
载畜率对荒漠草原冷蒿植物种群密度及其空间分布的影响
2022-05-20 09:47王梓晗王忠武吕世杰闫宝龙王占文门欣洋宝音贺西格
生态学报 2022年8期
王梓晗,王忠武,*,吕世杰,闫宝龙,王占文,门欣洋,宝音贺西格
1 内蒙古农业大学草原与资源环境学院,呼和浩特 010018 2 内蒙古自治区农牧业科学院,呼和浩特 010018
内蒙古高原荒漠草原地处草原与荒漠两大陆地生态系统的交错带[1—2],这种草原植物群落的种类组成、群落类型及其结构和功能的独特性显示出了该生态系统的脆弱性[3]。加之人类不合理的活动行为,导致草地发生退化甚至促进草原荒漠化的进程。伴随着全球变暖等环境问题发生,草地保护研究和草原可持续发展的探讨仍是学者们关注的重点。
冷蒿(Artemisiafrigida)是菊科的一种小半灌木,兼性克隆植物,耐干旱、耐践踏、耐土壤侵蚀,生根萌蘖的再生生长能力很强[4—6]。冷蒿作为荒漠草原优势种之一,对群落的结构和功能也有着十分重要的影响,甚至决定着植物群落的稳定性。Zhang等的研究结果表明,冷蒿在退化草地中的优势地位排放的化感作用机制影响其他植物的恢复,尤其是在过度放牧的严重干扰下[7]。
短花针茅荒漠草原具有特殊的地理位置和脆弱的生态系统。殷国梅等发现放牧强度对短花针茅种群密度的影响最小,而对冷蒿种群密度的影响最大[8]。单玉梅等结果表明冷蒿种群总密度在放牧区均高于对照区,在放牧期间(6—9月)的冷蒿枝条总密度的峰值均出现在HG(或MG)区[9]。
关于放牧对植物群落空间异质性的研究已有文献报道。Zuo等的研究发现,内蒙古荒漠草原中灌丛的入侵在多尺度上简化了群落组成,放大了草本植被的空间异质性,并且荒漠草原草本植被空间异质性的尺度效应主要依赖于以灌木或草为主的植物群落[10]。刘红梅等发现短花针茅密度均值和分布的空间异质性最大变异程度都在自由放牧区[11]。殷国梅等的结果表明,随着载畜率的增大,短花针茅荒漠草原群落空间异质性虽有增加的变化趋势,但植物空间分布仍具有特异性[12]。研究者们对短花针茅荒漠草原建群种的关注度较高的原因是其控制植物群落的结构和功能。然而,前期试验研究发现[13],荒漠草原冷蒿伴随载畜率增加密度急剧下降,耐牧性和抗性与前人研究结果存在较大的差异。
本研究以内蒙古乌兰察布市四子王旗短花针茅荒漠草原优势种冷蒿为研究对象,探究了冷蒿植物种群在不同载畜率下的密度和空间分布特点,进一步研究冷蒿在放牧胁迫下的密度变化趋势和空间分布状态[14—16],研究结果有利于明确放牧干扰对冷蒿生态分布格局的影响程度,并对冷蒿植物种群空间消长过程进行初步阐释,为进一步深入研究荒漠草原冷蒿植物种群耐牧性和抗性提供基础,也为荒漠草原生态系统的保护和防止荒漠草原退化给予一定的试验和理论指导。
1 试验研究内容与方法
1.1 试验地概况
试验地位于内蒙古自治区四子王旗王府一队,地理位置为41°47′17″N,111°53′46″E,海拔的平均高度为1450m,地处中温带大陆性季风气候,气候特点是春温骤升、秋温巨降,无霜期短,平均无霜期约为108d。近10年平均降水量在220mm。土壤为淡栗钙土。草地类型为短花针茅(Stipabreviflora)+冷蒿(Artemisiafrigida)+无芒隐子草(Cleistogenessongorica)荒漠草原。植被稀疏,植物种类组成比较简单,草层盖度较低。
1.2 试验设计
放牧试验区建于2004年,采用随机区组设计,如图1所示,3个区组,每个区组又包含4个不同的载畜率,即对照区CK(0只/hm2)、轻度放牧区LG(0.93只/hm2)、中度放牧区MG(1.82只/hm2)和重度放牧区HG(2.71只/hm2),各处理区随机排列[17]。放牧绵羊品种为蒙古羊。每日放牧时间为6:00时放牧,18:00时归牧。放牧时段为每年6—11月底,放牧时间为6个月。
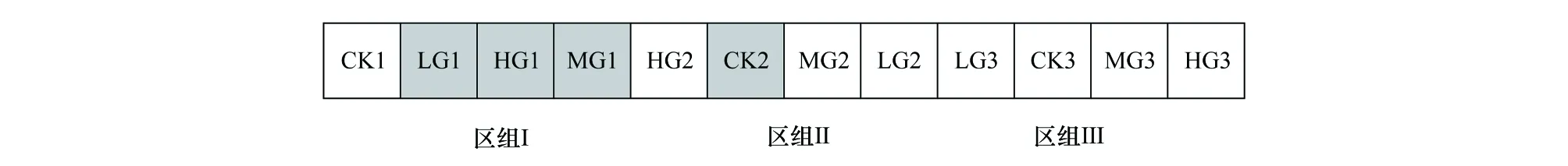
图1 试验小区和区组示意图Fig.1 Schematic diagram for experimental plot and blockCK:对照 Control check;LG:轻度放牧 Light grazing;MG:中度放牧 Moderate grazing;HG:重度放牧Heavy grazing;图中标记颜色的小区为固定样地取样区
1.3 研究方法
为研究荒漠草原冷蒿种群密度及其空间分布特征,本试验从每个载畜率相同的3个重复区中选定4个载畜率不同的代表性重复区进行试验。在每个不同载畜率处理样地内,各选择一块地形一致、与小区门口距离接近的代表性样地(面积为40m×40m)。采用机械取样法进行取样试验,将样地边界中的西南交汇处定为取样坐标原点(0,0),每10m设置一个0.5m×0.5m的样方,距离原点处最远的坐标为(16,16),并按照偶数行取5个样方,奇数行取4个样方的方法进行取样,因此每个样方地的样方数为77个(样点分布见图2,图中每一黑色方块代表一个50cm×50cm样方),于2017—2019年8月15日左右测定每个样方中冷蒿植物种群的密度。
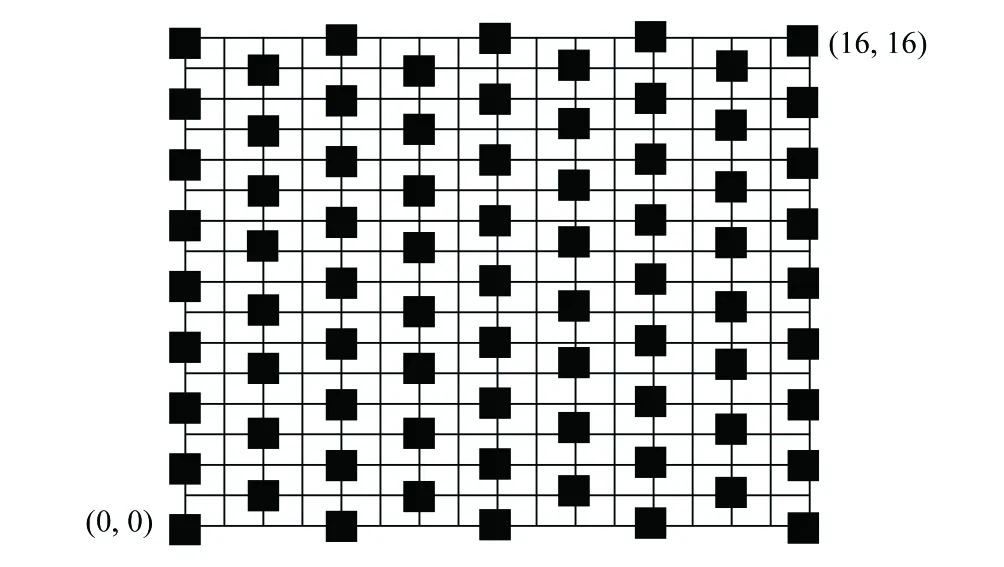
图2 固定样地取样样点分布情况Fig.2 Distribution of sampling points in fixed sites
本次试验结果采用SAS 9.4软件对2017—2019年8月冷蒿密度进行描述性统计和方差分析,并用地统计学软件GS+9.0进行空间异质性统计分析(首先构建最优拟合模型,并通过半方差函数和分形维数等方法分析其植物群落空间分布特点),箱线图和多重比较结果采用Sigmaplot 12.5进行图形绘制[18-19]。其中描述性统计调用MEANS过程,方差分析调用GLM过程。
2 结果与分析2.1 冷蒿种群密度对载畜率的响应
在不考虑样本空间位置及其样本间的间隔距离的条件下,不同年份下,冷蒿密度随着载畜率增加而下降(图3)。
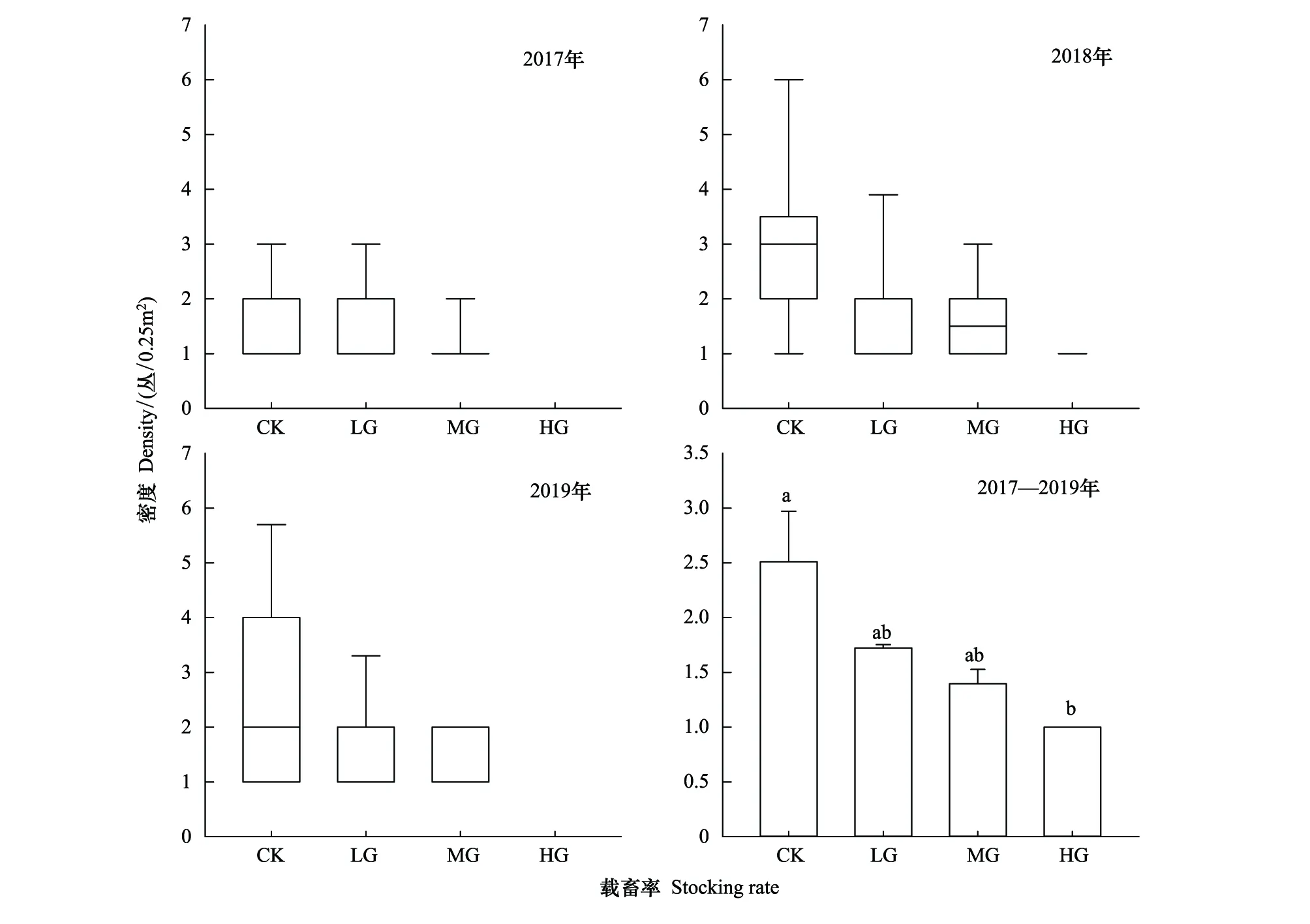
图3 冷蒿在不同载畜率下种群密度的变化图Fig.3 The variation of population density of Artemisia frigida under different stocking rates柱形图中字母不同表示不同处理间差异显著(P<0.05),字母相同表示差异不显著(P>0.05)
从年份上看,2017年冷蒿植物种群密度CK和LG区的冷蒿密度最大值(箱式图的上侧箱线)和上四分位数(箱体上侧的边缘)之间无差别,说明CK和LG区冷蒿密度的变动范围相近,MG和HG区的显示结果是由于冷蒿在MG和HG处理区分布较少(77个样点出现的频次较少)造成的结果;2018年,CK区冷蒿密度的最大值和最小值(箱式图的下侧箱线)之差即冷蒿样点间的变动幅度差别最大,说明冷蒿植物种群密度在该处理区分布较大,LG和MG处理区最大值有差异;2019年,CK和LG区的冷蒿密度分布较为分散,但CK区的密度变化程度均明显大于其他3个处理区,随着载畜率的增大,箱式图的箱体(中间矩形区域)逐渐减少,表明放牧强度增大导致冷蒿植物种群密度的集中程度增加,也就是其密度值落在上下四分位数之间的变化范围在减少,意味着家畜的选择性采食和践踏强度增加导致种群空间分布均匀程度降低,结合箱体和中位数可以看出,冷蒿密度空间分布随着载畜率的增加,其密度空间分布在降低。因此,2017—2019年这3年中随着载畜率增大,冷蒿的密度呈现出下降的变化趋势(P<0.05)。
将2017—2019年的不同载畜率下的冷蒿植物种群密度加权平均后进行方差分析,CK与HG处理区的差异性显著(P<0.05),HG区表现最低,CK区表现最高;在不同载畜率下,冷蒿植物的种群密度由大到小依次为CK>LG>MG>HG。同时,我们也发现,尽管冷蒿植物种群密度在MG处理区小于LG处理区,但是MG处理区年份之间的冷蒿密度的波动程度较LG处理区大。
2.2 冷蒿植物种群密度在不同载畜率下的变异
冷蒿在40m×40m样地范围内的变异分析如图4所示。从变异系数的角度来看(样本容量n=231),CK、LG和MG区的CV值分别为62.68%、55.19%、34.36%,HG区由于冷蒿数量急剧下降,无法计算变异系数(na)。因此,伴随载畜率增大冷蒿植物种群密度单位均值上的变异程度在下降。从冷蒿在样地出现的频率角度分析,冷蒿植物种群在CK区出现频率(P值)最大,在HG区出现的频率最小。由此可见,冷蒿种群出现的频率也在急剧下降。图4中的冷蒿密度的均值(虚线)也是随着载畜率的增大呈现出下降趋势。综合来看,载畜率增大导致冷蒿植物种群密度下降,单位均值上的变异程度下降,出现的频率下降。
2.3 冷蒿空间分布的地统计分析
对不同载畜率下各样点处的冷蒿密度进行变异函数分析,发现不同载畜率下冷蒿的变异函数值均呈现出理论模型的变化趋势。结果显示(表1),冷蒿在CK区中的变异函数值呈现指数理论模型的变化趋势;LG和MG区为球状理论模型;HG区为线性理论模型。如图5所示,半方差函数的数值随着空间取样分隔距离的增加而逐渐增大,但当达到某一特定分隔距离时,函数值逐渐趋于稳定状态。
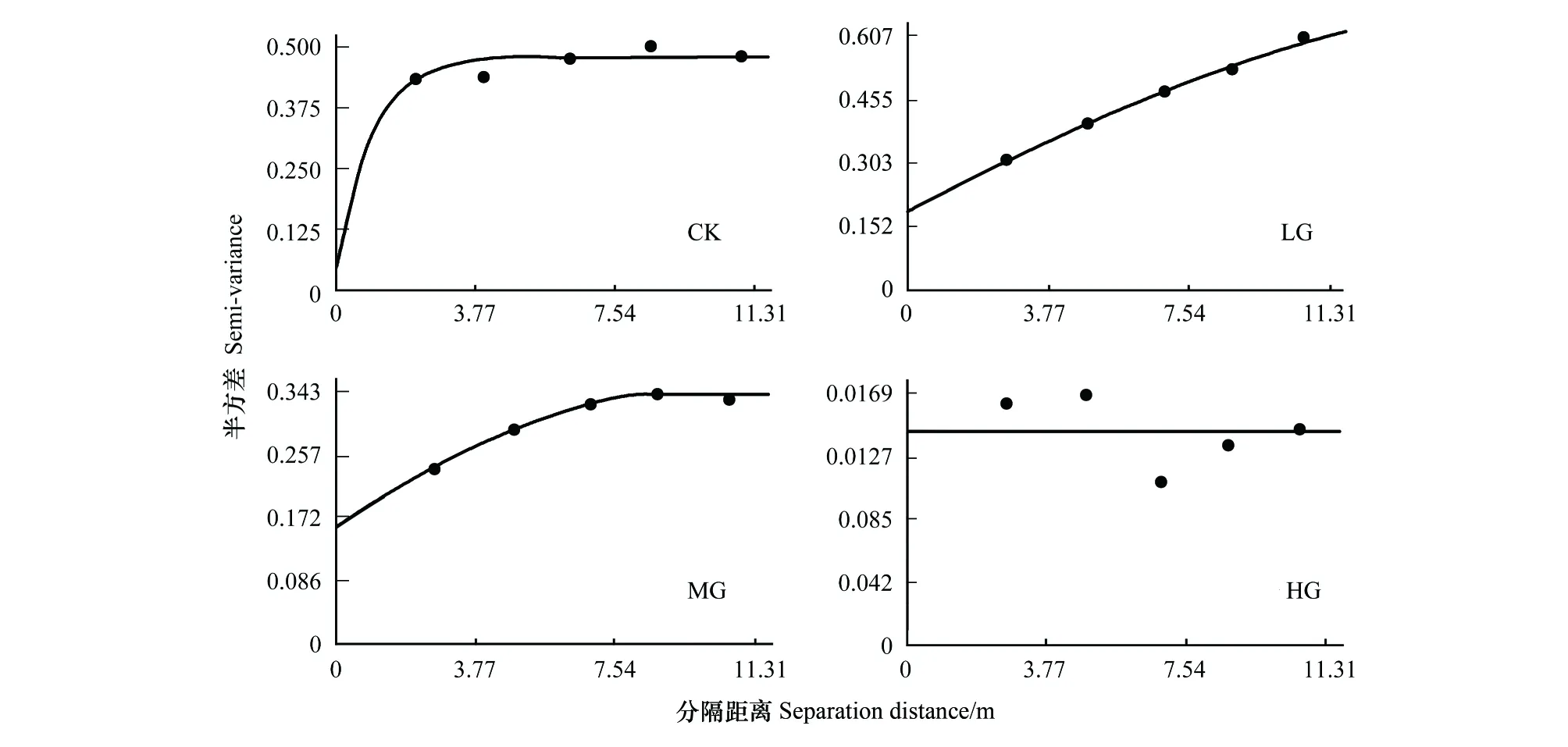
图5 冷蒿空间格局的变异函数Fig.5 The variogram of spatial pattern of Artemisia frigida
从半方差函数中的数值来看,块金值C0表示随机因素而导致的空间变异,LG和MG处理区的块金值较大,CK和HG区的块金值较小,说明冷蒿在LG和MG区中受随机因素的影响较大,在CK和HG区中受结构性因素影响较大。基台值C0+C是样地范围内最大的空间变异值,冷蒿密度分布的空间变异程度由大到小的顺序为LG>CK>MG>HG。结构比C/(C0+C)按放牧强度由小到大依次为0.935、0.712、0.530和0.000,这说明随着载畜率增加,冷蒿的空间相关性逐渐减弱。
在半方差函数分析的基础上,进一步对斑块参数进行研究。空间自相关距离因模型不同而存在计算差异,指数模型主轴变程和亚轴变程为表1中A0数值的3倍,球形和线性模型空间自相关距离在数值上即为表1中A0的数值[20]。由此可以看出,LG区的冷蒿密度的自相关范围最大,为164.8m;次之是HG和MG区,分别为106.2m和87.8m;冷蒿密度的自相关范围最小的是CK区,值为28.5m。分形维数D0表示结构空间分布因子在最大空间变异中的比例,在不同放牧情况下,分形维数的数值都是位于1.700和2.000之间,说明这4个不同载畜率的区域,其空间分布都是较为均匀的。
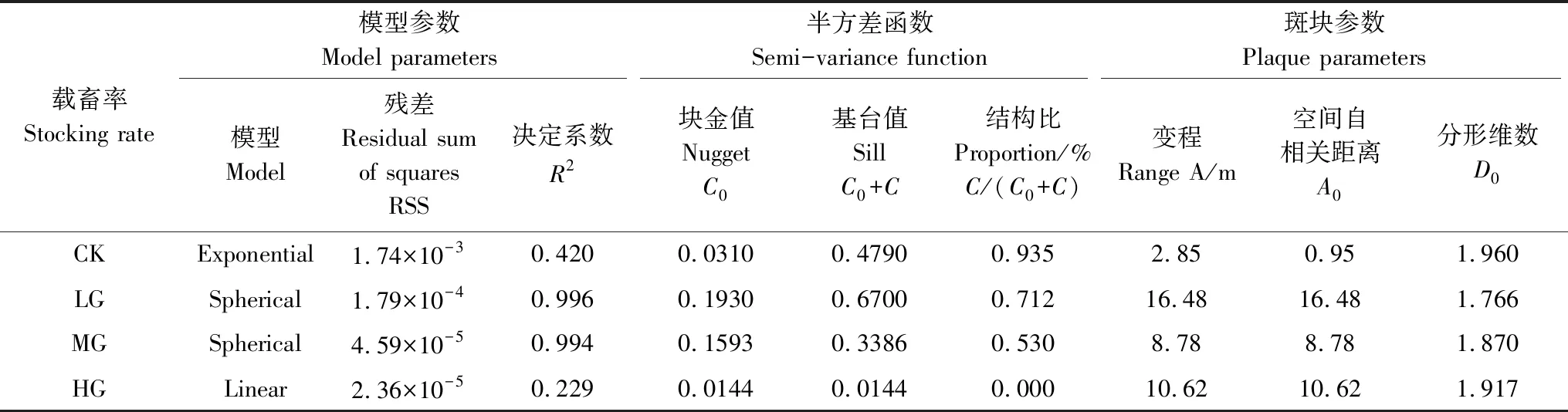
表1 冷蒿空间分布的地统计分析表Table 1 Statistical analysis table of spatial distribution of Artemisia frigida
2.4 冷蒿在不同载畜率下的空间分布状态
研究冷蒿空间分布状态采用克里格(Kriging)插值法。由图6可以看出,CK区呈现明显的2种斑块状的地理分布,上方斑块分布密度较小,下方斑块分布密度较大,这2类斑块镶嵌排列,其空间分布状态表现为网状结构。LG处理区出现2种带状分布,上边的带状分布是密度由内向外依次呈现出逐渐递减的规律,下边的带状分布是密度由内向外依次呈现出逐渐递增的趋势。MG区的上部分的冷蒿分布的密度由内向外呈现出逐渐递减的趋势,下部分则呈现出2处大斑块的分布,并以大斑块向四周递减。相比较而言,LG和MG区的密度分布相似,但MG区的均匀性要大于LG区。HG处理区分布比较单一,只在西北角有部分分布,其他地方几乎没有冷蒿分布的现象,这是由于过度放牧导致冷蒿密度急剧下降造成的。
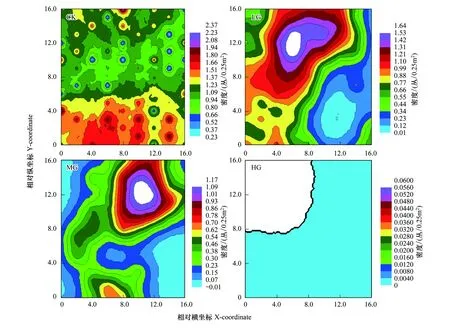
图6 冷蒿空间分布格局平面图Fig.6 Spatial distribution plan of Artemisia frigida
3 讨论
3.1 冷蒿密度对载畜率的响应
草原植物群落特征与载畜率紧密联系,冷蒿作为植物群落的优势种也不例外,在大气候一致时,放牧强度会成为主导因子,控制和影响着植物群落的特征[21—23]。杜利霞等人研究了不同放牧条件下冷蒿的种子繁殖和营养繁殖特性后,认为冷蒿在轻牧区和封育区单位面积种子产量较高,其营养繁殖的各构件在轻牧区表现最好,能促进冷蒿的营养繁殖更新[24]。本研究也认为轻度放牧不会改变其在植物群落中的地位和作用,这可能是有性繁殖补充了因放牧导致的种群数量衰减程度。Ye等的研究表明,短花针茅荒漠草原的植物对放牧干扰的抗性和恢复力主要依赖于降水条件,适度干扰有利于其生长[25]。冷蒿在荒漠草原具有一定的耐牧性,从方差分析可看出,CK与LG区的密度之间差别较MG与HG区的密度之间差别大,说明冷蒿适应了其放牧强度,因为冷蒿有为躲避家畜采食而调整的适应策略,并且作为多年生的草本植物,其枝条性别化是有性生殖的基础,可以直接影响到植株的数量,这不仅受其自身遗传特性的控制,也受环境条件的制约[26]。杜宇凡研究放牧强度对短花针茅荒漠草原群落特征的影响后,发现冷蒿在不同放牧强度下没有出现消失的迹象,但冷蒿出现的频率有降低的趋势。此研究与本研究结果一致,这说明荒漠草原的这种格局经过漫长时间形成后相对稳定[27]。臧晓琳的研究认为冷蒿可以改善土壤微环境,增加其根际土壤微生物群落功能多样性,促进地上植株生长,增强其抵抗放牧胁迫的能力[28],这可能是冷蒿植物种群适应轻度放牧或围封条件的地下适应过程。有学者认为,随着载畜率的增加,冷蒿通过增加根系的生物量来保证吸收足够的养分和水分供给地上部分的生长,使得其更加适应环境,促进扩散[29]。在有环境胁迫下,冷蒿具有极强的抗旱性、较高的物质生产量和节约用水的能力,其自身积累的大量脯氨酸、可溶性糖等有用物质,使得冷蒿不断有新枝萌发出来,第二年春仍能旺盛生长[30]。但在试验区,中度放牧和重度放牧导致冷蒿密度急剧下降,进而导致草地裸露面积增大,使得土壤风蚀程度增加[31],难以使植株匍匐枝产生不定根,无性繁殖受限导致其株丛扩展能力下降。这是冷蒿植物种群不能适应中度放牧和重度放牧的无性繁殖基础,也是影响有性繁殖的重要原因之一。
3.2 冷蒿空间分布对载畜率的响应
冷蒿的空间分布格局可以反映其在载畜率胁迫下退化演替阶段。根据克里格插值法对不同放牧区的冷蒿空间分布进行绘制,得到了空间分布图,可呈现出冷蒿空间分布的异质性和复杂性,显示出其斑块性、梯度性和镶嵌性的分布特点[32]。熊梅在短花针茅荒漠草原主要植物与土壤空间格局对载畜率的响应中发现,随着载畜率增加,冷蒿的聚集强度提高,生态位变窄,并且冷蒿在CK区空间分布呈斑块状分布,其空间自相关性增强[33]。这与本研究结果一致,在CK 区不仅呈现斑块分布,而且其分形维数的数值最大,说明冷蒿植物种群空间分布更加均匀,空间自相关性更好。随着载畜率的增加,冷蒿植物的种群空间分布变得复杂,放牧家畜的采食、游走、践踏等随机性牧食行为都会导致冷蒿空间分布的随机性增强,到了重度放牧区,被放牧牲畜采食后的冷蒿所剩无几,才会出现在HG区分形维数的数值大于LG和MG处理区的结果,并且在放牧胁迫下,冷蒿在资源竞争中处于劣势地位,为适应放牧而采取了主动防御措施,并且以聚集分布的形式抵抗外界干扰以维持自身的稳定性。刘先华等以内蒙古温带典型草原的物种为研究对象,探究载畜率对内蒙古典型草原物种分布空间异质性的影响,其研究结果表明,随着载畜率增大,冷蒿的空间分布随机性和自相关尺度逐渐增大,但载畜率超过一定水平后,冷蒿空间分布的自相关尺度开始下降[34]。本研究结果与之存在较大差异。首先,从随机因素引起的空间变异程度来看,符合刘先华等研究结果变化趋势,但本研究的指标是块金方差(随机因素影响程度);其次,从结构比来看,结构比反映了结构部分空间异质性(由地形、土壤母质、气候等结构性因素引起的空间变异)占总空间异质性的程度[35],结构因素对冷蒿植物种群空间分布的影响伴随载畜率增大而减小,取而代之的是家畜随机性牧食行为对其影响程度的增加,即随机性因素决定着冷蒿植物种群在中度和重度放牧区的空间分布特点;最后,空间自相关距离反映的是试验区冷蒿植物种群空间分布的斑块大小,自相关距离越大,表明冷蒿植物种群空间分布斑块越大,因此LG和HG处理区斑块性较大。但这并不代表冷蒿植物种群空间分布的斑块都比较大,LG属于放牧增加冷蒿空间分布斑块尺度(可以从分形维数和结构比看到),但HG却表征的是空地面积(可以从分形维数和密度数据可以看到);所以尽管分析的是冷蒿植物种群空间分布状态,但需要结合多个统计参数和冷蒿实际密度情况进行综合判断。
4 结论
研究结果表明,随着载畜率的增大,荒漠草原优势植物冷蒿的密度下降,分布范围减小。冷蒿植物种群空间变异的比重和空间自相关随载畜率增大而逐渐减弱。在不同的载畜率下,冷蒿存在着独特的空间分布斑块特征,围封区呈现斑块状分布,轻度放牧区呈现带状分布,中度放牧区呈现斑块状和带状混合分布,而在重度放牧区分布较单一。为进一步了解和揭示冷蒿种群在荒漠草原的分布机理,今后还需通过长期的野外观测,深入探讨影响冷蒿分布的其他相关生理指标,为荒漠草原的可持续管理提供理论支撑。
猜你喜欢
今日农业(2022年15期)2022-09-20
小哥白尼(趣味科学)(2022年5期)2022-08-15
自我保健(2021年2期)2021-11-30
妇女之友(2021年9期)2021-09-26
湖南电力(2021年1期)2021-04-13
昆明医科大学学报(2020年11期)2020-12-28
绿色中国(2019年14期)2019-11-26
百姓生活(2019年2期)2019-03-20
红土地(2018年7期)2018-09-26
儿童故事画报·智力大王(2016年7期)2017-02-08
