
抗氧化剂抑制血色素蛋白介导的脂质氧化的研究进展
2022-05-14 08:26:28曹珈荣雷雪晴秦志伟
食品工业科技 2022年10期
曹珈荣,雷雪晴,秦志伟,刘 玲
(沈阳农业大学食品学院,辽宁沈阳 110866)
肉类中含有丰富的营养成分,在贮藏和加工中容易出现质量劣变,导致肉色泽、风味和营养价值下降。肉在贮藏加工中劣变的最主要因素之一是氧化,氧化主要存在于肉的脂质和蛋白质中。脂质氧化导致肉产生不良气味,蛋白氧化导致肉色、肉质结构和蛋白功能改变,同时氧化也产生有毒有害的化学物质,严重威胁消费者健康[1]。
肉类中血红素蛋白诱导的脂质氧化是导致肉类食品品质恶化的主要原因。血红素蛋白主要包括血红蛋白(Hemoglobin,Hb)和肌红蛋白(Myoglobin,Mb)两类。研究表明,血红素蛋白对脂质氧化具有介导作用,但是两种血红素蛋白结构差别对氧化有影响。有人认为Hb比Mb更易使脂质氧化,因为在氧化条件下从血红素中释放的高铁血红蛋白(Methemoglobin,metHb)比保留原卟啉形式的Mb更容易促进脂肪的氧化[2]。有研究指出Hb促氧化活性的丧失是阻止鱼类脂质氧化的决定性因素[3]。
在控制肉类和肉类制品中的脂质氧化各种方法中,抗氧化剂的应用被认为是一种实用的选择。常用的肉制品抗氧化剂包括丁基羟基茴香醚(BHA)、二丁基羟基甲苯(BHT)和特丁基对苯二酚(TBHQ)等。它们主要用于清除自由基并终止自由基链的增殖[4]。有些抗氧化剂可以有效抑制血色素蛋白的介导作用[5]。但是,相较于对合成抗氧化剂安全性的担忧,大众普遍认为天然抗氧化剂本身更安全、更健康,这促使肉类工业寻找植物源抗氧化剂取代合成抗氧化剂。有研究表明天然抗氧化剂可以控制血色素蛋白介导的肉类脂质氧化[6]。
本文阐述了肉制品中血红素蛋白氧化和脂质氧化反应机理,并探讨抗氧化剂对抑制脂质和血红素蛋白氧化的作用,为肉制品在贮藏和加工中的质量控制提供基础,并为开发合适的抗氧化剂提供思路。
1 脂质氧化与血红素蛋白氧化
蛋白质和脂质是肉类的主要营养成分,它们在氧气、光照、酶等促氧因子的作用下发生氧化。氧化会导致肉制品的品质下降,其中脂质氧化不仅会导致肉质酸败,而且还会对肉的感官有很大影响[7]。特别是在富含n-3多不饱和脂肪酸(PUFA)的鱼类中,氧化导致质量损失的风险很高[8]。除了脂质氧化外,蛋白质的氧化也是导致肉品质下降的重要因素。肌浆蛋白中的血红素蛋白是肉制品氧化的主要催化剂,由于血红素蛋白自氧化产生活性氧和自由基,进而激活肉制品的氧化过程,同时血红素蛋白也能释放血红素和Fe2+促进脂质的氧化。脂质氧化和蛋白质氧化过程可以独立发生,但在肉制品中它们之间通常存在相互作用,蛋白质和脂质之间的相互作用明显增强了初始氧化进程[9]。
1.1 脂质氧化
脂质氧化是典型的自由基增殖过程,常见的自动氧化通过不饱和脂肪酸经增殖反应生成烷基自由基、羟基自由基和过氧化自由基等,进一步形成氢过氧化物(图1)。这些初级产物经过分解和聚合生成脂质氧化的次级产物,如脂质过氧化反应中产生的活性壬烯醛、丙烯醛、丙二醛等都具有很强的氧化能力,其中不饱和醛促氧化能力比饱和醛更强[10]。醛类氧化产物具有促进血红素蛋白氧化的能力,它们可通过增加氧合肌红蛋白的氧化、降低高铁肌红蛋白的酶促还原能力、提高高铁肌红蛋白的促氧化活性而改变其稳定性[11]。

图1 脂质自氧化[12]Fig.1 Lipid autoxidation[12]
1.2 血红素蛋白氧化
血红素蛋白的氧化主要由其中铁离子的氧化变价引发,这个过程逐步引起血红素蛋白变性[13]。变性后的蛋白对血红素的保护作用下降,导致血红素基团暴露和氧化敏感性增强,进一步引发血红素释放,同时铁离子部分游离,导致脂质氧化。有人在牛肉和鸡肉中直接添加metHb,并没有发现它明显地介导脂质氧化[14],这说明肌体中metHb需要在氧化条件下转变成ferryHb或者释放高价铁离子来氧化脂质,条件不具备时metHb对脂质氧化的作用有限。在动物组织中,90%的铁以血红蛋白、肌红蛋白和少量铁蛋白、转铁蛋白和一些酶的活性部位的形式存在。其中,肉中血红素蛋白含量最高的是肌红蛋白和血红蛋白[1]。
Hb分为脱氧血红蛋白(deoxyHb)、氧合血红蛋白(oxyHb)、高铁血红蛋白(metHb)和超铁血红蛋白(ferryHb),氧化过程见图2。deoxyHb(II)的卟啉环上的二价铁未携带氧气,呈蓝紫色,与氧气结合后生成oxyHb(II),氧气分子与二价铁以一种可逆的方式结合在一起。oxyHb(II)在常温下可以发生自动氧化反应,产生超氧阴离子自由基和褐色metHb(III)。oxyHb(II)的自动氧化反应是肉色红度值(a*)损失的一个重要原因[15]。metHb(III)能够被 H2O2激活而形成不稳定的ferryHb自由基(·HbFe4+),进一步还原成稳定的ferryHb(HbFe4+),它具有很强的促进肉中脂质氧化的能力[16]。高价铁Fe4+是强氧化剂,在动力学上等价于羟基自由基(·OH)的活性,能够快速从氢过氧化物上抽氢来促进脂质氧化。O2−等也可以进一步与肉中的铁发生反应生成·OH,而·OH可以扩散进入脂质的疏水区域,具有促进脂质氧化的能力[17]。由deoxyHb向metHb的转变过程中,生成的各类自由基等对促进脂质氧化具有明显作用。
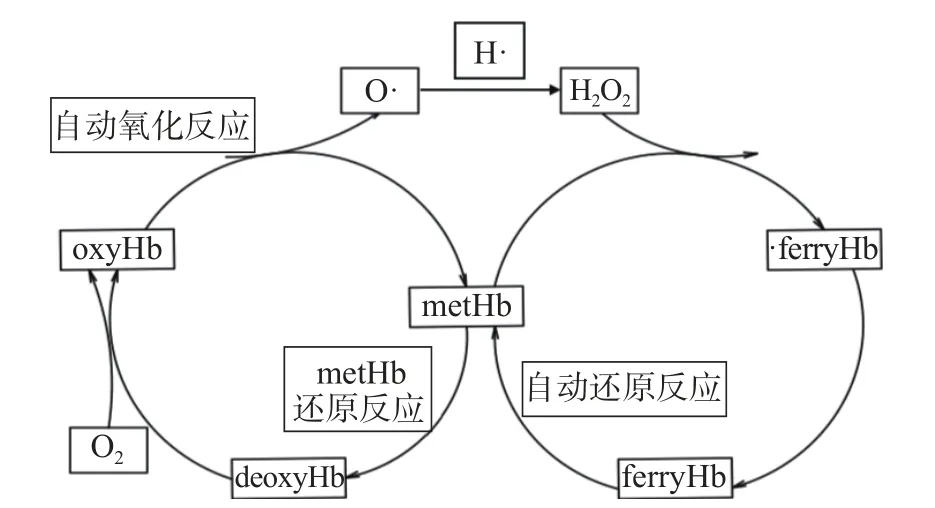
图2 Hb的氧化过程Fig.2 Oxidation process of the hemoglobin
1.3 血红素蛋白与脂质的相互氧化
蛋白与脂质的氧化是相互作用的。有人认为蛋白的氧化由脂质引发,这是一类脂质自由基促发蛋白自由基的反应[1]。但多数人认为,氧合血红素蛋白的氧化是导致脂质氧化和非血红素蛋白氧化的主因[18],也就是说 deoxyHb氧化形成 ferryHb后,血红素从ferryHb解离出来导致自由基的形成,自由基降解原卟啉并导致脂质氧化[19]。Hb诱导脂质氧化有两种方式:一种是蛋白氧化产物诱导脂质氧化,如超氧自由基、烷基自由基、高铁血红素等诱导脂质的氧化;另一种方式是通过Hb释放的铁离子催化脂质氧化[1]。铁离子在氧化的过程中生成具有高反应活性的自由基,启动了脂质氧化反应。
如图3所示,脂质氧化过程启动后,反过来血红素蛋白被进一步氧化,尤其是脂质中次级氧化产物不饱和醛的氧化作用。Chen等[20]发现4-羟基-2-壬醛和己烯醛能加快氧合肌红蛋白的氧化速率,抑制电子传递链介导的高铁肌红蛋白还原。Gunwald等[11]研究发现4-羟基-2-壬烯醛能够改变肌红蛋白的蛋白质结构和氧化还原性质,它甚至能在5个位点上与抹香鲸的肌红蛋白共价结合,使经过诱导的肌红蛋白有效地促进肌肉中的脂质氧化。Gęgotek等[21]研究发现,在较高浓度下的4-壬烯醛(4-HNE)可与组氨酸产生加合物,在DNA水平上修饰蛋白质的表达。有人总结出将脂质的初级和次级代谢产物与Hb反应,醛类比ROOH更易引发metHb形成[22]。由此可见,脂质氧化血色素蛋白的过程主要是通过脂质氧化产生的活性醛等中间产物实现对Hb的氧化。这些醛类物质不仅改变了血红素蛋白的氧化还原稳定性,而且促进含氧血红素蛋白的氧化,降低高铁血红素蛋白的还原并增强了其促氧化活性[1]。

图3 脂质氧化和肌红蛋白氧化的相互促进关系[23]Fig.3 Interacting oxidation reactions between myoglobin and lipid oxidation[23]
2 抗氧化作用机理的研究
为减少肉制品氧化,目前通常采用控制底物、加入抗氧化剂,糖基化反应改性及酶法处理等方法,其中最为有效的是添加抗氧化剂。对于抑制血红素蛋白介导脂质氧化,抗氧化剂作用主要表现为改变Hb结构、控制铁离子的释放、调控血红素蛋白的氧化状态、竞争氧化作用底物四个方面。
2.1 血红素蛋白结构对脂质氧化的影响和抗氧化作用
血色素蛋白氧化易于发生在各种肌肉中,尤其是一些鱼肉中[24]。研究发现,在pH5.7和pH6.3的条件下,鳟鱼和鲈鱼Hb的自动氧化速率可以达牛Hb的30~80倍,其血红素释放速率也可到达牛Hb的50倍,这些差异与鱼和哺乳动物Hb立体化学结构的不同有很大关系[25−26]。如图4所示,牛Hb的两个亚基Val E11的甲基基团与束缚配体之间距离为3.2~3.4 A,鲈鱼的Ile(E11)正丁基侧链上的Cd原子与束缚配体之间距离2.4 A左右,比牛Hb短很多,这种增加的空间位阻导致E11和束缚配体的结构错乱。而且鲈鱼Hb在E14位点常含有甘氨酸,而牛Hb含有较大的残基(如丙氨酸和丝氨酸)。E14中的甘氨酸为溶剂进入血红素缝隙提供了一个通道,从而增加了血红素损失率,促进自动氧化,这部分地解释了鱼类Hb比牛Hb更有效地促进脂质氧化的原因[27]。血红素裂缝中的氨基酸组成、排列顺序以及血红素口袋的大小和方向都影响质子结合和质子转移,而血红素结构和配体会影响质子的转移过程和高铁复合物的稳定性。另外,即使同样是哺乳动物,其血红素蛋白的结构差异也影响氧化水平。Suman等[28]比较牛和猪的Mb,发现牛OxyMb近端组氨酸(His 93)优先与HNE结合,而猪OxyMb中没有出现这种结合,因此牛肉中OxyMb的氧化比猪肉中OxyMb氧化更常见。
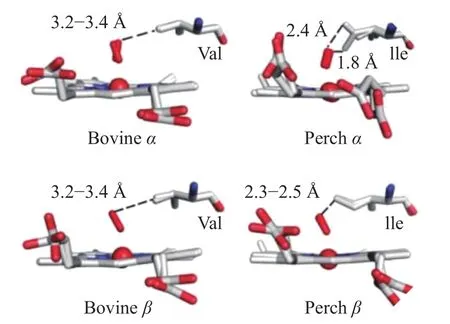
图4 不同pH下牛和鲈鱼Hb结构比较[25]Fig.4 Comparison of the hemoglobin structures between bovine and perch at different pH[25]
Khadega等[29]在研究重金属对两种蛋白质细胞色素C(一种Hb)和Mb构象的影响中,分别添加动物来源的甘氨酸和抗坏血酸及植物来源的没食子酸和芝麻酚,用吸收光谱的变化表示这些血红素蛋白的结构改变,发现这些抗氧化成分对重金属暴露具有保护作用,抗氧化剂抵消了重金属的有害作用。添加重金属和抗氧化剂后,过氧化物酶活性也发生改变。
槲皮素也是一种有效的抗Hb氧化的成分。MetHb能与一分子槲皮素通过共价键结合形成加合物,加合后Hb构象发生变化,血色素流失速率降低,从而导致Hb促脂质氧化能力下降。另外,槲皮素能够与细胞膜发生交互作用,改变细胞膜流动性和位阻作用,从而降低脂肪酸受脂质自由基攻击的概率,抑制脂质氧化[30]。
2.2 血色素蛋白中铁离子对脂质氧化的影响和相应抗氧化作用
有人认为铁元素催化脂质过氧化物形成,因此铁的释放和催化是脂质氧化的关键[2]。一方面,血色素蛋白降解产生游离铁,游离铁在肉中的催化氧化作用导致脂质氧化[31];另一方面铁离子在血色素蛋白中发生变价作用,从而导致脂质氧化[1]。例如在鲈鱼的氧化中,deoxyHb和metHb在氧合态变化过程、血红素铁的变价过程、高铁血红素蛋白中血红素的丧失及血红素铁的游离等过程中都能形成自由基,从而促发脂质氧化[32]。Reeder等[33]研究发现质子化铁离子从蛋白质或卟啉中提取电子,使铁离子快速再生。通常这种形成离子的过程发生在酸性条件下,此时还原酶被钝化,酸催化铁离子从二价氧化为三价,引发氧化;而碱性条件中的离子稳定作用使Mb对脂质和脂质过氧化物的反应大大降低。铁离子的存在是否直接引发肉的氧化还存在争议,有人在非加热条件下对比血红素蛋白铁离子和无机铁离子对脂质氧化的作用,发现无机铁对于脂质氧化作用不大[34],但是多数学者认为有机铁和无机铁的变价对于脂质氧化都相当重要。
抗氧化剂咖啡酸被用来防止卟啉降解和脂质氧化[19]。将小分子咖啡酸加入水洗肉中,它能够通过电子转移至Hb的配体O2上促进metHb的形成,但是同时它也束缚metHb中血红素的分离,因此抑制Hb介导的脂质氧化[35]。Fe4+Hb是Hb被过氧化氢(H2O2)氧化生成的产物之一,它是一种强氧化剂,可产生游离的球蛋白自由基。研究发现,在Hb中加入酪氨酸后,Fe2+Hb和metHb未被氧化成Fe4+Hb,说明酪氨酸在该反应中对Hb具有保护作用,可有效抑制Hb氧化[36]。
2.3 血红素蛋白的氧化状态对脂质氧化的影响和相关抗氧化作用
DeoxyHb和metHb都有助于促氧化。对鱼类的研究发现,还原态的deoxyHb比氧化态的metHb更易于氧化脂质,因为从还原态向氧化态氧化转变过程中形成了超氧阴离子从而加速脂质氧化。另外,血红素蛋白结构上具有一个血色素裂缝,当血红素蛋白由含氧状态转变成脱氧状态时,卟啉环上铁元素从这个裂缝中被排除,使铁更容易接触脂质过氧化物,更容易引发氧化[37]。不同来源鱼肉的Hb介导脂质氧化比较,因血红素裂缝的大小不同而氧化能力有差异,得到了血红素蛋白介导的脂质氧化与血红素铁的变化相关的类似结论[38]。Vuletich等[39]从另一个角度阐述Mb的氧化对脂质的影响,他认为Mb具有一种氧化变形态Mb-H,其氧化纯磷脂酰胆碱的速率比天然Mb氧化速率高至少7倍,说明Mb-H在Mb对脂质过氧化过程发挥重要作用。采用糖基化反应对血红素蛋白进行改性也有利于血红素蛋白氧化状态的改变,它能使高铁蛋白转化为氧合蛋白,从而降低血红素蛋白对脂质的氧化促进作用[40−42]。
在抗氧化方面,有研究表明槲皮素能使MetHb还原为OxyHb,从而降低Hb促氧能力,抑制脂质氧化[30]。一些鱼类的研究发现红细胞具有metHb还原酶活性,当metHb由Hb自氧化作用或与亚硝酸盐反应而产生,还原酶活性能抵消Hb自氧化作用和遇到亚硝酸盐的可能性,从而抵抗Hb向metHb的持续氧化,其中脱氧作用显著提高红细胞对metHb的还原率[43]。
2.4 肉中氧化底物对蛋白和脂质氧化的影响和相应抗氧化作用
在肉制品中主要的脂质包括磷脂、胆固醇和甘油三酯,它们水解能产生游离脂肪酸(Free fatty acid,FFA)。FFA特别是不饱和脂肪酸是脂质氧化的主要成分。肉的氧化也取决于其中血色素蛋白含量和水分含量。Sannaveerappa等[44]分析水洗鳕鱼肉糜模型发现Hb有时对其介导的脂质氧化不起作用,原因是水分已被洗至最低界限,因此降低Hb含量和减少水分含量可以有效降低氧化程度。Castell等[45]发现在含有Hb鳕鱼肉糜中添加FFA能有效抑制脂质氧化,认为这可能与FFA和血红素蛋白交互作用导致血红素蛋白失去促氧活性有关[46];也有人发现FFA能显著抑制火鸡肉糜的脂质氧化[30]。Dong等[31]发现没食子酸对Hb具有抗氧化还是促氧化作用,主要取决于其浓度和Hb的氧化还原状态。Wu等[47]发现添加的肌红蛋白(Mb)促进了鳕鱼洗过的肌肉中的脂质氧化,但没有促进猪洗过的肌肉中的脂质氧化。
Tatiyaborworntham等[24]以铁酰血红蛋白(feryl Hb)和metHb为研究对象,探讨磷脂酶A2(PLA2)对Hb介导的水洗鳕鱼肌肉脂质氧化的抑制作用。发现feryl Hb可能在Hb介导的脂质氧化中发挥作用,而PLA2抑制了metHb和feryl Hb的形成,且PLA2能够与Hb反应,清除了脂质底物中预先形成的脂质过氧化氢(LOOHs)。他们还发现随着更多的血红素与水洗肌肉的不溶性基质结合,脂质氧化进入指数期的滞后期也相应缩短,脂质氧化加速。而PLA2虽能够抑制脂质氧化,却不阻止血红素和水洗肌肉的不溶性基质之间的相互作用[3]。吴海舟[30]发现不仅PLA2在漂洗肌肉体系中能有效抑制Hb诱导的脂质氧化,而且PLA2水解产生的FFA在抑制脂质氧化过程中也发挥重要作用。他们的抑制脂质氧化机理可能涉及FFA与血红素蛋白结合形成了低促氧活性的高铁血色原,另外磷脂膜水解导致细胞膜结构发生改变可能是PLA2能抑制脂质氧化的一个原因。
3 抗氧化剂在肉制品中的应用
肉制品在贮藏加工过程中,脂质和蛋白质作为主要成分易于氧化,并且两者相互作用。其中血红素蛋白自氧化和脂质自由基形成是肉制品氧化的开始,随贮藏时间延长,蛋白和脂质的氧化相互作用。肉的脂质氧化不仅会影响感官特性和营养价值,还会导致有毒物质的形成,因此控制氧化过程对肉制品至关重要。尽管脂质氧化已被研究了数十年,但不同内外在条件引起的反应途径有差异,导致脂质氧化机制尚未完全被解析。食品研究人员一直在寻找抑制肉制品脂质氧化的有效方法,在食物中添加抗氧化剂是目前最常见、最有效的方法[48]。
抗氧化剂通过提供电子中和自由基、螯合氧气、改变系统的氧化还原电位来减少产品的的氧化劣变[49]。合成抗氧化剂对延长产品的保质期很有效,但因其具有潜在毒性,在食品中的使用存在很大局限性。目前在食品加工中用天然抗氧化剂部分或完全替代合成抗氧化剂已经成为必然。
植物富含生物活性化合物(多酚类和黄酮类等物质),可用作抗氧化剂的潜在来源,替代合成抗氧化剂用于保存肉制品并改善产品营养质量。表1是在目前肉制品中常使用的天然抗氧化剂,包括提取物的植物种类,提取物的浓度和在肉制品中的应用效果。这些天然成分以提取物和精油的形式从不同植物获得,如水果、蔬菜、草药和香料等。这些提取物与合成抗氧化物质相比,具有相似或更好的抗氧化性能,它们在新鲜肉制品中可以保持氧化稳定性,护色,减少脂质和蛋白质的氧化以及脂质衍生的挥发性化合物的生成,并不会影响感官属性,其中白藜芦醇具有显著的肉制品抗氧化作用[50]。此外,植物成分还有助于提高原料的功能特性,提供更加多样化的健康食品。

表1 肉制品中的天然抗氧化剂Table 1 Natural antioxidants in meat products
天然抗氧化剂在抑制血色素蛋白介导的脂质氧化也具有十分显著的效果。Lee等[51]研究表明,植物中花青素、黄酮醇、原花青素等酚酸在Hb介导的水洗鱼肌肉脂质氧化中具有抗氧化作用,其中槲皮素可有效抑制Hb介导的脂质氧化。他在鳟鱼和牛的Hb研究中发现Hb与甲基的分离会形成自由基,自由基会降解原卟啉并导致脂质氧化,而加入的咖啡酸可以有效阻止卟啉的降解和脂质氧化[13]。Harun等[52]研究发现MO叶提取物可以通过抑制MetHb起到抗氧化作用。Maqsood等[53]对罗非鱼Hb介导的脂质氧化和抗氧化研究中发现添加单宁酸后过氧化值降低,储存过程中TBARS生成量也很低。在鱼肌肉、鱼肉末、鱼油-水乳状液和鱼切片中,单宁酸对Hb介导的脂质氧化均有抑制作用。Das等[54]研究表明姜黄素、维生素E、维生素C等天然抗氧化剂能够抑制亚硝酸盐诱导的met-Hb形成;咖啡因及其代谢物与Hb的络合,可以防止二价铁的氧化。Wu等[5−6]研究发现迷迭香、抗坏血酸、生育酚和柠檬酸的混合物防止自氧化和鲱鱼Hb的血红素丢失,明显抑制鲱鱼副产品中Hb介导的脂质氧化,将保质期从1 d增加到12 d以上。Liu等[55]评估了三种天然抗氧化剂的相互作用,及其对生牛肉肉饼的颜色、脂质稳定性、metMb占比和metMb还原活性的影响,发现茶儿茶素,肌肽和α-生育酚混合物中儿茶素含量的增加可提高脂质稳定性,降低metHb百分比;肌肽对提高颜色稳定性具有显著作用;α-生育酚和茶儿茶素增强了脂质的稳定性;三者相互作用可改善生牛肉肉饼的脂质稳定性和颜色稳定性。从这些抗氧化剂的作用效果上看,抑制血红素蛋白的氧化是控制脂质的氧化的有效手段。
有些植物成分的抗氧化性往往具有剂量依赖性,常表现为对肉制品的氧化和抗氧化的双重作用。Chen等[56]研究VC对牛Hb氧化还原反应的影响,发现VC在反应初期表现出抗氧化作用,后期通过产生过氧化氢来促进MetHb含量的增加,证实VC既可以抗氧化又具有促氧化能力。日常饮食物质NO2−在理论上可作为还原剂去除具有细胞毒性的ferryl Hb,有效地抑制ferryl Hb引发的蛋白羰基,然而NO2−在低浓度时却能促进Hb-H2O2导致的蛋白质氧化[57]。Dong等[58]研究发现茶多酚通过插入其疏水口袋与Hb的色氨酸和酪氨酸残基相互作用。这种相互作用显示出对Hb的浓度依赖性影响,这可能导致完全相反的结果。高浓度茶多酚破坏了Hb的结构,使血红素铁的暴露促进Hb氧化和变色。然而,较低浓度的茶多酚不会破坏Hb结构,可以抑制高铁血红蛋白的形成。Naparło等[59]进一步研究EGCG和没食子酸,发现它们能够氧化Hb,因为它们都含有三羟基苯甲酸残基,但在一定浓度范围内它们具有抑制氧化的作用。
综上所述,大量研究证明了天然抗氧化剂可以有效抑制血红素蛋白介导的脂质氧化。不仅可以延长货架期,还可以改善感官属性,提高肉品品质。此外,植物源性食品添加剂的抗氧化特性和作用机制以及在肉类工业中应用前景的研究也取得了显著进展。尽管如此,但天然抗氧化剂的效用、可用性、市场性和潜在的健康益处尚未得到充分证明。
4 展望
肉制品中脂质和蛋白质氧化及其交互氧化导致肉品色泽、风味、质构等一系列改变。血红素蛋白介导脂质氧化,脂质氧化反过来对血红素蛋白也有促氧化作用。抗氧化剂可以有效地抑制血红素蛋白氧化而延缓脂质的氧化,从而延长肉制品的货架期。水果、蔬菜及一些农副产品等植物中的活性成分是抑制血红素蛋白氧化的良好选择,但尚未在肉制品中广泛使用。这些活性成分在加入肉制品之前还需要毒性评估,以便确定适用性。因此,对此类抗氧化剂进行深入研究,探究它们抑制肉制品氧化的深层机理,将对未来肉制品抗氧化剂的广泛使用起到至关重要的作用。
猜你喜欢
现代临床医学(2021年2期)2021-03-29 05:32:00
解放军医学院学报(2020年12期)2020-03-29 05:11:02
现代养生·下半月(2019年12期)2019-01-22 04:38:31
中学课程辅导·教师教育(中)(2017年9期)2017-10-24 13:03:32
医学研究杂志(2015年11期)2015-06-10 06:44:03
肝胆胰外科杂志(2015年2期)2015-02-27 11:11:49
食品科学(2013年13期)2013-03-11 18:24:06
吉林医学(2013年13期)2013-01-31 03:19:04
当代畜禽养殖业(2012年11期)2012-02-18 01:16:40
化学分析计量(2010年4期)2010-04-10 13:00:27
