
天目山常绿阔叶林土壤养分的空间异质性与植物组成之间的关系
2022-05-19 12:58徐钧杰陈旭刘宗悦庞春梅余树全
浙江林业科技 2022年3期
徐钧杰,陈旭,刘宗悦,庞春梅,余树全
(1.浙江农林大学 林业与生物技术学院,浙江 杭州 311300;2.浙江天目山国家级自然保护区管理局,浙江 杭州 311311)
由于受到气候因素、成土母质、地形地貌、生物等诸多方面的影响,不同地区的土壤性质存在明显的差异,这种差异被称为土壤的空间异质性[1]。从大尺度上看,对土壤的空间异质性起决定性作用的是土壤母质和气候因素,而在较小的尺度上,植物群落的组成以及种群分布是导致土壤异质性的主要原因[2]。植物群落与土壤空间异质性之间的相互影响、相互作用越来越成为生态学研究的重点。土壤的空间异质性不仅对土壤的结构功能有重要的影响,而且与植物的空间分布和群落组成也有密切的联系[3]。越来越多的研究表明[4-6],深入研究土壤性质和对植物生长以及植物群落组成结构等相互作用关系,对揭示生态系统结构与功能、生物多样性维持机制等具有重要意义。
近年来,国内外研究者利用地统计学方法对受人为干扰的林分和人工林进行土壤空间异质性的相关研究较多[7-9],而对于亚热带自然森林生态系统的相关研究开展较少。亚热带常绿阔叶林因其复杂的群落结构、丰富的生物多样性、良好的森林生态效益以及具有对维持全球性碳循环有积极作用等特点,在全球生态系统中占据着独一无二的地位[10]。常绿阔叶林是亚热带湿润地区典型的地带性森林植被类型,我国拥有丰富的亚热带常绿阔叶林资源,但由于以往认识不全面,以及长期的人类活动和高强度资源利用,使得森林面积减少,森林生产力退化,生物多样性降低。这些现象愈发严重,所以对森林生态系统的保护以及对已破坏林分的恢复重建工作值得每个人重视[11]。
本研究以浙江省天目山国家级自然保护区内的常绿阔叶林作为研究对象,通过野外群落调查和土壤采样分析,建立植物群落组成和土壤养分数据库,运用地统计学和Kriging 插值等方法对天目山常绿阔叶林土壤养分的空间异质性与植物组成的关系进行分析研究,旨在更好地理解天目山常绿阔叶林土壤养分的空间分布特点,揭示土壤养分与植物组成的相互关系,为该区域的生物多样性保护和生态系统的恢复提供指导。
1 研究区概况
浙江天目山国家级自然保护区位于浙江省西北部杭州市临安区境内的西天目山,地理坐标为30°18′30″~ 30°24′55″N,119°23′47″~ 119°28′27″E,海拔为300~ 1 556 m。天目山气候属中亚热带向北亚热带过渡型,年平均气温在8.8~ 14.8℃,年降水量达1 390~ 1 870 mm。该区植被类型多样,随海拔高度呈垂直地带分布,在海拔230~ 850 m 分布常绿阔叶林,在海拔>850~ 1 100 m 分布常绿落叶阔叶混交林,在海拔>1 100~ 1 350 m 分布落叶阔叶林,在海拔1 400 m 以上分布落叶矮林。天目山土壤随着海拔升高由亚热带红壤向湿润的温带型棕黄壤过渡,在海拔600 m 以下为红壤土,在海拔>600~ 850 m 主要为黄红壤土,在海拔>850~ 1 200 m 为黄壤土,在海拔1 200 m 以上为黄棕壤[12]。
2 材料与方法
2.1 样地设置与植物调查
在天目山国家级自然保护区内选择地形较为均匀、海拔为588 m 且面积为1 hm2的常绿阔叶林区域,并将其分为25 个20 m×20 m 样地。对样地内所有胸径≥1 cm 的植物个体进行树种识别、空间定位、挂牌并测量胸径、树高、枝下高以及冠幅等,建立植物群落组成特征数据库。根据样地内各树种的重要值大小,确定主要优势树种依次为紫楠Phoebe s heareri、杉木Cunninghamia l anceolata、榧树Torreya gr andis、细叶青冈Cyclobalanopsisgracilis和青钱柳Cyclocarya paliurus等。
2.2 土壤采样与分析
2.2.1 土壤采样点分布设计 2020 年10 月,对样地的土壤进行调查。首先,在每个20 m×20 m 样地四角上采取0~ 10 cm 表层土样,然后,随机选取4 个点中的1 个,从该点随机取一个指向样方内部的方向,在距离其2 m、8 m 或20 m 的三个点随机取一个点同样采取0~ 10 cm 表层土样,这样保证在每个20 m×20 m 样地内都有一个随机土壤采样点,共计采集61 个土壤样品。
2.2.2 土壤采样与预处理方法 在上述确定的61 个采样点上,用土钻法采集0~ 10 cm 表层鲜土,装入自封袋带回实验室,测定土壤含水量,土样经过自然风干、除去杂质、研磨、过0.149 mm 筛等预处理后装袋,用于测定。
2.2.3 土壤分析测定方法 土壤测定参照鲍士旦的《土壤农化分析》[13]。pH 采用电位法测定;有机质含量用重铬酸钾容量法-外加热法测定;全氮含量用硫酸-催化剂消解法测定;全磷含量用氢氧化钠熔融-钼锑抗比色法测定;全钾含量用氢氧化钠熔融法-火焰光度法测定;碱解氮含量用碱解-扩散法测定;速效磷含量用盐酸和硫酸浸提-钼锑抗比色法测定;速效钾含量用醋酸铵浸提-火焰光度法测定。
2.3 数据处理
地统计学中以半方差函数为基本工具来定量描述土壤性质的空间变异结构,并通过变异函数模型来表达这种关系,为Kriging 插值提供输入参数[14]。在进行分析前,需检验土壤养分数据是否符合正态分布,对不符合的数据进行对数转换或平方根转换以满足正态分布。半方差函数公式如下:
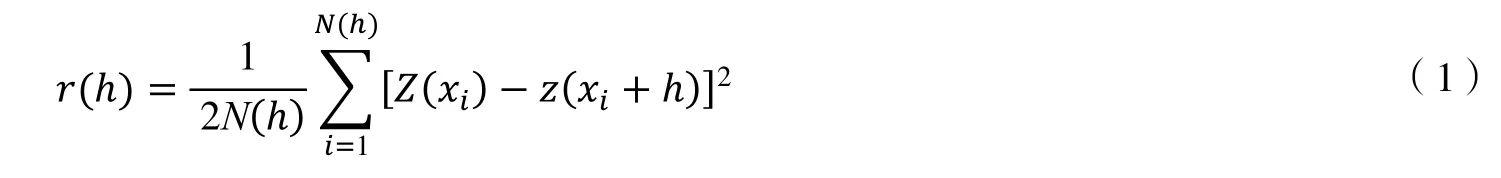
式(1)中,h为步长也称为位差;r(h) 是步长为h时的半方差函数值;N(h) 是步长为h时成对采样点的总数;Z(xi)、Z(xi+h) 表示在xi和xi+h位置上的土壤养分含量值。
分形维数(D)可以提供被研究对象空间格局的尺度及层次性和空间异质性在不同尺度间的相互关系等方面的信息[15]。分形维数越大,空间格局的空间依赖性就越弱,结构越差,空间格局就相对复杂。分形维数计算公式为:

式(2)中,m为双对数回归曲线的斜率。
各土壤养分半方差函数的计算以及理论模型的拟合在GS+9.0 软件中进行,并得到决定系数(R2)、块金值(C0)、基台值C0+C、块基比(C0/C0+C)等数据。以此为基础在ArcGIS10.3 软件中的Geostatistical Analyst地统计模块进行Kriging 插值。
采用Excel 2016 和SPSS 22.0 软件整合初始数据并进行描述性统计分析,计算平均值、极值、标准差、变异系数等指标,以评价土壤养分的变异程度。计算每个20 m×20 m 样地的生物多样性指数以及不同生活型树种重要值的占比,以此为植物组成数据。将Kriging 插值做出的养分空间分布格局图通过Arcgis10.3 计算出各样地的土壤养分平均值,用上述数据在SPSS 22.0 中对植物组成与土壤养分进行Pearson 相关性分析。其中,生物多样性指数按公式(3)、公式(4)进行计算,重要值数据按公式(5)进行计算。研究区中重要值排名前20 位树种的重要值之和达到86.7%,能较好地反映研究区内的植被情况,所以建立20×25 的树种重要值数据矩阵和8×25 的土壤养分数据矩阵,在Canoco 5.0 软件中进行冗余分析(Redundancy analysis,RDA),研究不同树种分布与土壤养分之间的关系。
Shannon-Wiener 多样性指数(H′)计算公式为:

式(3)中,S为物种数,Pi为第i个种的个体所占比例。
Margalef 丰富度指数(M)计算公式为:

式(4)中,S为物种数,N为所有物种个体数之和。
重要值计算公式为:

式(5)中,相对高度=(单个种的总高度÷所有种的总高度)×100%;相对多度=(该种的数量÷所有种的数量)×100%;相对显著度=(该种个体胸高断面积的和/所有种的胸高断面积的和×100%。
3 结果与分析
3.1 土壤养分含量的统计分析特征
土壤养分含量的描述性统计分析结果如表1。变异系数(CV)在一定程度上可以反映区域化变量的离散程度,CV>30%为高度变异,10%≤CV≤30%为中等变异;CV<10%为弱变异[16]。由表1 可知,研究区内土壤pH的平均值为3.96,呈强酸性,变化范围为2.84~ 5.27,极差大,CV为12.63%,属中等变异;土壤养分含量的变异也大,有机质、全氮、全磷、碱解氮含量的CV在26.62%~ 28.89%,均属强中等变异,全钾含量属弱中等变异,而速效磷和速效钾含量的CV分别达43.1%和30.33%,为高度变异。总体来看,天目山研究区常绿阔叶林土壤养分含量和pH 变异大。
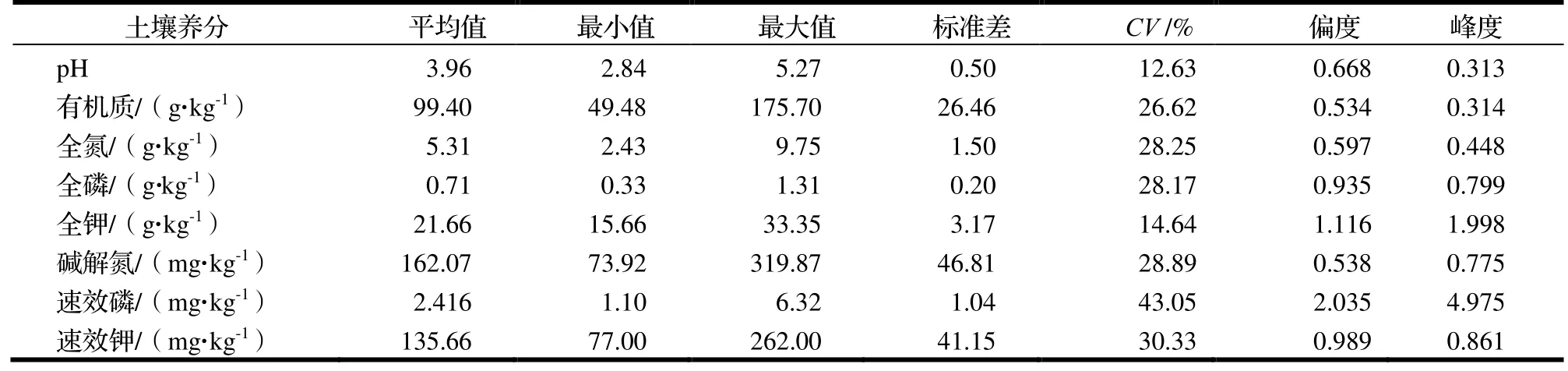
表1 研究区土壤pH 和养分含量的统计特征Table 1 Soil pH and nutrient contents in sample plots
3.2 土壤养分的空间变异性
由表2 可知,各土壤养分含量的R2在0.501~ 0.887,除有机质和碱解氮含量的R2相对较低以外,其他指标与理想模型的拟合度均较高,能较好地反映研究区内土壤的空间结构特征。土壤pH 和全磷、碱解氮和速效钾含量的最佳拟合模型为球状模型,有机质、全氮和速效磷含量的最佳拟合模型为指数模型,全钾含量的最佳拟合模型为高斯模型。
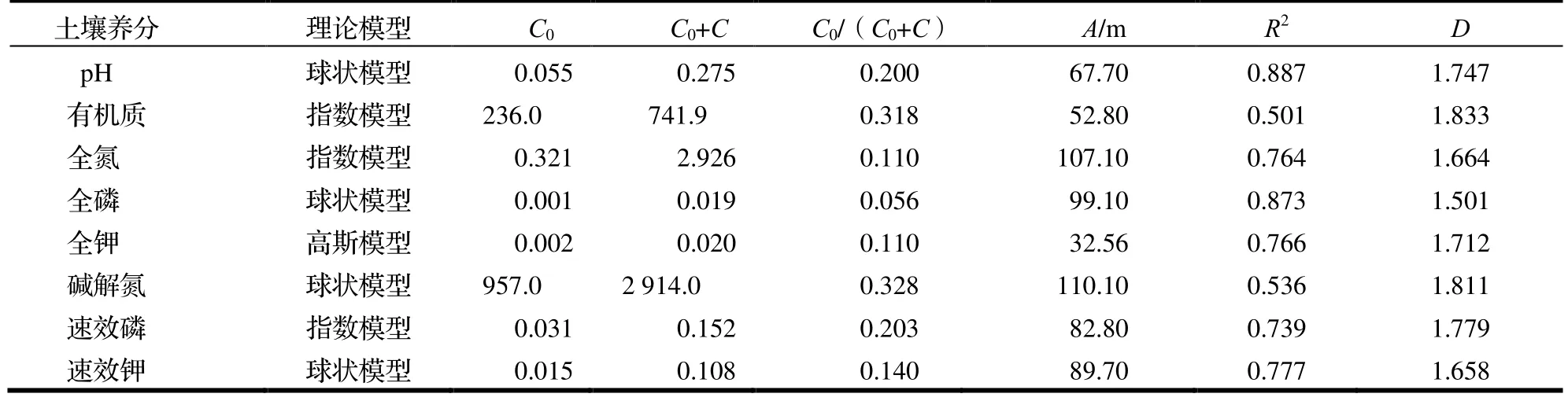
表2 研究区土壤pH 和养分变异函数理论模型及其相关参数Table 2 Variation function models and parameters of soil pH and nutrient content in sample plots
块金值(C0)是步长为0 时的半方差值,基台值(C0+C)是半方差的最大值[14]。土壤有机质与碱解氮含量的块金值分别为236 和957,基台值分别为741.9 和2 914,均明显大于其他土壤养分指标。这表示土壤有机质与碱解氮含量在研究范围内的随机性强,在小于现有采样尺度下有其他生物或非生物过程产生影响。
Cambardella 等[17]认为块基比<0.25 时,存在强烈的空间自相关性;块基比在0.25~ 0.75 时,具有中等程度的空间自相关性;当块基比>0.75 时,空间自相关性较弱。研究区土壤pH 和全氮、全磷、全钾、速效磷和速效钾含量的块基比较小,均<0.25,表现出变量存在强烈的空间自相关性。土壤有机质、碱解氮含量的块基比分别为0.318 和0.328,在0.25~ 0.75 之间,表明变量有强中等程度的空间自相关性。各土壤养分含量块基比由小到大依次为全磷、全氮、全钾、速效钾、pH、速效磷、有机质和碱解氮。
变程(A)表示土壤养分含量的空间相关范围,如果采样点之间的距离小于变程时,说明土壤养分含量之间存在一定的空间自相关性,反之则相互独立。其中,碱解氮含量的变程最大,为110.1 m,表明碱解氮相对于其他土壤养分在研究区内分布较为简单,空间变异弱,空间连续性较好,而全钾含量的变程仅为32.563 m,空间连续性相对较差,空间变异强。
从分形维数(D)来看,数值由小到大依次为全磷、速效钾、全氮、全钾、pH、速效磷、碱解氮和有机质含量,而空间分布格局对尺度的依赖性则相反。全磷含量的分形维数值最小,为1.501,说明具有良好的结构性,这与全磷含量块基比最小且具有强烈的空间自相关性是一致的。
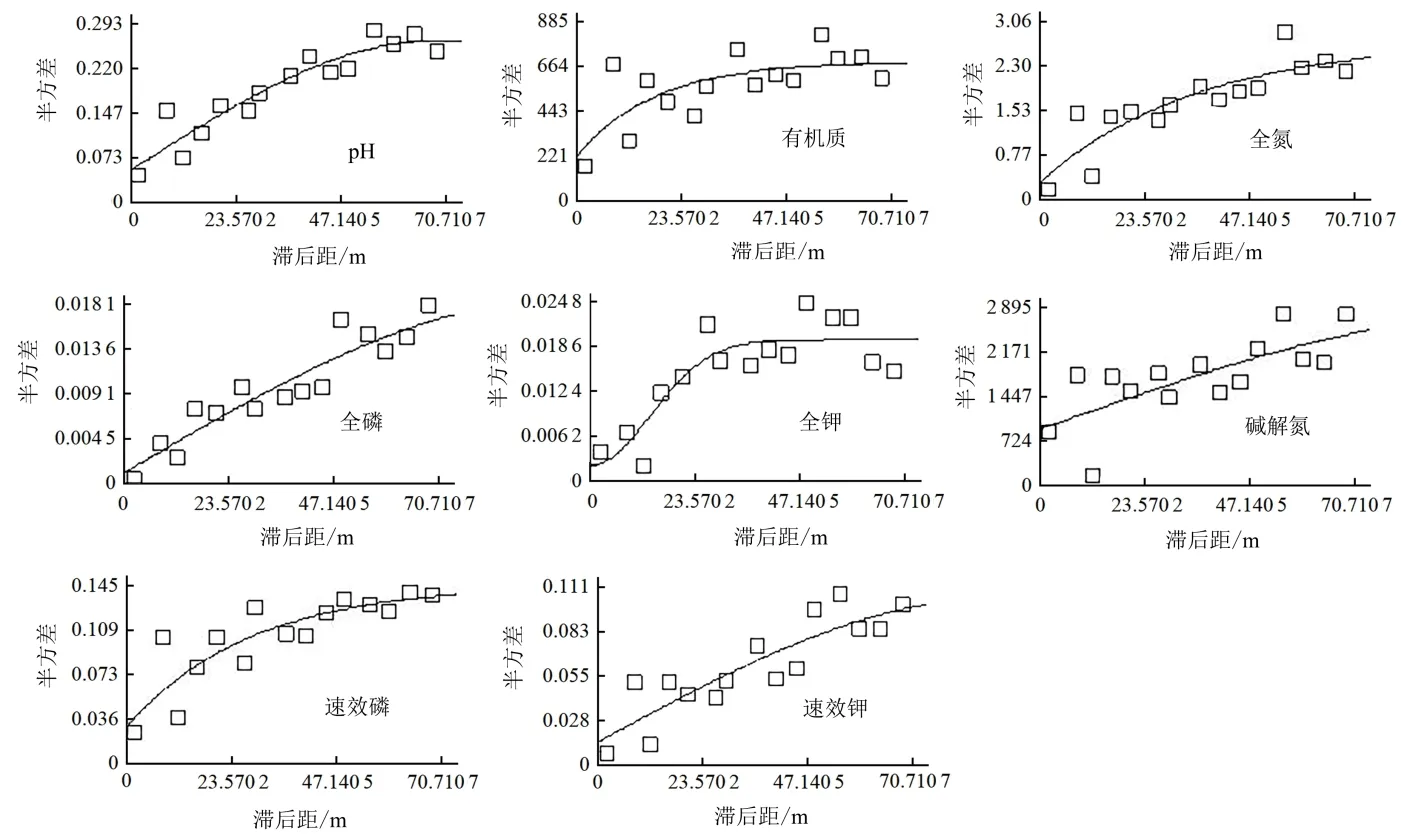
图1 土壤pH 和养分半方差函数图Figure 1 Semivariogram of soil pH and nutrient content
3.3 土壤养分的空间分布格局
由图2 可知,各土壤养分含量主要呈现出斑块状或带状分布格局。土壤pH 相对较高的区域主要集中在研究区北部,呈现带状分布,研究区南部pH 较低,土壤酸化严重,总体呈现西北高东南低的趋势。土壤有机质、全氮和碱解氮含量的总体分布情况相似,表现为研究区西侧高,东南及东北处低,最大值在西及西北角。有机质含量的分布呈现斑状镶嵌分布,空间破碎性高,连续性差。速效磷含量呈明显的“岛”状斑块分布,总体西高东低。全磷含量在研究区南部区域低,西北角高。全钾含量的分布没有明显的规则,高值在研究区北部、东北部顶端、东南底端等均有分布。速效钾含量的分布出现明显的一个块状高值区域与一个低值区域,分别位于研究区西北部和中部偏东处,高低界限比较分明。
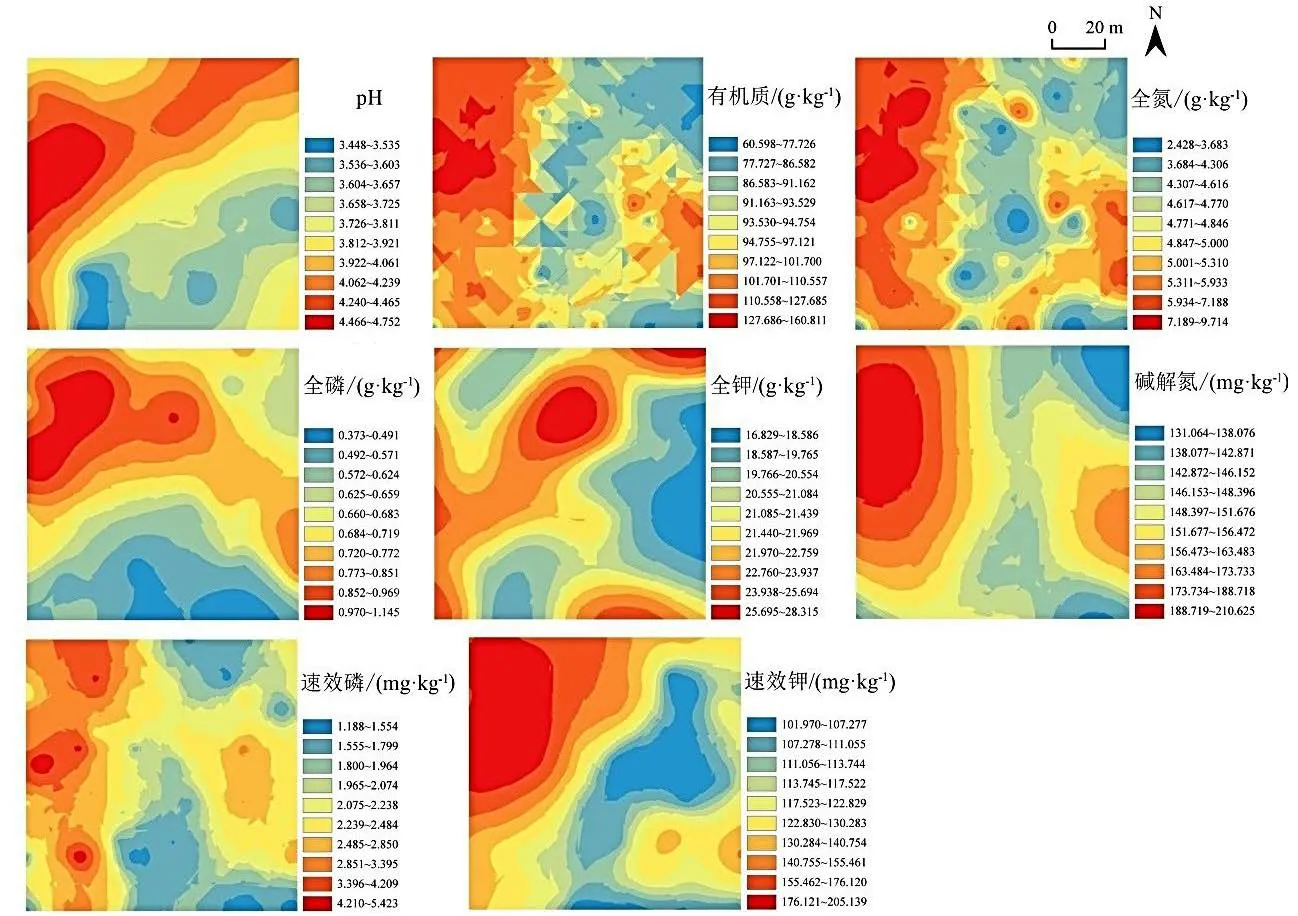
图2 土壤pH 和养分的空间分布格局Figure 2 Spatial distribution pattern of soil pH and nutrient content
3.4 土壤养分与群落组成的相关性
如表3 所示,Shannon-Wiener 多样性指数与土壤pH 和全磷、全钾含量均呈极显著的负相关(P<0.01),与土壤速效钾含量呈显著的负相关(P<0.05),而与其他土壤养分含量无显著相关性。Margalef 丰富度指数与土壤pH 和全磷含量呈极显著的负相关(P<0.01),与土壤全钾和速效钾含量呈显著的负相关(P<0.05)。
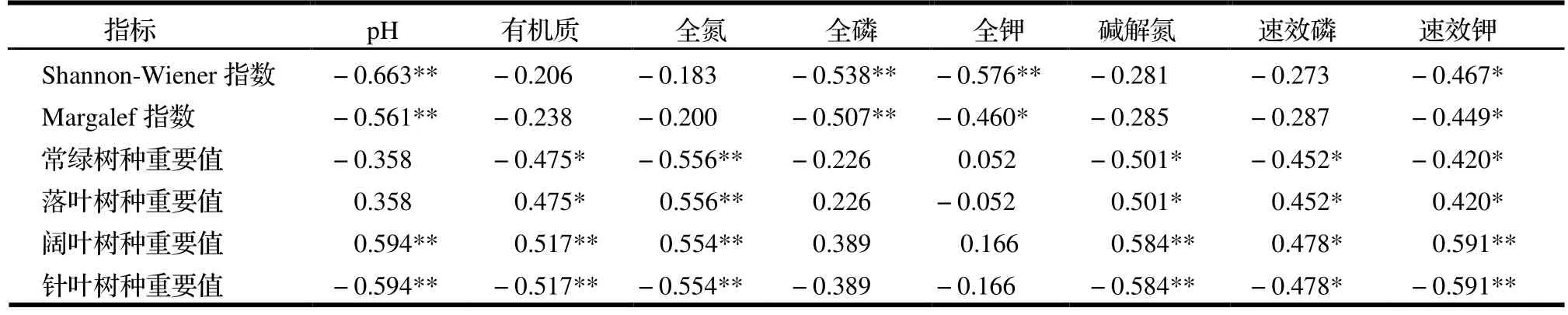
表3 土壤养分与群落组成和多样性之间的相关关系Table 3 Correlation between soil nutrient content and community composition or diversities
根据不同生活型的树种来看,常绿树种的重要值与土壤有机质、碱解氮、速效磷含量有显著的正相关关系(P<0.05),与全氮含量呈极显著正相关(P<0.01),而落叶树种与其相反。针叶树种重要值与土壤速效磷含量存在显著的负相关关系(P<0.05),与土壤pH、有机质、全氮和碱解氮含量存在极显著的负相关关系(P<0.01),而阔叶树种与其相反。这表明不同特性的树种对土壤养分含量有显著的影响。
3.5 土壤养分与树种分布的关系
由图3 冗余分析的结果表明,土壤养分对树种分布具有显著影响(F=2.8,P=0.004),土壤养分对树种分布的解释量为57.9%,第一轴和第二轴的特征值分别为0.485 和0.035,其中土壤pH 是对树种分布影响最大的因素(P=0.002),解释了树种分布变化的38.6%。图中所有线段与排序轴1 的夹角均小于90°,除了全钾位于第4 象限,其余的土壤养分因子都分布在第1 象限,说明所有土壤养分与排序轴1 都呈正相关。从各采样点周围的树种来看,pH 最高的采样点附近紫楠较多,与结果相符;而微毛柃Eurya hebeclados、豹皮樟Litsea coreanavar.sinensis、杉木等较适应在土壤pH 和速效磷、速效钾、全磷含量等较低的环境中。天目木兰Magnolia amoena、星毛稠李Padus stellipila和青钱柳与土壤有机质、全氮、碱解氮含量等线段呈锐角,主要与较高的土 壤有机质、全氮、碱解氮含量相对应。
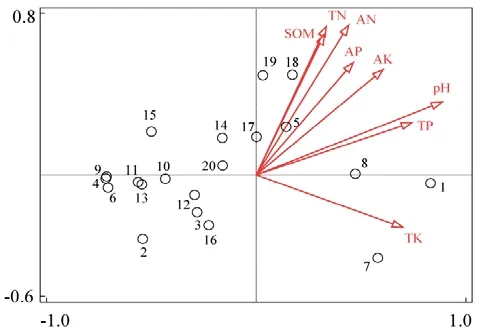
图3 研究区中主要树种与土壤养分的RDA 排序图Figure 3 Redundancy analysis on main tree species and soil nutrient content in sample plots
4 讨论
4.1 土壤养分的空间异质性
土壤养分的空间异质性是由随机性因素和结构性因素共同作用产生的,其结构性因素使同一层面的空间相关性增强,随机性因素使其空间相关性减弱,并朝同质化、均一化方向发展[18]。本研究分析得出,有机质和碱解氮含量具有中等程度的空间自相关性,说明其空间变异是由结构性因素和随机因素共同作用引起的,随机因素又与取样设计、试验误差、生物干扰等原因有关;其余的土壤养分含量存在强烈的空间自相关性,说明随机因素对其空间分布的影响较小,其空间变异主要由结构性因素,例如环境气候、成土母质、地形地貌引起的。研究区土壤养分中碱解氮含量的空间自相关性相对最弱,而全磷含量的块基比最小,分形维数最小,说明其空间自相关性最强,这可能与磷在土壤中移动慢且容易被固定有关,该结果与杜华强等[19]研究天目山常绿阔叶林时发现碱解氮含量为中等程度的空间自相关性和具有最大变程的结果一致。研究区内各土壤养分含量块基比不同,变程也有一定差异,说明土壤各养分的空间异质性对尺度的依赖程度各不相同。
4.2 土壤养分与群落组成的关系
本研究发现,天目山常绿阔叶林土壤pH 和全磷、全钾和速效钾含量均与Shannon-Wiener 多样性指数以及Margalef 丰富度指数呈负相关。Ilka 等[20]和Yang 等[21]的研究也得出了土壤养分含量与物种丰富度呈负相关的结论,随着土壤养分含量的增加,物种的数量却在减少。这也与Rosenzweig[22]的养分富集破坏种群和群落动态的预测一致。土壤养分含量的增加会导致植物种间的竞争加剧,优势种能利用有利的自然条件迅速生长而导致其他共存物种数量降低。但Zheng 等[23]研究表明,天然林中物种多样性与凋落物产量之间存在正相关关系,生物多样性越高的森林中凋落物数量就越多,所以土壤养分也相对更高。我们认为植物物种多样性和土壤养分含量之间不单单只是简单的线性相关,可能在养分贫瘠的土壤中,增加养分含量可以提高物种多样性,而在养分含量丰富的土壤中,少量物种容易形成竞争优势,限制其它物种进入,减少生物多样性。不同研究中植物多样性和土壤pH 的关系也存在一定的差异。Pärtel[24]研究表明,物种丰富度和土壤酸碱度之间的正相关关系更可能出现在进化中心位于高酸碱度土壤的植物区,而物种丰富度和土壤酸碱度之间的负相关关系更可能出现在进化中心位于低酸碱度土壤的植物区。通过RDA 分析可知,土壤pH 是影响研究区植物群落及物种分布最重要的因子,研究区内各土壤养分含量对树种的影响不同,紫楠作为优势树种适应pH 较高的环境。张潇月等[25]的研究也认为土壤pH 是影响植物区系组成的最主要条件。
大量研究只针对了不同类型的森林对土壤性质的影响,而本研究针对天目山常绿阔叶林中不同生活型的树种和土壤养分之间的关系进行进一步的分析。本研究发现,落叶树种和阔叶树种重要值增加对应了较高的土壤养分含量。这主要与不同树种的特性有关,树种的不同会直接影响凋落物的数量以及凋落物的分解速度等。有研究表明,落叶树种凋落物量显著大于常绿树种[26-27]。也有研究说明落叶树种凋落叶比常绿树种凋落叶具有更快的分解速率和养分释放速率[28]。本研究土壤采样在10 月完成,正处于落叶树种凋落期,以及落叶树种凋落物更易被分解的原因,均与本研究中落叶树种重要值与土壤养分含量呈正相关相符。土壤磷和钾含量主要取决于成土母质、土壤类型以及相应肥料的施加,而不易受到植被的影响[1]。本研究结果也表明,植物生活型组成与土壤全磷和全钾含量无显著相关。于法展等[29]研究表明,落叶阔叶林土壤有机质含量最高,林下枯落物厚度以及腐殖质厚度均高于常绿阔叶林。Gustafson[30]认为针叶树种凋落物中所含单宁、树脂类物质较多使得分解后形成酸性环境,这与本研究中pH 与针叶树种重要值呈极显著的负相关相符。在外界环境条件基本相同的情况下,针叶树种的凋落物因为比表面积小、本身含角质类物质较多、分解后呈酸性环境不利于微生物活动等原因,其分解速率要显著小于阔叶树种[31-32]。因此,不同的植物组成因其树种特性会对土壤养分产生一定影响。
5 结论
天目山常绿阔叶林土壤养分具有明显的空间异质性。通过地统计学分析方法得出研究区土壤pH 和全氮、全磷、全钾、速效磷、速效钾含量存在强烈的空间自相关性,其中,有机质和碱解氮含量呈现中等程度的空间自相关性。土壤和植物之间是一个相互作用相互影响的统一整体。研究区土壤pH、全磷、全钾、速效钾含量与生物多样性指数之间呈显著的负相关,落叶树种与阔叶树种对应了相对较高的土壤养分含量,常绿树种与针叶树种对应了相对较低的土壤养分含量,表明天目山常绿阔叶林中植物组成对土壤养分含量有明显的影响。
猜你喜欢
养生阅刊(2021年5期)2021-06-03
上海文化(文化研究)(2021年2期)2021-05-08
湖北农业科学(2019年10期)2019-07-08
安徽农业科学(2018年15期)2018-05-14
文化交流(2018年1期)2018-01-09
现代农业科技(2016年24期)2017-04-19
科技资讯(2017年3期)2017-03-25
中老年健康(2016年12期)2017-01-18
绿色科技(2016年16期)2016-10-11
家庭医药·快乐养生(2015年8期)2015-09-10
