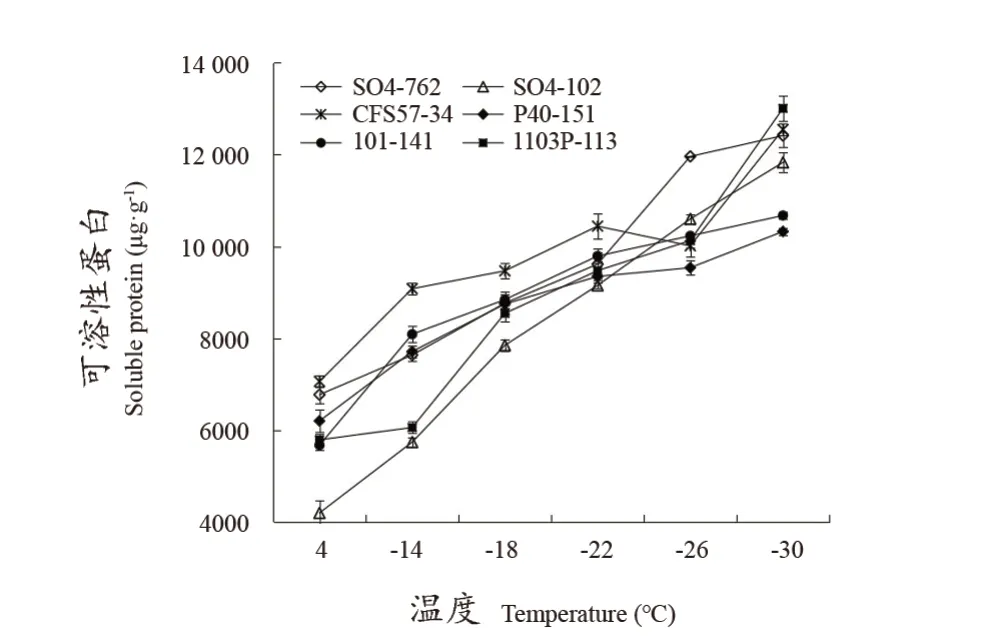
图5 低温处理下不同砧木品种枝条的可溶性蛋白Figure 5 Soluble protein content in branches of different rootstocks under low temperature treatments
2.3.5 低温处理对CAT活性的影响
由图6可以看出,不同砧木枝条的CAT活性随温度的变化不同。SO4-102和P40-151枝条的CAT活性随温度降低先升高后下降。在-22 ℃时升至最高水平,而后随着温度的继续降低出现下降;其余4个品种枝条的CAT活性随温度降低持续上升,至-30 ℃时达到最高,且1103P-113、CFS57-34的CAT活性显著高于其他品种,而P40-151的CAT活性最低。

图6 低温处理下不同砧木品种枝条的CAT活性Figure 6 CAT activity in branches of different rootstocks under low temperature treatments
2.4 不同砧木品种抗寒性分析
采用隶属函数法对6个葡萄砧木品种(系)枝条的相对电导率及各抗寒相关生理指标进行综合分析及评价,结果见表2。根据抗寒指标的平均隶属函数值大小可知,各砧木品种的(系)抗寒性由强到弱依次为:CFS57-34>1103P-113>101-141>SO4-762>SO4-102>P40-151。

表2 不同砧木品种各抗寒指标的隶属函数值Table 2 Subordination value of cold resistance indexs of different rootstock varieties
3 讨论与结论
低温会破坏植物细胞的结构与功能,使细胞质膜发生改变,细胞内离子、糖类等物质会出现外渗,从而导致电解质的升高[11]。细胞电解质的外渗程度可用相对电导率来表示,且相对电导率越高,表明细胞内的电解质外渗越多,细胞膜的透性越大,也说明植株细胞膜受到伤害的程度越大,抗寒能力越弱[12]。目前,利用相对电导率法对果树抗寒性进行研究已得到了广泛应用[13-17]。本文测定结果显示,各砧木枝条的相对电导率均随温度的降低而升高,且相对电导率随温度降低的增幅越多,抗寒性越弱,反之抗寒性越强。这与前人研究结果一致[18-20]。根据Logistic方程预测得出6个葡萄砧木品种(系)枝条的半致死温度在-25.86~-21.85℃,部分品种的低温半致死温度与刘钰玺等[18]的研究结果有一定差异,这主要与枝条的低温处理方式、相对电导率的测定方法等因素有关。
植物的生理生化指标与其抗寒性有着密切联系[21]。植物在受到低温胁迫时,可以通过调节体内的生理生化过程来诱导自身细胞代谢向着适应低温的方向发展,从而提高抗寒能力[19]。MDA是膜脂过氧化反应的最终产物,是植物细胞膜系统受害的重要标志之一,其含量的增加与植物的受害程度有极显著的相关性[22-23]。研究结果表明,随着温度的降低,各品种(系)砧木枝条的MDA含量均逐渐升高,其中,P40-151枝条MDA含量增幅最快且最终维持在较高水平,而CFS57-34枝条MDA含量增幅最慢且维持在较低水平,这与相对电导率变化一致,也进一步证实了MDA变化与其抗寒性的关系[24-25]。
游离脯氨酸、可溶性糖、可溶性蛋白是植物体内重要的渗透调节物质,与植物的抗寒性密切相关。游离脯氨酸的积累可以提高植物体内渗透压,增强保水力,提高植物的抗寒性[26];可溶性糖含量的增加可使植物细胞液浓度升高,降低冰点温度,可溶性蛋白具有较高的亲水性,其含量与可溶性糖含量密切相关,因此常被作为植物抗寒性鉴定的重要指标[27-28]。本研究中,各品种(系)砧木枝条的游离脯氨酸含量随温度的降低先上升后下降,可溶性糖含量和可溶性蛋白含量随温度的降低而升高,且抗寒性较强的品种其脯氨酸、可溶性糖和可溶性蛋白含量随温度的降低升高较快,并且最终保持在较高水平,而抗寒性较弱的品种则正好相反,脯氨酸、可溶性糖、可溶性蛋白含量的升高较慢且水平较低。这与施明[12]、钟海霞[29]、付晓伟[30]等的研究结果基本一致,说明在遭受低温胁迫时,葡萄枝条可以通过代谢增加体内的渗透调节物质,以此来保护机体免受低温伤害。
CAT是植物体内的活性氧化保护酶之一,低温环境下,它可以通过自身的调控来保护植株,以减轻植物的低温损伤。在本研究中,SO4-102和P40-151枝条的CAT活性随温度降低先升高后下降,而其余品种枝条的CAT活性随温度降低持续上升,表明在一定的低温范围内,葡萄枝条可以通过提高氧化防御酶活性来增强自身的耐寒能力,但当低温超出了该品种的耐受范围时,细胞膜受到损害,酶活降低,这主要与砧木品种(系)的抗寒能力强弱有关[31-32]。
与植物抗寒性相关的指标有很多,单一的某个指标具有片面性,在植物抗寒性研究中,通常需要对多个抗寒指标进行综合分析,以此来判断其抗寒性[33,31,29]。本文采用隶属函数法对6个砧木品种(系)的抗寒性进行了综合分析,得出各品种(系)的抗寒性强弱顺序为CFS57-34>1103P-113>101-141>SO4-762>SO4-102>P40-151,这与预测得出的各砧木品种(系)的LT50排序略有差异,可能与LT50为单一指标有关。
综合以上表明,法国和意大利引入武威地区的6个酿酒葡萄砧木品种(系)中,CFS57-34的抗寒性最强,1103P-113、101-141、SO4-762、SO4-102的抗寒性较强,P40-151的抗寒性最弱。
猜你喜欢
热带作物学报(2022年5期)2022-06-01
安徽化工(2021年3期)2021-05-29
现代农业科技(2020年10期)2020-06-04
科学与财富(2019年15期)2019-10-21
山西果树(2017年4期)2018-02-08
物联网技术(2017年5期)2017-06-03
安徽农学通报(2016年24期)2017-01-12
科技资讯(2015年7期)2015-07-02
湖北农业科学(2014年13期)2014-08-28
山东农业科学(2014年3期)2014-08-20
