
我国草莓胶孢炭疽菌的多基因联合鉴定与致病性分析
2022-05-26 02:53侯圣凡华战迎刘峻杰董振飞冯泽坤闫佳琪王红清
中国农业大学学报 2022年4期
侯圣凡 华战迎 刘峻杰 董振飞 冯泽坤 闫佳琪 王红清
(中国农业大学 园艺学院,北京 100193)
草莓(Fragaria×ananassaDuch)是一种重要的经济作物,因其种植周期短、结果早且经济效益高,所以在地方特色经济中占有重要的地位[1]。我国草莓主产区有山东、安徽、辽宁、河北和江苏等地。近年来,山西和云南等地的草莓产业也得到迅速发展。我国草莓主要是依靠设施栽培,由于多年连续种植,导致草莓根部病害近年来频繁发生,其中以真菌性病害对草莓的影响最为显著,对我国草莓产业的健康发展造成了巨大的危害[2]。
炭疽病是一种寄主广泛且种类繁多的常见植物病害,而且此类真菌存在的地域范围广,可侵染多种农作物。1931年,Brooks等[3]最先报道了侵染草莓匍匐茎和叶柄的病原菌为草莓炭疽菌,早期草莓炭疽病只是零星分布,后蔓延至整个佛罗里达州,经鉴定引起炭疽病的病原为草莓炭疽菌(Colletotrichumfragariae)。1965年,澳大利亚等地公布了草莓炭疽菌和尖孢炭疽菌(C.acutatum)的存在[4]。1978年Mass等[5]根据对马里兰的草莓病害研究调查认为草莓炭疽病病原主要为胶孢炭疽菌(C.gloeosporioides)、草莓炭疽菌和尖孢炭疽菌。近年来,炭疽病在草莓生长中危害日趋严重,已经成为草莓苗期甚至整个生长期的主要病害。设施环境下,草莓炭疽病大多不在地上部表现症状,而是在定植后的缓苗期危害根颈,引起根部腐烂,导致植株萎蔫和枯死。其致死率一般在10%~20%,有的可达到80%以上[6-7]。叶正文等[8]于1997年首次在我国报道草莓炭疽病并初步鉴定草莓炭疽病病原菌有尖孢炭疽菌和草莓炭疽菌2种,且以尖孢炭疽菌为主。2007年张海英等[9]经过鉴定发现引起抚顺草莓炭疽病的病原菌为胶孢炭疽菌和草莓炭疽菌。
尖孢炭疽菌与草莓炭疽菌通常对草莓地上部叶片及果实产生危害,地上部症状容易发现,可以及时防治。但胶孢炭疽菌引起的根部病害难以及时发现与控制,对草莓危害较重。本研究通过对草莓炭疽根腐病病原菌胶孢炭疽菌分离纯化发现,菌株在PDA平板上呈现不同的形态特征,因此对胶孢炭疽菌展开进一步的分析研究。胶孢炭疽菌分布地理范围广泛,寄主众多,具有复合种和种群集合的特征[10],传统的使用单一ITS序列鉴定真菌有时会出现混淆错误,结果不可靠。Crouch等[11]通过对炭疽菌ITS序列比对分析发现错误率可高达86%。因此本研究将通过菌落形态鉴定、分离纯化与分子鉴定、多基因序列(ITS-ACT-GAPDH-CAL-TUB2-CHS)联合鉴定以及致病性鉴定的方法,对我国草莓主产区的草莓炭疽病病原菌胶孢炭疽菌复合种进行系统发育与致病性分析,以期为病害防控提供科学依据。
1 材料与方法
1.1 材料
本研究于2017—2019年进行。样品采自草莓主产区山东临沂、山东聊城、河北怀来、辽宁东港、江苏徐州、安徽合肥、北京昌平、北京顺义、四川成都、甘肃定西、云南玉溪和山西晋城12个地区,采集草莓根腐病样品480株。经过对草莓病样的归类整理得到109株菌株进行后续研究(表1)。供试草莓品种有‘红颜’‘哈尼’‘宁玉’‘香野’‘妙香’‘甜查理’。

表1 草莓根腐病病样菌株采集信息Table 1 Collection information of strawberry root rot disease strains

表1(续)
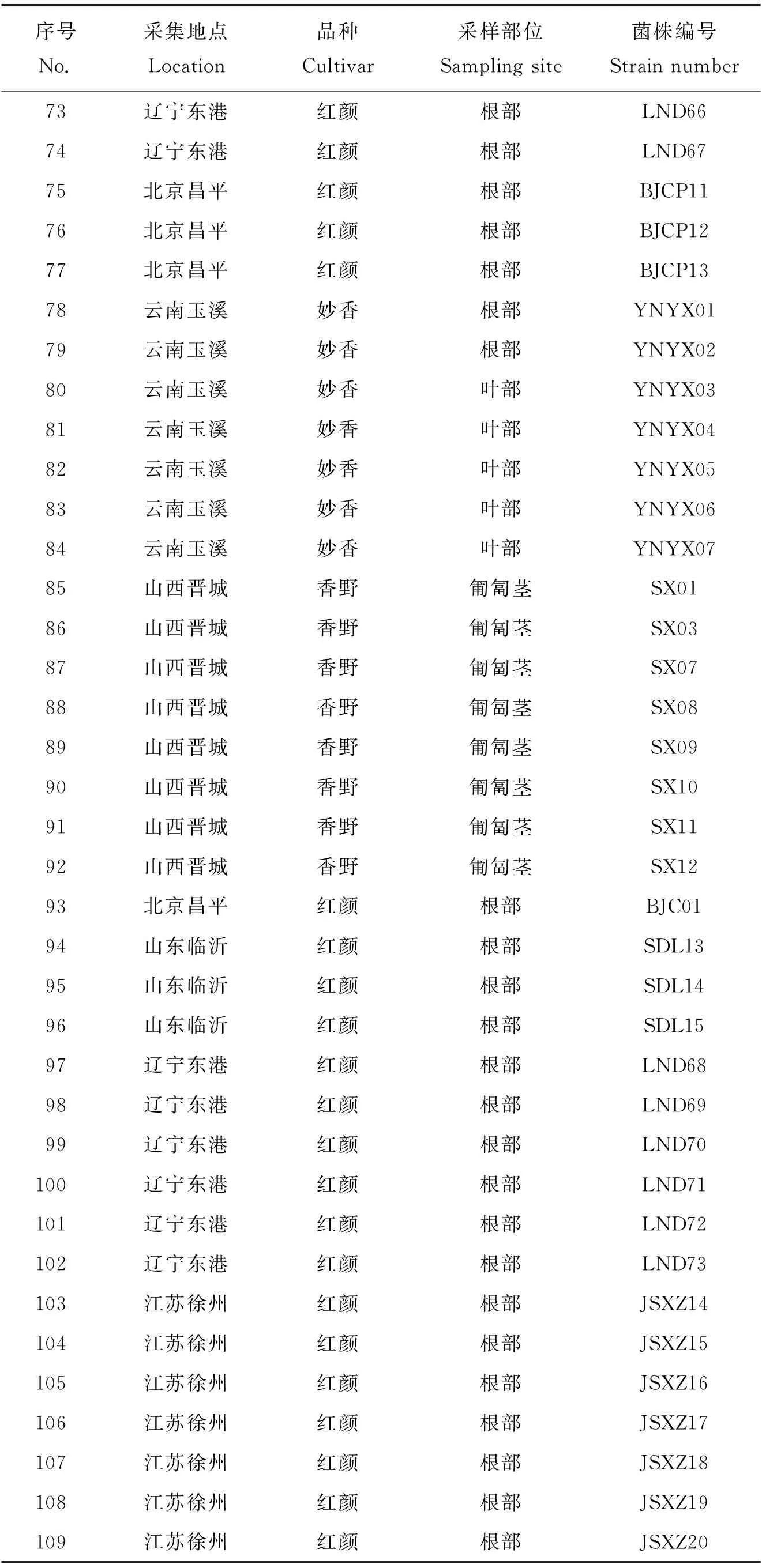
表1(续)
1.2 试验方法
1.2.1病原菌分离与纯化
取草莓根颈部病健交界处的组织(0.5~1 cm2左右),用75%酒精灭菌30 s,无菌水洗涤3次;之后用2%的次氯酸钠消毒6~8 min,无菌水洗涤3~4次;晾干置于PDA培养基,在25 ℃的恒温恒湿培养箱生长5~7 d。从菌落边缘处挑取菌落菌丝置于PDA液体培养基中,在25 ℃摇床中培养1 d。采用单孢划线分离法将单孢菌转移到PDA平板上用于鉴定和后续保存。
1.2.2DNA提取及多基因序列扩增
根据BufferA和BufferB溶液进行DNA的快速提取[12]。挑取少量菌丝放于装有50 μL BufferA溶液的PCR管中,之后置于PCR仪内,95 ℃裂解10 min;再向每个PCR管内加入50 μL Buffer B溶液,震荡混匀,12 000 r/min离心20 s,取上清液50 μL于新PCR管中,即获得所需DNA并测定其浓度。基于多基因序列各引物(表2)进行PCR扩增,反应体系(25 μL)包括:PCR mix 12.5 μL、上下游引物(10 μmol)各1 μL、DNA模板1 μL,加dd H2O 9.5 μL。PCR反应程序:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火45 s,72 ℃延伸1 min,30个循环;72 ℃延伸3 min。放于4 ℃保存。扩增产物使用1.5%琼脂糖凝胶电泳进行PCR检测,PCR产物送擎科公司测序。把测序成功的目的序列登录到NCBI数据库进行比对,下载同源性序列(表3)。用MEGA 7.0软件进行多重比对并修正[13],利用邻近法构建系统发育树[14]。

表2 各引物及PCR循环条件Table 2 Primers and PCR cycle conditions

1.2.3真菌致病性测定
选取生长状况一致的草莓苗‘红颜’进行喷雾法接种。将不同菌株的分生孢子制成1.0×106个/mL孢子悬浮液,均匀喷洒于草莓叶片上(以有水滴流下为度),以喷洒无菌水为对照,每处理喷洒5株,重复3次。接种后草莓植株置于培养箱(25 ℃)中黑暗覆膜保湿培养24 h。之后揭膜,在25 ℃,光周期为12 h/12 h下培养。
参考王丰等[15]方法,并稍加改动,对接种草莓苗病情分级与菌株致病性进行等级划分。0级:无病;1级:叶片枯萎面积占叶面积5%以下;3级:叶片枯萎面积占叶面积5.1%~15%;5级:叶片枯萎面积占叶面积15.1%~30%;7级:叶片枯萎面积占叶面积30.1%~50%;9级:叶片枯萎面积占叶面积50%以上或叶片枯死。发病率(DR)=发病株数/调查株数×100%,病情指数(DI)=[(各级发病株数×病级数)/(最高发病级数×调查株数)]×100。利用SPSS 26.0软件进行致病性分析,采用最小显著差数法(LSD)进行差异显著性检测,并完成柯赫氏法则验证[16]。
2 结果与分析
2.1 草莓炭疽根腐病田间症状
草莓定植初期,炭疽根腐病病原菌通过侵染草莓根部,分泌相应毒素。其孢子在维管系统中定植,破坏水分传导的木质部导管,导致植物萎蔫、干枯直至死亡(图1(a));切开病株短缩茎会发现根颈褐变、腐烂(图1(b))。

(a)感病植株萎蔫症状;(b)感病植株根颈横切面症状。(a) Symptoms of wilting of infected plants; (b) Transverse section of the Crown of infected plants.图1 草莓炭疽根腐病田间症状Fig.1 Field symptoms of strawberry anthracnose crown rot
2.2 草莓炭疽根腐病病原菌鉴定
2.2.1病原菌形态观察
暹罗炭疽菌菌株:在培养过程中菌落正面呈现白色,后期白色变暗,略微出现灰色;菌丝稀疏呈絮状,后期会产生孢子泥和分生孢子盘;菌落背面中间区域为黑褐色,从中间到边缘颜色逐渐变淡(图2(a))。分生孢子柱状、两端钝圆,尺寸为(11.73~13.99) μm×(3.00~5.02) μm(图2(b))。隐秘炭疽菌菌株:菌落正面呈现乳白色,菌丝较为致密;菌落背面呈白色,其中间区域出现较小的褐色圆圈(图3(a))。分生孢子柱状、两端钝圆,大小较为均一,尺寸为(16.56~17.90) μm×(4.32~5.50) μm(图3(b))。果生刺盘孢菌菌株:初始菌落正面边缘为白色,中间略微显现灰色,生长到第8天时,菌落正面出现大面积灰褐色,菌丝稀疏,气生菌丝较为茂盛,菌落背面出现大面积灰黑色(图4(a))。分生孢子柱状、两端钝圆,尺寸为(9.96~13.03) μm×(3.30~3.81) μm(图4(b))。暹罗炭疽菌附着胞(图2(c))、隐秘炭疽菌附着胞(图3(c))和果生刺盘孢菌附着胞(图4(c))分别由是相应的3种分生孢子在水中萌发芽管后形成的梨形和卵形等暗褐色结构,其形态稳定且大小不均一。其中暹罗炭疽菌和隐秘炭疽菌菌株中有一定数量的分生孢子会产生多个附着胞,隐秘炭疽菌的附着胞芽管偏短。

(a)菌落形态特征;(b)孢子形态特征;(c)附着胞形态特征。(a) Morphological characteristics of colony; (b) Morphological characteristics of conidia; (c) Morphological characteristics appressorium.图2 暹罗炭疽菌形态特征Fig.2 Morphological characteristics of C.siamense

(a)菌落形态特征;(b)孢子形态特征;(c)附着胞形态特征。(a) Morphological characteristics of colony; (b) Morphological characteristics of conidia; (c) Morphological characteristics appressorium.图3 隐秘炭疽菌形态特征Fig.3 Morphological characteristics of C.aenigma

(a)菌落形态特征;(b)孢子形态特征;(c)附着胞形态特征。(a) Morphological characteristics of colony; (b) Morphological characteristics of conidia; (c) Morphological characteristics appressorium.图4 果生刺盘孢菌形态特征Fig.4 Morphological characteristics of C.fructicola
2.2.2病原菌多基因序列联合鉴定
提取真菌基因组DNA后,采用真菌ITS1和ITS4通用引物序列,成功从不同菌株DNA样品中获得DNA扩增产物。以rDNA-ITS序列构建系统发育树,结果表明,分离纯化的109个菌株中有30个菌株归类于胶孢炭疽菌复合种。30个胶孢炭疽菌复合种可根据不同菌株形态特征分为3类。从3类菌株中各选1株代表菌株SCD26、LND07和SCD15进行多基因序列鉴定,构建胶孢炭疽菌的多基因联合系统发育树(图5)。结果发现胶孢炭疽菌代表菌株聚类为3个分支,其中SCD26病原菌聚类于暹罗炭疽菌;LND07病原菌聚类于隐秘炭疽菌;SCD15病原菌聚类于果生刺盘孢菌。

SCD15、SCD26和LND07为胶孢炭疽菌复合种代表菌株。SCD15, SCD26 and LND07 are representative strains of C.gloeosporioides complex.图5 胶孢炭疽菌复合种多基因联合系统发育树Fig.5 Phylogenetic tree of the C.gloeosporioides complex based on multi-gene combination
2.3 草莓炭疽根腐病病原菌致病性试验
通过对代表性菌株隐秘炭疽菌、果生刺盘孢菌和暹罗炭疽菌进行接种实验,发现在接种隐秘炭疽菌第9天、接种果生刺盘孢菌和暹罗炭疽菌第12和13天时草莓叶片开始发黄萎蔫,3种炭疽菌引起的病害程度差异显著。其中隐秘炭疽菌病情指数与致死率分别为82.2和66.7%,显著高于果生刺盘孢菌和暹罗炭疽菌。隐秘炭疽菌发病较快,引起病害较重。本研究采用叶片喷雾法接种病原菌,但在感染病原菌早期,草莓叶片并没有出现病斑,只出现叶片发黄萎蔫直到枯死,切开草莓短缩茎,其横截面出现大面积坏死褐变。隐秘炭疽菌感染的草莓,从短缩茎一侧向髓感染(图7(c)),而果生刺盘孢菌(图8(c))和暹罗炭疽菌(图6(c))感染的草莓,短缩茎髓处优先发病。按柯赫氏法则,将上述感病草莓进行组织分离,病原菌纯化,鉴定其病原菌为暹罗炭疽菌、隐秘炭疽菌与果生刺盘孢菌,与最初各接种的病原菌相同,即完成了柯赫氏法则。

(a)接种第1天;(b)接种第18天;(c)患病植株根颈横截面。(a) Day 1 of inoculation; (b) Day 18 of inoculation; (c) Cross section of rhizome of diseased plants.图6 暹罗炭疽菌致病性测定Fig.6 Determination of pathogenicity of C.siamense

(a)接种第1天;(b)接种第18天;(c)患病植株根颈横截面。(a) Day 1 of inoculation; (b) Day 18 of inoculation; (c) Cross section of rhizome of diseased plants.图7 隐秘炭疽菌致病性测定Fig.7 Determination of pathogenicity of C.aenigma

(a)接种第1天;(b)接种第18天;(c)患病植株根颈横截面。(a) Day 1 of inoculation; (b) Day 18 of inoculation; (c) Cross section of rhizome of diseased plants.图8 果生刺盘孢炭疽菌致病性测定Fig.8 Determination of pathogenicity of C.fructicola
3 讨论与结论
3.1 多基因联合鉴定的必要性
草莓整个生育期都可发生炭疽根腐病。在草莓育苗期和定植初期,发病率可高达90%以上,给健康草莓苗的繁育造成巨大破坏[21]。草莓炭疽病的鉴定对该病害的防治有重要的参考价值。由于病原菌菌落形态特征稳定性差且易受培养条件影响,单独依靠形态特征进行系统分析是不可靠的[22]。随着分子生物学的发展,基因序列鉴定成为一种常用的手段。目前草莓炭疽病的分类依据主要结合于前人的分类系统,以分生孢子和附着胞的形态特征、大小、纯培养特征以及ITS基因序列进行构建系统发育树来确定其归属。但是仅用1种基因的测序结果,一些亲缘关系较近的种仍不能进行有效区分,所以需要使用多基因序列进行物种进化的研究[23]。目前用于炭疽病鉴定分类的常用基因有ITS(核糖体DNA内转录间隔区)、TUB2(β-微管蛋白2)、ACT(肌动蛋白)、GAPDH(3-磷酸甘油醛脱氢酶)、CHS(几丁质合成酶)和CAL(钙调蛋白)等。Weir等[20]通过多基因联合建树的方法将胶孢炭疽菌复合种划分为22个种和1个亚种;宋丽丽等[24]利用多基因联合建树法对分离自上海和安徽的6个草莓炭疽菌菌株进行分子鉴定,发现这6个炭疽菌菌株均为果生刺盘孢菌;韩永超等[25]通过多基因联合分析明确了引起武汉地区草莓根颈腐病的病原为胶孢炭疽菌复合种内的暹罗炭疽菌;张方博[26]通过对草莓根腐病研究也发现,暹罗炭疽菌是草莓根腐病病原之一;Zhang等[27]对草莓炭疽病进行多基因联合鉴定发现隐秘炭疽菌也属于胶孢炭疽菌复合种。本研究发现草莓胶孢炭疽复合种目前包含3个小种即暹罗炭疽菌、隐秘炭疽菌和果生刺盘孢菌,与前人研究结果一致。
3.2 草莓炭疽根腐病优势菌种分析
同一寄主的炭疽病病原菌菌株之间,致病力存在差异。冯乐乐等[23]对浙江省鲜食大豆炭疽病病原平头炭疽菌进行分离鉴定,发现分离出的9株病原菌株虽同属于平头炭疽菌,但致病力却不同。郭艳春等[28]认为不同地区引起黄麻炭疽病的病原菌致病能力也不同。草莓胶孢炭疽菌复合种下的暹罗炭疽菌、隐秘炭疽菌和果生刺盘孢菌可在不同程度上引起草莓根部腐烂、地上部萎蔫和干枯等症状。Chen等[29]认为果生刺盘孢菌为浙江地区草莓炭疽菌的优势菌株,其致病力是胶孢炭疽菌复合种中最强的;Zhang等[27]研究结果表明在中国东部草莓栽培区中暹罗炭疽菌才是草莓炭疽病的优势菌株,致病力最强。本研究发现隐秘炭疽菌与果生刺盘孢菌和暹罗炭疽菌相比致病力最强。不同地区气候、土壤条件、栽培品种以及栽培措施不同可能造成草莓炭疽根腐病病原菌优势种不同,具体是由于何种原因导致优势菌株不同,还有待进一步研究。
本研究通过广泛采集全国不同草莓主产区根部腐烂样品发现,有27.5%的样品是由胶孢炭疽菌复合种引起。胶孢炭疽菌复合种可分为3种生理小种,分别为隐秘炭疽菌、果生刺盘孢菌和暹罗炭疽菌,其中隐秘炭疽菌发病较快,引起的病症较为严重。本研究还分离出43.4%的尖孢镰刀菌专化型(Fusariumoxysporumf. sp.fragariae)、11.1%的拟盘多毛孢(Neopestalotiopsisclavispora)以及其他病原菌,这些病原菌都可以加重草莓根腐病的发生。此外,关于草莓炭疽根腐病病原菌如何与其他病原菌复合侵染引起草莓病害也需进一步研究。
猜你喜欢
今日农业(2022年15期)2022-09-20
今日农业(2022年15期)2022-09-20
甘肃农业科技(2022年4期)2022-05-10
传播力研究(2020年20期)2020-11-11
大众摄影(2020年6期)2020-09-02
农民致富之友(2020年11期)2020-05-11
农民致富之友(2018年8期)2018-06-27
宠物世界·猫迷(2016年4期)2016-10-19
秀·媛尚(2013年8期)2013-10-12
